Open Journal of Ophthalmology
Vol.04 No.04(2014), Article ID:51509,14 pages
10.4236/ojoph.2014.44021
The Emerging Role of Statins in Glaucoma Pathological Mechanisms and Therapeutics
O. Pokrovskaya1, D. Wallace1,2, C. O’Brien1,2
1School of Medicine and Medical Science, University College Dublin, Dublin, Ireland
2Department of Ophthalmology, Mater Misericordiae University Hospital, Dublin, Ireland
Email: olya.pokrovskaya@gmail.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 15 September 2014; revised 16 October 2014; accepted 13 November 2014
ABSTRACT
Statins inhibit the enzyme 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase, and hence have a profound effect in lowering serum cholesterol. Their predominant clinical use to date is in primary and secondary prevention of cardiovascular disease. However recently interest has developed regarding the so-called “pleiotropic” effects of statins—these drugs have significant anti-fibrotic, anti-inflammatory, and immunomodulatory properties. Such effects of statins have already been shown to be beneficial in modulating the pathological mechanisms involved in pulmonary fibrosis, renal disease, non-ischaemic cardiac failure, and tissue scarring. Many of these actions are mediated by inhibition of the Rho kinase pathway. Epidemiological studies suggest that patients who take statins have a lower risk of developing glaucoma, and lower rates of glaucoma progression. Here, we review what is known about the pleiotropic effect of statins to date, and examine how these effects may modulate the molecular mechanisms involved in glaucoma pathogenesis.
Keywords:
Statin, Glaucoma, Rho Kinase, Optic Nerve, Trabecular Meshwork

1. Introduction
Recent population-based studies suggest that the use of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase inhibitors (commonly known as statins) may reduce the risk of glaucoma development and progression [1] [2] . Statins are already established as a first-line treatment in cardiovascular disease due to their cholesterol lowering effect. However, statins also possess important pleiotropic immunomodulatory and anti-inflam- matory capabilities, which have resulted in an expansion of their indications for usage [3] . They appear to have a neuroprotective effect in several diseases of the central nervous system including Alzheimer’s disease, and ischaemic stroke [3] . They also have an antifibrotic action—as seen in models of pulmonary and renal fibrosis [4] - [6] . The mechanism of action of statins in these disease processes is still the subject of much debate. Numerous molecular pathways seem to be involved, including the Rho-kinase pathway and modulation of the actin cytoskeleton [7] - [9] . We hypothesise that statins may also be protective in glaucoma. Glaucoma is a progressive optic neuropathy, with a known fibrotic and neurodegenerative component [10] . The purpose of this article is to review the emerging role of statins in primary open angle glaucoma (POAG). In order to understand the potential mechanisms of action of statins in glaucoma, it is beneficial to first review the aetiology of the disease process and current treatment options.
2. POAG
Glaucoma is the second leading cause of irreversible, but potentially preventable vision loss worldwide, affecting an estimated 60.5 million people [11] - [13] . It refers to a spectrum of clinical conditions distinguished by a progressive optic neuropathy and associated visual field loss, with raised intraocular pressure (IOP) being one of the primary risk factors [14] [15] . Two principal theories have been described for the pathophysiological mechanism for primary open angle glaucoma (POAG), but these are not mutually exclusive. The vascular theory proposes that optic neuropathy results from a compromise to the microvasculature resulting in ischaemic damage to the optic nerve head. This may be as a consequence of the raised IOP or other factors which influence ocular blood flow [16] . The mechanical theory suggests that raised IOP alters the architecture of the lamina cribrosa (LC), compromising the retinal ganglion cell (RGC) axons as they pass through this structure. The LC is a specialised series of plates of connective tissue which acts as a support for axons as they traverse the corneoscleral shell.
The normal intraocular pressure (range 11 - 21 mmHg) is dependent upon a delicate balance of aqueous humour production, inflow and outflow [17] . Aqueous humour is produced by the ciliary body, and provides nourishment to the avascular cornea and lens (Figure 1). The main drainage pathway for the aqueous from the anterior chamber is the pressure-dependent conventional outflow pathway—through the trabecular meshwork (TM).
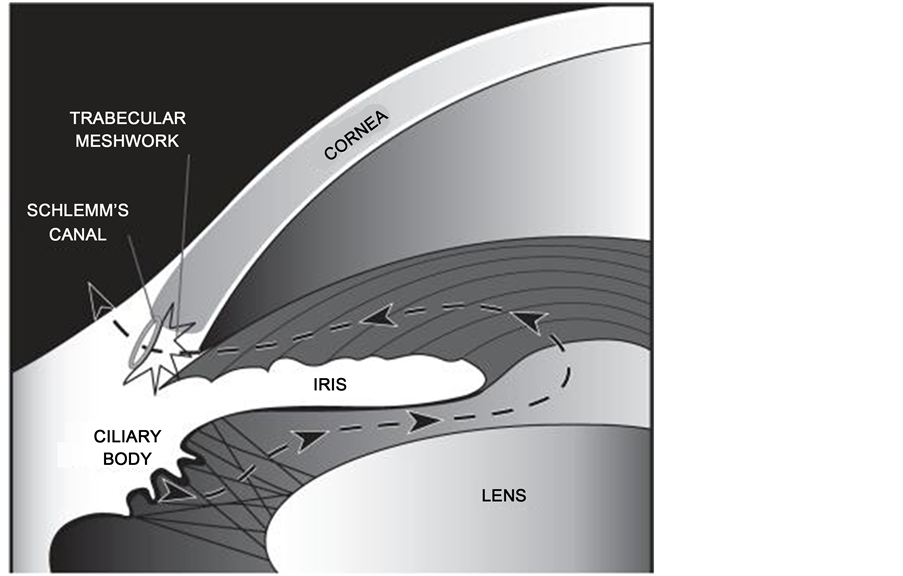
Figure 1. The aqueous outflow pathway (Goel, M., et al., Aqueous humor dynamics: a review. Open Ophthalmol J, 2010, 4, pp. 52-59).
This normally accounts for up to 90% of aqueous outflow. A framework of lattice-like collagen beams forms the TM, with extracellular matrix (ECM) residing in the paracellular spaces between the beams [18] . Aqueous humour drains through the TM, then the juxtacanalicular connective tissue (JCT) region, and then into Schlemm’s canal (SC), from where it is returned to the systemic circulation via the episcleral veins. In order to enter SC, aqueous must diffuse through the paracellular spaces between the endothelial cells of SC, or through giant vacuoles which form as outpouchings into SC. The unconventional, or uveoscleral outflow pathway normally accounts for the remaining ~10% of aqueous outflow. However, with increasing age, up to 50% of aqueous outflow can occur by the uveoscleral pathway [18] .
Extracellular matrix remodelling is at the heart of glaucomatous pathophysiology, and has been shown to be a feature of glaucomatous damage both at the TM where it impairs aqueous drainage, and at the optic nerve head— particularly the LC [19] [20] . Importantly in the LC, remodelling affects its biomechanical properties—leading to increased tissue stiffness, fibrosis, and posterior bowing of this structure [14] [16] [21] [22] . The resultant mechanical compression of the RGC in this area induces axoplasmic stasis contributing to cell death [10] [23] . Two important types of cell that populate the LC include astrocytes (glial fibrillary acidic protein (GFAP) positive) and LC cells (GFAP negative) [24] . Results of previous research in our laboratory support the profibrotic nature of LC cells when exposed to glaucomatous stimuli, suggesting they play an integral role in glaucomatous ECM remodelling [25] - [27] .
Present day glaucoma therapies aim only to reduce the IOP—the only modifiable risk factor for glaucoma progression. Most agents aim to reduce aqueous production (beta-blockers, carbonic anhydrase inhibitors, alpha- agonists), and some increase outflow via the uveoscleral pathway (prostaglandin analogues). However glaucoma may progress despite apparently normal IOP, suggesting the need for novel treatments that may modulate the inflammatory and fibrotic aspects of glaucoma pathophysiology. Our own research group has investigated the role of anti-connective tissue growth factor (CTGF) as a potential anti-fibrotic agent in glaucoma. We have shown that anti-CTGF is effective in blocking extracellular matrix production in human TM and LC cells [28] . Very promising novel agents are inhibitors of the Rho pathway, which is integral to the regulation of aqueous outflow and matrix composition [29] [30] . Various Rho kinase inhibitors are already in phase 2 and 3 clinical trials, with conjunctival hyperaemia being one of the main adverse effects [31] . Clinical trials of combination therapies of Rho kinase inhibitors with prostaglandin analogues are also underway.
3. The HMG CoA Pathway
Statins are inhibitors of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase—the crucial rate limiting step in hepatic cholesterol biosynthesis. Independently of the cholesterol lowering effect, the inhibition of HMG-CoA leads to reduced synthesis of isoprenoid intermediaries—farnesyl pyrophosphate (FPP) and geranylgeranyl pyrophosphate (GGPP) (Figure 2) [32] . These precursors of cholesterol biosynthesis are essential for post-translational modification and prenylation of certain small GTPase proteins, include the Ras and Rho superfamilies [33] . Prenylation of these proteins facilitates their intracellular trafficking and covalent attachment to the lipid membrane, which is often essential for biological function. These small GTPases cycle between an inactive GDP bound form found in the cytosol in association with guanine dissociation inhibitor (Rho GDI), and an active GTP bound form usually associated with the cell membrane (Figure 3). When Rho proteins are released from GDIs, they can insert into the cell membrane where they are activated by guanine nucleotide exchange factors (GEFs), and this initiates interaction with membrane effector proteins such as Rho kinase (ROCK). The Rho family of proteins is implicated in many key intracellular events and signalling pathways, including regulation of the actin cytoskeleton, cell adhesion, cell to cell interaction, and cell-cycle progression. This topic is comprehensively reviewed by Burridge and colleagues [34] . It is thought that inhibition of the Rho pathway by statins is one of the major mechanisms via which statins affect cell physiology.
4. Statins
4.1. Chemical Structure, Pharmacokinetic, and Pharmacodynamics Properties
While all statins share a common mechanism of action, they differ in the chemical structures, pharmacokinetic profiles, and lipid-modifying efficacy. Lovastatin, pravastatin, and simvastatin are fungal-derived, whereas atorvastatin, cerivastatin, fluvastatin, pravastatin, pitavastatin, and rosuvastatin are fully synthetic compounds [35] . All statins have a similar chemical structure composed of the following: an analogue of the target enzyme

Figure 2. The HMG CoA pathway-inhibition by statins. Inhibition of HMG-CoA by statins inhibits the rate-limiting step in cholesterol biosynthesis, as well as reducing the production of isoprenoid intermediaries. The latter limits the isoprenylation of small GTPase proteins such as Rho, Rac1, and CDC42, and results in modulation of various cellular functions.
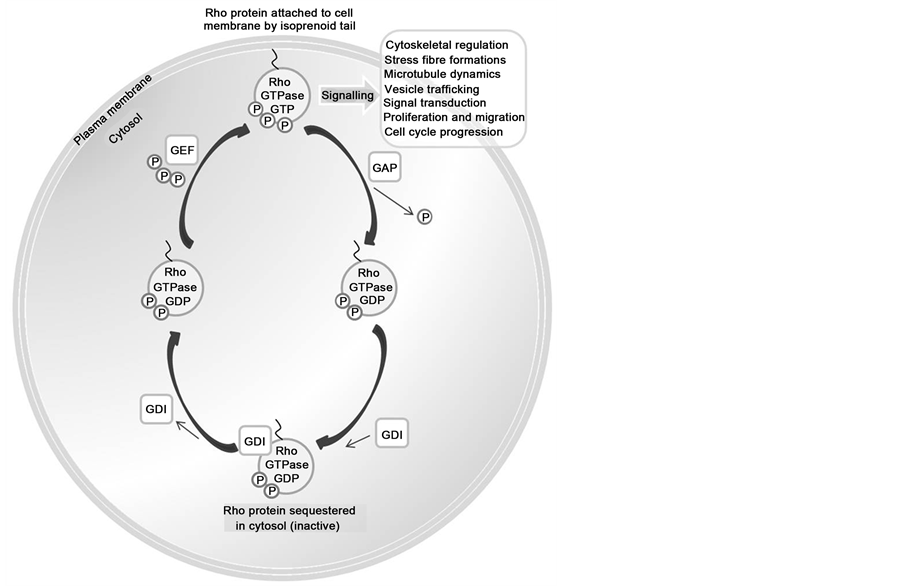
Figure 3. The Rho cycle. GEF: Guanine Exchange Factor. GAP: GTPase-Activating Protein. GDI: Guanine Dissociation Inhibitor. GEF proteins promote the exchange of GDP for GTP on Rho proteins. Binding of GTP facilitates a conformational change in Rho proteins leading to their interaction with effector molecules, and initiation of downstream signalling. GAPs facilitate GTP hydrolysis—so the Rho protein is left bound to GDP. In this state GDIs bind the inactive Rho protein. GDIs prevent membrane translocation of Rho proteins, so they are sequestered in the cytosol.
substrate HMG-CoA; a photophobic ring structure that is involved in binding of the statin to the enzyme; and side groups of the rings that define the solubility properties of the drug [35] . Through examination of this structure the mechanism of action of statins becomes evident—competitive inhibition of HMG-CoA through mimicry of the enzymes substrate.
Much of the pharmacokinetic properties of statins stem from the structure of the side groups—atorvastatin, fluvastatin, lovastatin, and simvastatin are relatively lipophilic compounds, whereas pravastatin and rosuavastatin are more hydrophilic [35] . The absorption of all statins is rapid, and peak plasma concentration is reached within 4 hours after administration [36] - [38] . All statins show great hepatoselectivity with respect to inhibition of HMG-CoA reductase, largely because of efficient first-pass uptake. The bioavailability of statins varies, from 5% for lovastatin, to 60% for pitavastatin [38] [39] . Statins are predominantly metabolised by the cytochrome P450 family of enzymes (CYP450) [40] . The main route for elimination for the majority of statins is via the bile after metabolism by the liver [41] . Statins are highly efficacious at lowering serum low-density lipoprotein (LDL), and can also increase serum high-density lipoprotein (HDL) by varying degrees [42] [43] . The widespread use of statins is now testament to their safety and efficacy. Generally statins are well tolerated with serious adverse effects being very rare [44] . The most serious adverse effect is myopathy, which may progress to rhabdomyolysis. The incidence of myopathy is dose-related, with an approximate incidence of 1 in 1000 patients treated [45] . The risk is increased when statins are used alongside other agents that share common metabolic pathways.
4.2. Application of Statins
The clinical application of statins is rapidly expanding. While prevention of cardiovascular disease is still the primary indication for statins, recent research in the fields of pulmonary and renal disease strongly suggests that statins would also be of benefit in these diseases. Lowering of serum cholesterol is considered to be the main mechanism responsible for the widely recognised beneficial effect of statins in cardiovascular disease [46] - [48] . As well as being of benefit in atherosclerosis and hypercholesterolaemia, statins also improve survival in non- ischaemic heart failure—which is generally unrelated to atherosclerosis [49] . This variant of heart failure has a prominent inflammatory component, and is associated with neurohormonal imbalance—for example activation of the renin-angiotensin-aldosterone and sympathetic system by the low-output state. In vascular cells, angiotensin-II is a key stimulant of reactive oxygen species (ROS) production [50] . Statins are effective in modifying these maladaptive responses, reducing angiotension-II-induced release of ROS, and improving cardiac function in non-ischaemic heart failure [50] - [52] . Recent clinical trials have also demonstrated the benefit of statins as stroke prophylactic agents—reducing the sequelae of stroke by 25% to 35% [53] [54] .
The beneficial effect of statins in pulmonary diseases such as chronic obstructive pulmonary disease (COPD) and idiopathic pulmonary fibrosis (IPF) has recently come to light. Observational studies suggest that patients with COPD who take statins have reduced hospitalisation and mortality rates from COPD exacerbations [55] [56] . The prominent inflammatory and fibrotic components in these lung pathologies bear similarity to the molecular mechanisms observed in the glaucomatous trabecular meshwork and lamina cribrosa. These pathological mechanisms—including abnormal matrix remodelling secondary to enhanced matrix metalloprotease action, increased release of pro-inflammatory cytokines (including TGF-β1, IL-6 and IL-8), dysregulated apoptosis, increased reactive oxygen species—are modulated by statins in in vitro models of pulmonary disease [57] - [60] .
A recent meta-analysis showed that statins are effective in reducing progression in chronic kidney disease [61] . In renal diseases such as renal interstitial fibrosis and diabetic nephropathy, statins have been shown to reduce fibrosis. Statins are capable of inhibiting TGF-β expression and collagen production [62] , but may also reduce fibrosis via a TGF-β independent mechanism [63] . Importantly, recent laboratory evidence shows that statins are effective in inhibiting the pressure-induced fibrotic response of rat renal tubular cells—rosuvastatin reduced the expression of CTGF, TGF-β, fibronectin, and SMAD3 in cells cultured in a high pressure environment [4] . The anti-inflammatory and anti-fibrotic effect of statins was also observed in a model of diabetic nephropathy—rosuvastatin reverses the angiotensin-II-induced upregulation of TGF-β1, fibronectin, and collagen-IV [64] . These studies are very relevant to the field of glaucoma—where differential expression of genes involved in fibrosis and inflammation occurs in the setting of high intraocular pressure [19] .
The anti-fibrotic effect of statins observed in pulmonary and renal disease has prompted innovative research in the area of anti-scarring therapeutics. Treatment of experimental wounds to rabbit ears with local statin injections resulted in reduced hypertrophic scar formation [65] , where mRNA analysis of the tissue revealed significantly lower CTGF expression. Prevention of intra-abdominal scarring and adhesion formation is another emerging therapeutic target. Intra-peritoneal administration of losartan and atorvastatin in mice resulted in reduction of intra-peritoneal bands to a minimum [66] . Statin-containing cellulose film (statofilm) has also been shown to be superior to the established SeprafilmTM in preventing post-operative intraperitoneal adhesion in a rat model, opening up new avenues for future clinical application [67] .
5. Statins and Glaucoma
5.1. Epidemiological Evidence
There are conflicting findings in the literature regarding whether statins are beneficial in patients with POAG. Stein et al conducted a retrospective longitudinal cohort analysis which found that statin use was associated with a significant reduction in the risk of POAG [1] . A dose response effect was also observed—suggesting that longer duration of statin exposure was associated with great reduction in the risk of developing glaucoma or requiring medical treatment for glaucoma. McGwin and colleagues carried out a case control study in male patients which showed that persons who had been prescribed statins for at least 2 years had a 40% reduced odds of developing glaucoma [68] . An interesting study by Leung and co-workers prospectively examined rates of visual field progression in patients who were taking statins versus those who were not. They found that statins were associated with visual field stabilisation in patients with normal tension glaucoma (NTG) [2] . The optic nerve and retinal nerve fibre layer in glaucoma suspects was examined by De Castro and colleagues using the confocal scanning laser ophthalmoscope—and findings suggested that glaucomatous progression was slowed in patients who were taking a statin medication [69] . In contrast to this, other researchers have found no support for the beneficial effect of statins in glaucoma [70] [71] .
By what molecular mechanisms might statins be potentially beneficial in glaucoma? This question opens up many intriguing avenues regarding the pleiotropic effects of statins, and few studies have directly addressed this dilemma to date. Evidence from both human and animal studies has shown that statins have prominent anti-fi- brotic and immune-modulating actions in several organ systems including the eye (Table 1), which may have useful effects in glaucoma. Let us review the known pleiotropic effects of statins and see how these may apply to glaucoma.
5.2. Rho Inhibition
Probably the most important pleiotropic effect of statins is the inhibition of small GTPases, including the Rho family of proteins [77] . The Rho pathway is responsible for various integral intracellular processes and for the interaction between cells and their environment. Three subfamilies constitute the Rho superfamily—Rho, Rac, and Cdc42. Regulation of the actin cytoskeleton, microtubule dynamics, vesicle trafficking, cell polarity and cell-cycle progression are all under the control of the Rho proteins (Figure 4). This topic is comprehensively reviewed by Burridge et al. [34] . The human trabecular meshwork expresses many components of the Rho pathway, suggesting that this is a key player in regulating the contractile tone and cellular morphology of the aqueous outflow pathway [30] [78] . The Rho proteins contain a lipophilic isoprenoid group, which permits their attachment to the cell membrane and is generally essential for biological function. By inhibiting the conversion of HMG-CoA to L-mevalonic acid, statins inhibit the synthesis of isoprenoid intermediaries, including FFP and GGPP. Hence proper subcellular localisation and trafficking of these GTPase proteins is inhibited, with significant functional consequences. Importantly, post-translationally immature forms of G-proteins may maintain partial function [79] [80] , and interfere with the activity of mature membrane-anchored proteins.
By inhibiting Rho and its downstream effector proteins including Rho kinase (ROCK), statins are likely to affect the contractile properties of the conventional outflow pathway. Cells of the conventional pathway possess a contractile tone which is regulated through Rho signaling [81] . ROCK phosphorylates and inhibits the myosin-binding subunit of myosin light chain (MLC) phosphatase. This action increases MLC phosphorylation and myosin contractility, hence driving the formation of stress fibres and focal adhesion [34] . Early work on specific inhibitors of the Rho pathway has shown that Rho inhibition results in relaxation of the contractile tone of cells of the aqueous outflow pathway in vitro and ex vivo [78] [82] [83] . This increases aqueous outflow and reduces intraocular pressure. Indeed Rho kinase inhibitors have been shown to be potent agents in lowering intraocular pressure, and are undergoing phase 2 and 3 clinical trials [31] [84] . This effect of Rho inhibition in lowering the

Table 1. Summary of statin-mediated pharmacological effects on pathological mechanisms in glaucoma.
TGF: Transforming growth factor; CTGF: Connective tissue growth factor; IL: Interleukin; eNOS: Endothelial nitric oxide synthase; iNOS: Inducible nitric oxide synthase; HSP70: Heat shock protein-70; Cav-1: Caveolin-1; tPA: Tissue plasminogen activator; PAI-1: Plasminogen activator inhibitor-1; Ang-II: Angiotensin-II; S-ICAM1: Soluble intercellular adhesion molecule-1; vWF: Von Willibrand factor; CRP: C-reactive protein; TNF: Tumour necrosis factor; TM: Trabecular meshwork.
contractile tone of smooth muscle structures has already proven beneficial in other pathologies—including subarachnoid haemorrhage—where Rho inhibitors have been shown to reduce the incidence of cerebral vasospasm [85] . More recent studies in this field have demonstrated the potential beneficial effect of combination therapy with a statin plus a Rho kinase inhibitor [86] .
Rho inhibitors also affect the actin cytoskeleton, and cellular morphology of the aqueous outflow pathway. In vitro work has shown decreases in actin stress fibres and focal adhesions in cultured porcine and human TM cells [83] . Treatment of monkey Schlemm’s canal (SC) cells with a Rho inhibitor increases the number of giant vacuoles within cell, and decreases the expression of certain cytoskeletal proteins (ZO-1 and claudin-5) [87] . This leads to morphological changes—cell rounding and detachment of cells from each other, as well as wider paracellular spaces [78] [88] . In cultured cells of the SC, Rho inhibition results in increased permeation in vitro, which facilitates aqueous drainage [88] . Ex vivo perfusion experiments, where porcine, monkey, cow and cadaver eyes are perfused with an aqueous humour substitute, have shown that perfusion with a Rho inhibitor increases the conventional outflow facility [78] [83] [88] [89] . To our knowledge, no such experiments have been
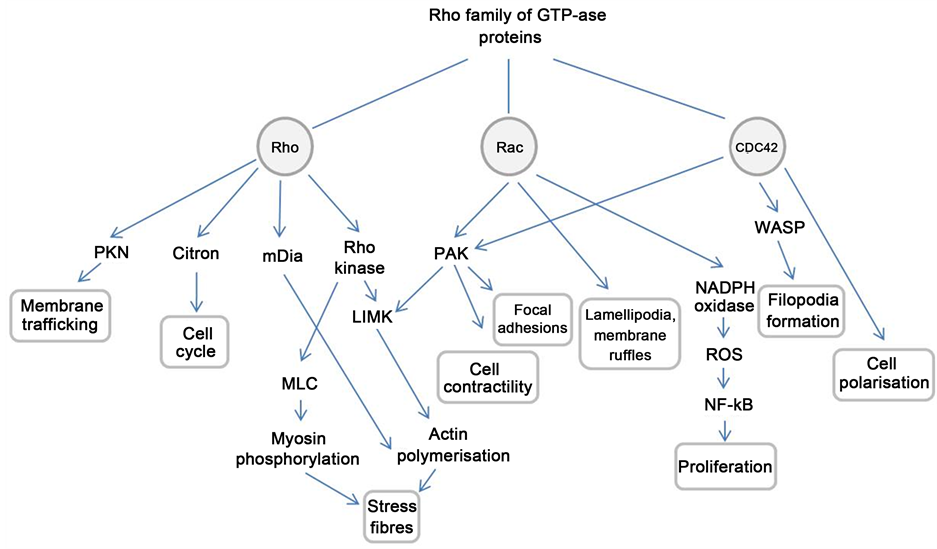
Figure 4. The Rho signalling pathway. PKN: Protein kinase N; mDia: Mammalian diaphanous protein; MLC: Myosin light chain; LIMK: LIM kinase; PAK: p21-activated kinase; NADPH: Nicotinamide adenine dinucleotide phosphate; ROS: Reactive oxygen species; NF-κB: Nuclear factor kappa-light-chain-enhancer of activated B cells; WASP: Wiskott-Aldrich syndrome protein. This diagram illustrates the major effector pathways for the Rho family of proteins (Rho, Rac and CDC-42) which are relevant to glaucoma pathogenesis.
carried out to date with statins, however considering the Rho inhibiting action of statins, it is likely that they have similar results. There is certainly potential for further studies to ascertain the effect of statins themselves on the cell morphology and contractile tone of the aqueous outflow pathway.
5.3. Rac and Reactive Oxygen Species (ROS)
One of the 3 key members of the Rho family is Rac1—this important GTPase protein is responsible for cytoskeletal remodelling—specifically the formation of lamellipodia and membrane ruffles [90] . Lamellipodia are actin-rich cellular protrusions, essential for cell migration, and play an important role in the invasion and metastases of cancer cells [91] . Furthermore, Rac1 binds to p67phox which leads to activation of the NADPH oxidase system and generation of ROS [92] . The presence of high concentrations of ROS can overwhelm the cell’s natural defence mechanisms and lead to programmed cell death. However the role of ROS in cell physiology is more complex thanthat and more recent studies have shown that in some scenarios, ROS (in small doses) promote cell survival—contrary to the traditional view that they are solely destructive molecules [93] [94] . ROS have also been shown to act as signalling molecules in their own right [95] . In smooth muscle and heart cells, it has been shown that by inhibiting the prenylation and subsequent activation of Rac1, statins inactivate NADPH oxidase and hence reduce angiotensin-II-induced ROS production [50] [96] . Our own research group has previously demonstrated evidence of oxidative stress and mitochondrial dysfunction in lamina cribrosa cells from the optic nerve heads of glaucoma donors, compared to normal donors [97] . Furthermore, our group has shown that up to 50% of POAG patients have a pathogenic mitochondrial DNA mutation, which may lead to mitochondrial respiratory dysfunction and subsequent predisposition to oxidative stress in TM, LC and RGC [98] . Increased levels of ROS have been found in the aqueous humour of glaucoma patients [99] [100] . Glutathione is a tripeptide found in the eye and other tissues, and is a key element of the protective mechanism of the eye against oxidative stress [101] . Altered glutathione levels have been reported in the aqueous humour of glaucoma patients [102] , and abnormally low levels of glutathione have even been demonstrated in the serum of glaucoma patients [101] . ROS affect the cellularity of the trabecular meshwork, and may cause endothelial dysfunction— which would contribute to impaired aqueous outflow and higher IOP [103] . Mitochondrial dysfunction in LC and RG cells allows the build-up of ROS, and may lead to increased susceptibility to cell death and impaired repair mechanisms [97] [104] . By reducing the production of ROS in ocular tissues, statins may help to reduce ROS-induced damage and glaucoma progression.
5.4. Endothelium-Derived Nitric Oxide
In the eye, endothelium-derived nitric oxide (NO) is an important regulator of blood flow to the choroid, optic nerve and retina [105] . Since NO is derived from the endothelium, endothelial dysfunction impairs ocular haemodynamics by reducing the production of NO and increasing the production of ROS. Endothelial dysfunction and abnormalities of the L-arginine/nitric oxide system in the vascular tree have been observed in a variety of ocular pathologies, including glaucoma [106] - [108] . Furthermore, studies in humans using laser Doppler flowmetry have demonstrated that inhibition of NO synthase leads to reduced choroidal and optic nerve head blood flow [105] .
Statins influence NO levels by a number of mechanisms. Firstly—the cholesterol lowering effect. Raised serum LDL-cholesterol causes endothelial dysfunction, downregulated endothelial nitric oxide synthase (eNOS) expression, decreased receptor-mediated NO release [109] , and increased ROS production which decreases NO bioavailability [110] . By reducing LDL-cholesterol, statins therefore improve endothelial function and facilitate NO production. Statins also affect NO production through cholesterol independent mechanisms. Statins increase eNOS gene expression by prolonging eNOS mRNA half-life, via a mechanism owing to inhibition of RhoA [111] . Furthermore, statins reduce the bio-availability of the membrane protein Caveolin-1. eNOS binds to Caveolin-1 in caveolae, which results in reduced NO production [112] . Genome wide association studies have suggested Caveolin-1 and -2 mutations in glaucoma [113] . Finally, statins activate the phosphatidylinositol 3-kinase (PI3K)/protein kinase Akt pathway. Akt is an important kinase involved in a number of key cellular functions including cell survival and growth, but it also serves to phosphorylate and activate eNOS—hence facilitating NO production. Statins therefore improve endothelial and vascular function via numerous cholesterol- dependent and -independent mechanisms. Such evidence suggests that statins would be of benefit in improving ocular blood flow in pathologies with compromised vasculature—including glaucoma. By improving endothelial function, statins may also mediate improved function of the endothelium-lined Schlemm’s canal, which has altered biomechanical behaviour in glaucoma [114] .
5.5. The Immune System
Immunomodulation is a well-known pleiotropic effect of statins. This has been shown to be beneficial in several pathologies where the immune system is upregulated, including multiple sclerosis and systemic lupus erythematosus [115] . Antigen presentation requires endocytosis of the antigen, internal processing, and presentation of major histocompatibility complex (MHC) II molecules at the cell surface. MHC II has been shown to be upregulated in a mouse model of glaucoma [116] . Furthermore, in a rat model of glaucoma microglial activation in the optic nerve correlated with the degree of axonal damage, and both MHC I and II were persistently upregulated in the microglia of optic nerves in the rat glaucoma model [117] . Gene expression of cytokines, including IL-6, have been shown to be upregulated in rat optic nerve heads subjected to raised intraocular pressure [118] . Many of the immunomodulatory effects of statins occur via the inhibition of small GTPase proteins [50] [77] . Antigen presentation and MHC II expression is related to changes in the actin cytoskeleton—which is controlled by the Rho pathway. Statins also decrease macrophage expression of tumour necrosis factor and interleukin-1β [75] .
6. Effect of Statins on Ocular Tissues
Novel work by Villarreal et al. at the Massachusetts Eye and Ear Infirmary is one of the few to examine the molecular mechanisms involved in ocular tissues treated with statins [74] . In this project, human trabecular meshwork cells were treated with lovastatin in vitro, and SPARC mRNA and protein expression were determined. SPARC is a matricellular protein—and a critical mediator of aqueous outflow and IOP [119] - [121] . It is associated with increased tissue fibrosis and aberrant tissue remodelling, and it has been shown to be upregulated in TM cells following TGF-β2 treatment [122] . Statin treatment suppressed SPARC expression in TM cells— supporting the potential therapeutic role for statins in glaucoma.
The Rho signalling pathway has also been examined in TM cells treated with statins. Stubbs et al. showed that lovastatin increases the accumulation of unprenylated forms of RhoA and RhoB in human TM cells [123] . This may disrupt the Rho-dependent regulation of TM cell cytoskeletal organisation. Treatment of human TM cells with lovastatin resulted in disruption of filamentous actin organisation, an effect which was reversed by the supplementation of the cell cultures with GGPP.
Krempler et al. conducted novel research into the effect of statins on retinal ganglion cell survival and visual function following acute retinal ischaemia/reperfusion in mice [76] . Treatment of the animals with statins after the ischaemic insult resulted not only in histological evidence of improved retinal ganglion cell survival, but also improved visual function as measured behaviourally (by visual acuity and contrast sensitivity). This technique of producing acute global ischaemia represents a model of the changes observed after central retinal artery occlusion, as well as acute angle closure glaucoma [124] . This work hence supports the role for statins as a useful pharmacological tool in various ocular pathologies associated with ischaemic damage.
7. Concluding Remarks
The beneficial effect of statins extends far beyond cholesterol-lowering. Their pleiotropic effects include anti- fibrotic, anti-inflammatory, and immunomodulatory action, many of which are mediated by inhibition of the Rho pathway. These effects are evident in a multitude of human tissues, and statins as pleiotropic agents are already being studied in the fields of cardiovascular, pulmonary, and renal disease. Now, statins have caught the eye of glaucoma scholars as an intriguing new therapeutic agent, opening up many new avenues for molecular and clinical research.
References
- Stein, J.D., et al. (2012) The Relationship between Statin Use and Open-Angle Glaucoma. Ophthalmology, 119, 2074- 2081. http://dx.doi.org/10.1016/j.ophtha.2012.04.029
- Leung, D.Y., et al. (2010) Simvastatin and Disease Stabilization in Normal Tension Glaucoma: A Cohort Study. Ophthalmology, 117, 471-476. http://dx.doi.org/10.1016/j.ophtha.2009.08.016
- Schmeer, C., Kretz, A. and Isenmann, S. (2006) Statin-Mediated Protective Effects in the Central Nervous System: General Mechanisms and Putative Role of Stress Proteins. Restorative Neurology and Neuroscience, 24, 79-95.
- Chen, C.H., et al. (2013) Rosuvastatin Inhibits Pressure-Induced Fibrotic Responses via the Expression Regulation of Prostacyclin and Prostaglandin E2 in Rat Renal Tubular Cells. European Journal of Pharmacology, 700, 65-73. http://dx.doi.org/10.1016/j.ejphar.2012.12.017
- Schaafsma, D., et al. (2011) Simvastatin Inhibits TGFbeta1-Induced Fibronectin in Human Airway Fibroblasts. Respiratory Research, 12, 113. http://dx.doi.org/10.1186/1465-9921-12-113
- Watts, K.L., et al. (2005) Simvastatin Inhibits Growth Factor Expression and Modulates Profibrogenic Markers in Lung Fibroblasts. American Journal of Respiratory Cell and Molecular Biology, 32, 290-300. http://dx.doi.org/10.1165/rcmb.2004-0127OC
- Cordle, A., et al. (2005) Mechanisms of Statin-Mediated Inhibition of Small G-Protein Function. Journal of Biological Chemistry, 280, 34202-34209. http://dx.doi.org/10.1074/jbc.M505268200
- Laufs, U., et al. (2000) Neuroprotection Mediated by Changes in the Endothelial Actin Cytoskeleton. Journal of Clinical Investigation, 106, 15-24. http://dx.doi.org/10.1172/JCI9639
- Kato, T., et al. (2004) Statin Blocks Rho/Rho-Kinase Signalling and Disrupts the Actin Cytoskeleton: Relationship to Enhancement of LPS-Mediated Nitric Oxide Synthesis in Vascular Smooth Muscle Cells. Biochimica et Biophysica Acta, 1689, 267-272. http://dx.doi.org/10.1016/j.bbadis.2004.04.006
- Quigley, H.A., et al. (1981) Optic Nerve Damage in Human Glaucoma. II. The Site of Injury and Susceptibility to Damage. Archives of Ophthalmology, 99, 635-649. http://dx.doi.org/10.1001/archopht.1981.03930010635009
- Quigley,
- Lambert, W., et al. (2001) Neurotrophin and Neurotrophin Receptor Expression by Cells of the Human Lamina Cribrosa. Investigative Ophthalmology & Visual Science, 42, 2315-2323.
- Cassard, S.D., et al. (2012) Regional Variations and Trends in the Prevalence of Diagnosed Glaucoma in the Medicare Population. Ophthalmology, 119, 1342-1351. http://dx.doi.org/10.1016/j.ophtha.2012.01.032
-
-
-
- Fautsch, M.P. and Johnson, D.H. (2006) Aqueous Humor Outflow: What Do We Know? Where Will It Lead Us? Investigative Ophthalmology & Visual Science, 47, 4181-4187. http://dx.doi.org/10.1167/iovs.06-0830
- Johnson, M. (2006) What Controls Aqueous Humour Outflow Resistance? Experimental Eye Research, 82, 545-557. http://dx.doi.org/10.1016/j.exer.2005.10.011
- Kirwan, R.P., et al. (2009) Differential Global and Extra-Cellular Matrix Focused Gene Expression Patterns between Normal and Glaucomatous Human Lamina Cribrosa Cells. Molecular Vision, 15, 76-88.
- Liu, Y., et al. (2013) Gene Expression Profile in Human Trabecular Meshwork with Primary Open Angle Glaucoma. Investigative Ophthalmology & Visual Science, 54, 6382-6389.
- Coleman, A.L., et al. (1991) Displacement of the Optic Nerve Head by Acute Changes in Intraocular Pressure in Monkey Eyes. Ophthalmology, 98, 35-40. http://dx.doi.org/10.1016/S0161-6420(91)32345-5
- Hernandez, M.R. (1992) Ultrastructural Immunocytochemical Analysis of Elastin in the Human Lamina Cribrosa. Changes in Elastic Fibers in Primary Open-Angle Glaucoma. Investigative Ophthalmology & Visual Science, 33, 2891- 2903.
- Quigley, H.A., et al. (2000) Retrograde Axonal Transport of BDNF in Retinal Ganglion Cells Is Blocked by Acute IOP Elevation in Rats. Investigative Ophthalmology & Visual Science, 41, 3460-3466.
- Hernandez, M.R., Igoe, F. and Neufeld, A.H. (1988) Cell Culture of the Human Lamina Cribrosa. Investigative Ophthalmology & Visual Science, 29, 78-89.
- Kirwan, R.P., et al. (2005) Influence of Cyclical Mechanical Strain on Extracellular Matrix Gene Expression in Human Lamina Cribrosa Cells in Vitro. Molecular Vision, 11, 798-810.
- Kirwan, R.P., et al. (2005) Transforming Growth Factor-Beta-Regulated Gene Transcription and Protein Expression in Human GFAP-Negative Lamina Cribrosa Cells. Glia, 52, 309-324. http://dx.doi.org/10.1002/glia.20247
- Quill, B., et al. (2011) The Effect of Graded Cyclic Stretching on Extracellular Matrix-Related Gene Expression Profiles in Cultured Primary Human Lamina Cribrosa Cells. Investigative Ophthalmology & Visual Science, 52, 1908- 1915. http://dx.doi.org/10.1167/iovs.10-5467
- Wallace, D.M., et al. (2013) Anti-Connective Tissue Growth Factor Antibody Treatment Reduces Extracellular Matrix Production in Trabecular Meshwork and Lamina Cribrosa Cells. Investigative Ophthalmology & Visual Science, 54, 7836-7848. http://dx.doi.org/10.1167/iovs.13-12494
- Pattabiraman, P.P., Maddala, R. and Rao, P.V. (2014) Regulation of Plasticity and Fibrogenic Activity of Trabecular Meshwork Cells by Rho GTPase Signaling. Journal of Cellular Physiology, 229, 927-942. http://dx.doi.org/10.1002/jcp.24524
- Honjo, M., et al. (2001) Effects of Rho-Associated Protein Kinase Inhibitor Y-27632 on Intraocular Pressure and Outflow Facility. Investigative Ophthalmology & Visual Science, 42, 137-144.
- Tanihara, H., et al. (2013) Phase 2 Randomized Clinical Study of a Rho Kinase Inhibitor, K-115, in Primary Open- Angle Glaucoma and Ocular Hypertension. American Journal of Ophthalmology, 156, 731-736. http://dx.doi.org/10.1016/j.ajo.2013.05.016
- Goldstein, J.L. and Brown, M.S. (1990) Regulation of the Mevalonate Pathway. Nature, 343, 425-430. http://dx.doi.org/10.1038/343425a0
- Van Aelst, L. and D’Souza-Schorey, C. (1997) Rho GTPases and Signaling Networks. Genes Development, 11, 2295- 2322. http://dx.doi.org/10.1101/gad.11.18.2295
- Burridge, K. and Wennerberg, K. (2004) Rho and Rac Take Center Stage. Cell, 116, 167-179. http://dx.doi.org/10.1016/S0092-8674(04)00003-0
- Gaw, A. (2000) Statins: The HMG CoA Reductase Inhibitors in Perspective. Martin Dunitz Ltd.
- Cilla Jr., D.D., et al. (1996) Pharmacodynamic Effects and Pharmacokinetics of Atorvastatin after Administration to Normocholesterolemic Subjects in the Morning and Evening. The Journal of Clinical Pharmacology, 36, 604-609. http://dx.doi.org/10.1002/j.1552-4604.1996.tb04224.x
- Tse, F.L., Jaffe, J.M. and Troendle, A. (1992) Pharmacokinetics of Fluvastatin after Single and Multiple Doses in Normal Volunteers. The Journal of Clinical Pharmacology, 32, 630-638. http://dx.doi.org/10.1002/j.1552-4604.1992.tb05773.x
- Pan, H.Y., et al. (1990) Comparative Pharmacokinetics and Pharmacodynamics of Pravastatin and Lovastatin. The Journal of Clinical Pharmacology, 30, 1128-1135. http://dx.doi.org/10.1002/j.1552-4604.1990.tb01856.x
- Kajinami, K., Mabuchi, H. and Saito, Y. (2000) NK-104: A Novel Synthetic HMG-CoA Reductase Inhibitor. Expert Opinion on Investigational Drugs, 9, 2653-2661. http://dx.doi.org/10.1517/13543784.9.11.2653
- Bottorff, M. and Hansten, P. (2000) Long-Term Safety of Hepatic Hydroxymethyl Glutaryl Coenzyme A Reductase Inhibitors: The Role of Metabolism-Monograph for Physicians. Archives of Internal Medicine, 160, 2273-2280. http://dx.doi.org/10.1001/archinte.160.15.2273
- Knopp, R.H. (1999) Drug Treatment of Lipid Disorders. The New England Journal of Medicine, 341, 498-511. http://dx.doi.org/10.1056/NEJM199908123410707
- McKenney, J.M., et al. (2003) Comparison of the Efficacy of Rosuvastatin versus Atorvastatin, Simvastatin, and Pravastatin in Achieving Lipid Goals: Results from the STELLAR Trial. Current Medical Research and Opinion, 19, 689- 698. http://dx.doi.org/10.1185/030079903125002405
- Olsson, A.G., et al. (2001) Effect of Rosuvastatin on Low-Density Lipoprotein Cholesterol in Patients with Hypercholesterolemia. American Journal of Cardiology, 88, 504-508. http://dx.doi.org/10.1016/S0002-9149(01)01727-1
- Black, D.M. (2002) A General Assessment of the Safety of HMG CoA Reductase Inhibitors (Statins). Current Atherosclerosis Reports, 4, 34-41. http://dx.doi.org/10.1007/s11883-002-0060-0
- Ballantyne, C.M., et al. (2003) Risk for Myopathy with Statin Therapy in High-Risk Patients. Archives of Internal Medicine, 163, 553-564. http://dx.doi.org/10.1001/archinte.163.5.553
- Scandinavian Simvastatin Survival Study Group (1994) Randomised Trial of Cholesterol Lowering in 4444 Patients with Coronary Heart Disease: The Scandinavian Simvastatin Survival Study (4S). The Lancet, 344, 1383-1389.
- Shepherd, J., et al. (1995) Prevention of Coronary Heart Disease with Pravastatin in Men with Hypercholesterolemia. West of Scotland Coronary Prevention Study Group. The New England Journal of Medicine, 333, 1301-1307. http://dx.doi.org/10.1056/NEJM199511163332001
- Heart Protection Study Collaborative Group (2002) MRC/BHF Heart Protection Study of Cholesterol Lowering with Simvastatin in 20,536 High-Risk Individuals: A Randomised Placebo-Controlled Trial. The Lancet, 360, 7-22. http://dx.doi.org/10.1016/S0140-6736(02)09327-3
- Horwich, T.B., MacLellan, W.R. and Fonarow, G.C. (2004) Statin Therapy Is Associated with Improved Survival in Ischemic and Non-Ischemic Heart Failure. Journal of the American College of Cardiology, 43, 642-648. http://dx.doi.org/10.1016/j.jacc.2003.07.049
- Wassmann, S., et al. (2001) Inhibition of Geranylgeranylation Reduces Angiotensin II-Mediated Free Radical Production in Vascular Smooth Muscle Cells: Involvement of Angiotensin AT1 Receptor Expression and Rac1 GTPase. Molecular Pharmacology, 59, 646-654.
- Liu, M., et al. (2009) Atorvastatin Improves Endothelial Function and Cardiac Performance in Patients with Dilated Cardiomyopathy: The Role of Inflammation. Cardiovascular Drugs and Therapy, 23, 369-376. http://dx.doi.org/10.1007/s10557-009-6186-3
- Landmesser, U., et al. (2005) Simvastatin versus Ezetimibe: Pleiotropic and Lipid-Lowering Effects on Endothelial Function in Humans. Circulation, 111, 2356-2363. http://dx.doi.org/10.1161/01.CIR.0000164260.82417.3F
- Gil-Nunez, A.C. and Villanueva, J.A. (2001) Advantages of Lipid-Lowering Therapy in Cerebral Ischemia: Role of HMG-CoA Reductase Inhibitors. Cerebrovascular Diseases, 11, 85-95. http://dx.doi.org/10.1159/000049130
- Vaughan, C.J., Delanty, N. and Basson, C.T. (2001) Statin Therapy and Stroke Prevention. Current Opinion in Cardiology, 16, 219-224. http://dx.doi.org/10.1097/00001573-200107000-00001
- Mancini, G.B., et al. (2006) Reduction of Morbidity and Mortality by Statins, Angiotensin-Converting Enzyme Inhibitors, and Angiotensin Receptor Blockers in Patients with Chronic Obstructive Pulmonary Disease. Journal of the American College of Cardiology, 47, 2554-2560. http://dx.doi.org/10.1016/j.jacc.2006.04.039
- Soyseth, V., et al. (2007) Statin Use Is Associated with Reduced Mortality in COPD. European Respiratory Journal, 29, 279-283. http://dx.doi.org/10.1183/09031936.00106406
- Murphy, D.M., et al. (2008) Simvastatin Attenuates Release of Neutrophilic and Remodeling Factors from Primary Bronchial Epithelial Cells Derived from Stable Lung Transplant Recipients. American Journal of Physiology-Lung Cellular and Molecular Physiology, 294, L592-L599. http://dx.doi.org/10.1152/ajplung.00386.2007
- Ito, T., et al. (2002) Regulation of Interleukin-8 Expression by HMG-CoA Reductase Inhibitors in Human Vascular Smooth Muscle Cells. Atherosclerosis, 165, 51-55. http://dx.doi.org/10.1016/S0021-9150(02)00194-6
- Newton, C.J., et al. (2002) Statin-Induced Apoptosis of Vascular Endothelial Cells Is Blocked by Dexamethasone. Journal of Endocrinology, 174, 7-16. http://dx.doi.org/10.1677/joe.0.1740007
- Shishehbor, M.H., et al. (2003) Statins Promote Potent Systemic Antioxidant Effects through Specific Inflammatory Pathways. Circulation, 108, 426-431. http://dx.doi.org/10.1161/01.CIR.0000080895.05158.8B
- Geng, Q., et al. (2014) Meta-Analysis of the Effect of Statins on Renal Function. American Journal of Cardiology, 114, 562-570. http://dx.doi.org/10.1016/j.amjcard.2014.05.033
- Li, Y.B., et al. (2011) Effect of Simvastatin on Expression of Transforming Growth Factor-Beta and Collagen Type IV in Rat Mesangial Cells. Pharmacology, 88, 188-192. http://dx.doi.org/10.1159/000330739
- Rodrigues Diez, R., et al. (2010) Statins Inhibit Angiotensin II/Smad Pathway and Related Vascular Fibrosis, by a TGF-Beta-Independent Process. PLoS ONE, 5, e14145. http://dx.doi.org/10.1371/journal.pone.0014145
- Solini, A., et al. (2011) Angiotensin-II and Rosuvastatin Influence Matrix Remodeling in Human Mesangial Cells via Metalloproteinase Modulation. Journal of Hypertension, 29, 1930-1939. http://dx.doi.org/10.1097/HJH.0b013e32834abceb
- Ko, J.H., et al. (2012) HMG-CoA Reductase Inhibitors (Statins) Reduce Hypertrophic Scar Formation in a Rabbit Ear Wounding Model. Plastic and Reconstructive Surgery, 129, 252e-261e. http://dx.doi.org/10.1097/PRS.0b013e31823aea10
- Dinarvand, P., et al. (2013) Novel Approach to Reduce Postsurgical Adhesions to a Minimum: Administration of Losartan plus Atorvastatin Intraperitoneally. Journal of Surgical Research, 181, 91-98. http://dx.doi.org/10.1016/j.jss.2012.05.035
- Lalountas, M., et al. (2012) Postoperative Adhesion Prevention Using a Statin-Containing Cellulose Film in an Experimental Model. British Journal of Surgery, 99, 423-429. http://dx.doi.org/10.1002/bjs.7817
- McGwin Jr., G., et al. (2004) Statins and Other Cholesterol-Lowering Medications and the Presence of Glaucoma. Archives of Ophthalmology, 122, 822-826. http://dx.doi.org/10.1001/archopht.122.6.822
- De Castro, D.K., et al. (2007) Effect of Statin Drugs and Aspirin on Progression in Open-Angle Glaucoma Suspects Using Confocal Scanning Laser Ophthalmoscopy. Clinical & Experimental Ophthalmology, 35, 506-513. http://dx.doi.org/10.1111/j.1442-9071.2007.01529.x
- Owen, C.G., et al. (2010) Hypotensive Medication, Statins, and the Risk of Glaucoma. Investigative Ophthalmology & Visual Science, 51, 3524-3530. http://dx.doi.org/10.1167/iovs.09-4821
- Iskedjian, M., et al. (2009) Effect of Selected Antihypertensives, Antidiabetics, Statins and Diuretics on Adjunctive Medical Treatment of Glaucoma: A Population Based Study. Current Medical Research and Opinion, 25, 1879-1888. http://dx.doi.org/10.1185/03007990903035083
- Garcia, I.M., et al. (2012) Caveolin-1-eNOS/Hsp70 Interactions Mediate Rosuvastatin Antifibrotic Effects in Neonatal Obstructive Nephropathy. Nitric Oxide, 27, 95-105. http://dx.doi.org/10.1016/j.niox.2012.05.006
- Tang, X.L., et al. (2011) Atorvastatin Therapy during the Peri-Infarct Period Attenuates Left Ventricular Dysfunction and Remodeling after Myocardial Infarction. PLoS ONE, 6, e25320. http://dx.doi.org/10.1371/journal.pone.0025320
- Villarreal Jr., G., et al. (2014) Pharmacological Regulation of SPARC by Lovastatin in Human Trabecular Meshwork Cells. Investigative Ophthalmology & Visual Science, 55, 1657-1665. http://dx.doi.org/10.1167/iovs.13-12712
- Pahan, K., et al. (1997) Lovastatin and Phenylacetate Inhibit the Induction of Nitric Oxide Synthase and Cytokines in Rat Primary Astrocytes, Microglia, and Macrophages. Journal of Clinical Investigation, 100, 2671-2679. http://dx.doi.org/10.1172/JCI119812
- Krempler, K., et al. (2011) Simvastatin Improves Retinal Ganglion Cell Survival and Spatial Vision after Acute Retinal Ischemia/Reperfusion in Mice. Investigative Ophthalmology & Visual Science, 52, 2606-2618. http://dx.doi.org/10.1167/iovs.10-6005
- Tanaka, S., et al. (2013) Statins Exert the Pleiotropic Effects through Small GTP-Binding Protein Dissociation Stimulator Upregulation with a Resultant Rac1 Degradation. Arteriosclerosis, Thrombosis, and Vascular Biology, 33, 1591- 1600. http://dx.doi.org/10.1161/ATVBAHA.112.300922
- Rao, P.V., et al. (2001) Modulation of Aqueous Humor Outflow Facility by the Rho Kinase-Specific Inhibitor Y- 27632. Investigative Ophthalmology & Visual Science, 42, 1029-1037.
- Allal, C., et al. (2000) RhoA Prenylation Is Required for Promotion of Cell Growth and Transformation and Cytoskeleton Organization but Not for Induction of Serum Response Element Transcription. The Journal of Biological Chemistry, 275, 31001-31008. http://dx.doi.org/10.1074/jbc.M005264200
- Lebowitz, P.F., Du, W. and Prendergast, G.C. (1997) Prenylation of RhoB Is Required for Its Cell Transforming Function but Not Its Ability to Activate Serum Response Element-Dependent Transcription. The Journal of Biological Chemistry, 272, 16093-16095. http://dx.doi.org/10.1074/jbc.272.26.16093
- Ramos, R.F., Sumida, G.M. and Stamer, W.D. (2009) Cyclic Mechanical Stress and Trabecular Meshwork Cell Contractility. Investigative Ophthalmology & Visual Science, 50, 3826-3832. http://dx.doi.org/10.1167/iovs.08-2694
- Honjo, M., et al. (2001) Effects of Protein Kinase Inhibitor, HA1077, on Intraocular Pressure and Outflow Facility in Rabbit Eyes. Archives of Ophthalmology, 119, 1171-8. http://dx.doi.org/10.1001/archopht.119.8.1171
- Rao, P.V., et al. (2005) Expression of Dominant Negative Rho-Binding Domain of Rho-Kinase in Organ Cultured Human Eye Anterior Segments Increases Aqueous Humor Outflow. Molecular Vision, 11, 288-297.
- Tanihara, H., et al. (2013) Phase 1 Clinical Trials of a Selective Rho Kinase Inhibitor, K-115. JAMA Ophthalmology, 131, 1288-1295. http://dx.doi.org/10.1001/jamaophthalmol.2013.323
- Takanashi, Y., et al. (2001) Efficacy of Intrathecal Liposomal Fasudil for Experimental Cerebral Vasospasm after Subarachnoid Hemorrhage. Neurosurgery, 48, 894-900.
- Naraoka, M., et al. (2013) Suppression of the Rho/Rho-Kinase Pathway and Prevention of Cerebral Vasospasm by Combination Treatment with Statin and Fasudil after Subarachnoid Hemorrhage in Rabbit. Translational Stroke Research, 4, 368-374. http://dx.doi.org/10.1007/s12975-012-0247-9
- Thieme, H., et al. (2000) Mediation of Calcium-Independent Contraction in Trabecular Meshwork through Protein Kinase C and Rho-A. Investigative Ophthalmology & Visual Science, 41, 4240-4246.
- Kameda, T., et al. (2012) The Effect of Rho-Associated Protein Kinase Inhibitor on Monkey Schlemm’s Canal Endothelial Cells. Investigative Ophthalmology & Visual Science, 53, 3092-3103.
- Lu, Z., et al. (2008) The Mechanism of Increasing Outflow Facility by Rho-Kinase Inhibition with Y-27632 in Bovine Eyes. Experimental Eye Research, 86, 271-281. http://dx.doi.org/10.1016/j.exer.2007.10.018
- Nobes, C.D. and Hall, A. (1995) Rho, Rac, and Cdc42 GTPases Regulate the Assembly of Multimolecular Focal Complexes Associated with Actin Stress Fibers, Lamellipodia, and Filopodia. Cell, 81, 53-62. http://dx.doi.org/10.1016/0092-8674(95)90370-4
- Kato, T., et al. (2014) Rac1-Dependent Lamellipodial Motility in Prostate Cancer PC-3 Cells Revealed by Optogenetic Control of Rac1 Activity. PLoS ONE, 9, e97749. http://dx.doi.org/10.1371/journal.pone.0097749
- Sundaresan, M., et al. (1996) Regulation of Reactive-Oxygen-Species Generation in Fibroblasts by Rac1. Biochemical Journal, 318, 379-382.
- Farrell, S.M., et al. (2011) bFGF-Mediated Redox Activation of the PI3K/Akt Pathway in Retinal Photoreceptor Cells. European Journal of Neuroscience, 33, 632-641. http://dx.doi.org/10.1111/j.1460-9568.2010.07559.x
- Groeger, G., Quiney, C. and Cotter, T.G. (2009) Hydrogen Peroxide as a Cell-Survival Signaling Molecule. Antioxidants Redox Signaling, 11, 2655-2671. http://dx.doi.org/10.1089/ars.2009.2728
- Rhee, S.G. (2006) Cell Signaling. H2O2, a Necessary Evil for Cell Signaling. Science, 312, 1882-1883. http://dx.doi.org/10.1126/science.1130481
- Takemoto, M., et al. (2001) Statins as Antioxidant Therapy for Preventing Cardiac Myocyte Hypertrophy. Journal of Clinical Investigation, 108, 1429-1437. http://dx.doi.org/10.1172/JCI13350
- McElnea, E.M., et al. (2011) Oxidative Stress, Mitochondrial Dysfunction and Calcium Overload in Human Lamina Cribrosa Cells from Glaucoma Donors. Molecular Vision, 17, 1182-1191.
- Sundaresan, P., et al. (2014) Whole-Mitochondrial Genome Sequencing in Primary Open-Angle Glaucoma Using Massively Parallel Sequencing Identifies Novel and Known Pathogenic Variants. Genetics in Medicine, Published Online. http://dx.doi.org/10.1038/gim.2014.121
- Schlotzer-Schrehardt, U. (2010) Oxidative Stress and Pseudoexfoliation Glaucoma. Klinische Monatsblätter für Augenheilkunde, 227, 108-113. http://dx.doi.org/10.1055/s-0028-1109977
- Sacca, S.C. and Izzotti, A. (2008) Oxidative Stress and Glaucoma: Injury in the Anterior Segment of the Eye. Progress in Brain Research, 173, 385-407. http://dx.doi.org/10.1016/S0079-6123(08)01127-8
- Gherghel, D., et al. (2005) Systemic Reduction in Glutathione Levels Occurs in Patients with Primary Open-Angle Glaucoma. Investigative Ophthalmology & Visual Science, 46, 877-883. http://dx.doi.org/10.1167/iovs.04-0777
- Ferreira, S.M., et al. (2004) Oxidative Stress Markers in Aqueous Humor of Glaucoma Patients. American Journal of Ophthalmology, 137, 62-69. http://dx.doi.org/10.1016/S0002-9394(03)00788-8
- Sacca, S.C., et al. (2007) Glaucomatous Outflow Pathway and Oxidative Stress. Experimental Eye Research, 84, 389- 399. http://dx.doi.org/10.1016/j.exer.2006.10.008
- Maher, P. and Hanneken, A. (2005) The Molecular Basis of Oxidative Stress-Induced Cell Death in an Immortalized Retinal Ganglion Cell Line. Investigative Ophthalmology & Visual Science, 46, 749-757. http://dx.doi.org/10.1167/iovs.04-0883
- Luksch, A., et al. (2000) Effects of Systemic NO Synthase Inhibition on Choroidal and Optic Nerve Head Blood Flow in Healthy Subjects. Investigative Ophthalmology & Visual Science, 41, 3080-3084.
- Polak, K., et al. (2007) Altered Nitric Oxide System in Patients with Open-Angle Glaucoma. Archives of Ophthalmology, 125, 494-498. http://dx.doi.org/10.1001/archopht.125.4.494
- Henry, E., et al. (2006) Altered Endothelin-1 Vasoreactivity in Patients with Untreated Normal-Pressure Glaucoma. Investigative Ophthalmology & Visual Science, 47, 2528-2532. http://dx.doi.org/10.1167/iovs.05-0240
- Henry, E., et al. (1999) Peripheral Endothelial Dysfunction in Normal Pressure Glaucoma. Investigative Ophthalmology & Visual Science, 40, 1710-1714.
- Liao, J.K. (1994) Inhibition of Gi Proteins by Low Density Lipoprotein Attenuates Bradykinin-Stimulated Release of Endothelial-Derived Nitric Oxide. The Journal of Biological Chemistry, 269, 12987-12992.
- Alderson, L.M., et al. (1986) LDL Enhances Monocyte Adhesion to Endothelial Cells in Vitro. The American Journal of Pathology, 123, 334-342.
- Laufs, U. and Liao, J.K. (1998) Post-Transcriptional Regulation of Endothelial Nitric Oxide Synthase mRNA Stability by Rho GTPase. The Journal of Biological Chemistry, 273, 24266-24271. http://dx.doi.org/10.1074/jbc.273.37.24266
- Plenz, G.A., Hofnagel, O. and Robenek, H. (2004) Differential Modulation of Caveolin-1 Expression in Cells of the Vasculature by Statins. Circulation, 109, e7-e8.
- Thorleifsson, G., et al. (2010) Common Variants near CAV1 and CAV2 Are Associated with Primary Open-Angle Glaucoma. Nature Genetics, 42, 906-909. http://dx.doi.org/10.1038/ng.661
- Overby, D.R., et al. (2014) Altered Mechanobiology of Schlemm’s Canal Endothelial Cells in Glaucoma. Proceedings of the National Academy of Sciences of the United States of America, 111, 13876-13881. http://dx.doi.org/10.1073/pnas.1410602111
- Ulivieri, C. and Baldari, C.T. (2014) Statins: From Cholesterol-Lowering Drugs to Novel Immunomodulators for the Treatment of Th17-Mediated Autoimmune Diseases. Pharmacological Research, 88, 41-52. http://dx.doi.org/10.1016/j.phrs.2014.03.001
- Gallego, B.I., et al. (2012) IOP Induces Upregulation of GFAP and MHC-II and Microglia Reactivity in Mice Retina Contralateral to Experimental Glaucoma. Journal of Neuroinflammation, 9, 92. http://dx.doi.org/10.1186/1742-2094-9-92
- Ebneter, A., et al. (2010) Microglial Activation in the Visual Pathway in Experimental Glaucoma: Spatiotemporal Characterization and Correlation with Axonal Injury. Investigative Ophthalmology & Visual Science, 51, 6448-6460. http://dx.doi.org/10.1167/iovs.10-5284
- Johnson, E.C., et al. (2011) Cell Proliferation and Interleukin-6-Type Cytokine Signaling Are Implicated by Gene Expression Responses in Early Optic Nerve Head Injury in Rat Glaucoma. Investigative Ophthalmology & Visual Science, 52, 504-518. http://dx.doi.org/10.1167/iovs.10-5317
- Haddadin, R.I., et al. (2009) SPARC-Null Mice Exhibit Lower Intraocular Pressures. Investigative Ophthalmology & Visual Science, 50, 3771-3777. http://dx.doi.org/10.1167/iovs.08-2489
- Swaminathan, S.S., et al. (2013) Secreted Protein Acidic and Rich in Cysteine (SPARC)-Null Mice Exhibit More Uniform Outflow. Investigative Ophthalmology & Visual Science, 54, 2035-2047. http://dx.doi.org/10.1167/iovs.12-10950
- Oh, D.J., et al. (2013) Overexpression of SPARC in Human Trabecular Meshwork Increases Intraocular Pressure and Alters Extracellular Matrix. Investigative Ophthalmology & Visual Science, 54, 3309-3319. http://dx.doi.org/10.1167/iovs.12-11362
- Bollinger, K.E., et al. (2011) Quantitative Proteomics: TGFbeta(2) Signaling in Trabecular Meshwork Cells. Investigative Ophthalmology & Visual Science, 52, 8287-8294. http://dx.doi.org/10.1167/iovs.11-8218
- Von Zee, C.L., et al. (2009) Increased RhoA and RhoB Protein Accumulation in Cultured Human Trabecular Meshwork Cells by Lovastatin. Investigative Ophthalmology & Visual Science, 50, 2816-2823. http://dx.doi.org/10.1167/iovs.08-2466
- Osborne, N.N., et al. (2004) Retinal Ischemia: Mechanisms of Damage and Potential Therapeutic Strategies. Progress in Retinal and Eye Research, 23, 91-147. http://dx.doi.org/10.1016/j.preteyeres.2003.12.001