Open Journal of Marine Science
Vol.3 No.2A(2013), Article ID:33668,12 pages DOI:10.4236/ojms.2013.32A005
Long-Term Effect of Photoperiod, Temperature and Feeding Regimes on the Respiration Rates of Antarctic Krill (Euphausia superba)
1Institute of Antarctic and Southern Ocean Studies, University of Tasmania, Hobart, Australia
2Australian Antarctic Division, Kingston, Australia
3Antarctic Climate and Ecosystems Cooperative Research Centre, Hobart, Australia
4Institute for Antarctic and Marine Studies, University of Tasmania, Hobart, Australia
Email: *So.Kawaguchi@aad.gov.au
Copyright © 2013 Matthew Brown et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received April 1, 2013; revised May 16, 2013; accepted June 30, 2013
Keywords: Antarctic Krill; Aquarium; Maturity; Endogenous Rhythm; Overwintering Strategy
ABSTRACT
Antarctic krill is thought to undergo an annual cycle of biological processes to cope with seasonal changes in their environment. The question of whether, and to what degree, seasonal environmental parameters such as photoperiod, food availability and temperature govern metabolism in krill is not clear. In this long-term laboratory study, respiration rates were determined in krill incubated under simulated natural light cycle or total darkness, subjected to fed or starved conditions and on krill kept at different temperatures (−1˚C, 1˚C and 3˚C). There was a strong and significant increasing trend of respiration rates with month in all experimental treatments. In August (late winter), the mean respiration rates ranged between 0.22 - 0.35 μL O2 mg∙DW−1∙hr−1 for krill in both simulated seasonal light and complete darkness, and 0.25 - 0.26 μL O2 mg∙DW−1∙hr−1 for both fed and starved krill. Mean maximal respiration rates were recorded in October and December for all light and feeding treatments (0.46 - 0.56 μL O2 mg∙DW−1∙hr−1). Mean respiration rates for krill in the temperature treatments ranged between 0.24 - 0.30 μL O2 mg∙DW−1∙hr−1 in September reaching mean maximal rates in November and February (0.60 - 0.71 μL O2 mg∙DW−1∙hr−1). The covariate total length of krill was found to be non-significant and there was generally no significant interaction of experimental treatment with month and only for photoperiod comparison was the treatment main effect significant. The dark treatment gave higher respiration rates, and this needs careful interpretation. Results here suggest that, although light, food availability and temperature significantly affect metabolic rates, overall general seasonal pattern of winter low-summer high in respiration observed through the current experiment seemed to have followed an endogenous seasonal rhythm which had already been adjusted to the annual cycle prior to commencement of the experiment.
1. Introduction
Antarctic krill, Euphausia superba (hereafter krill), is a dominant pelagic species in the Southern Ocean and has significant economic and ecological importance [1-4]. Their overall prosperity and high biomass in the Southern Ocean is due to their ability to adapt to large seasonal changes in their environment; mainly sea-ice extent, food availability and light intensity and duration [5]. Currently, there are various theories on the overwintering strategies of krill [5]. The reduced primary productivity in the ocean, for up to eight months of the year, requires krill to adopt a range of strategies to survive and avoid periods of starvation [5-8]. Such strategies include overall body shrinkage and protein catabolism [9,10], utilization of lipid reserves [5,7,11-13], switching to a more omnivorous [14-16] and/or carnivorous [8,17,18] diet, as well as feeding on ice-algae [19,20] and sea-floor detritus [21], and suppression of metabolism [5,21,22].
Respiration rates of krill have been extensively studied in the past [5,18,21,23-32], but understanding the physiological responses of krill to a changing environment is scarce. A major question whether the observed decrease in metabolism in winter is caused simply by reduced food availability. Other important influential environmental factors may include photoperiod, temperature, or more complicated, an endogenous annual rhythm that is affected by a strong seasonal cue in the Southern Ocean.
Reduced metabolic rate and feeding activity are suggested to be the most effective energy-saving mechanisms for adult krill during the less productive months [5]. Nevertheless, Huntley et al. [17] suggests that starvation, body shrinkage and reduced metabolism are not common for krill in winter given they found winter krill feeding carnivorously and excreting at summer rates. This contrasts with Atkinson et al. [18] who demonstrated that respiration and clearance rates of juvenile and adult krill during autumn were only one-third of summer rates, which even failed to increase after 11 days of abundant food. Whereas, during the summer season of the same study, Atkinson and Snÿder [33] concluded that the metabolic and feeding activity of krill did respond positively to periods of high food concentrations. Irrespective of feeding conditions, Teschke et al. [32] revealed for the first time that the environmental light regime triggers the changes in metabolic rates of krill, suggesting the Antarctic light cycle is possibly the most important effect on the physiological parameters of krill. Hirano et al. [34] and Brown et al. [35] also suggested importance of light regime in governing maturation cycle of krill.
One of the main characteristics of the Southern Ocean is that water temperatures remain within a narrow range, with an annual variation rarely exceeding 5˚C [36]. An increase in temperature increases growth rate and metabolic activity in crustaceans [37]. Studies by McWhinnie and Marciniak [38], Rakusa-Suszczewski and Opalinski [23] and Segawa et al. [24] all conclude that respiration rate increased with increasing temperature for krill. However, to date, no study has examined the effects of this narrow temperature range on krill physiology within the context of other environmental factors varying over a long-term.
A series of recent work collectively suggest that an endogenous circannual timing mechanism is operating in krill controlling their physiology and behaviour, and that photoperiod is probably acting as the main Zeitgeber [39]. Information on respiration rates over a long period spanning a complete maturity cycle is therefore desired for a greater understanding of the drivers of seasonal patterns of krill metabolism.
In the present investigation, a long-term controlled experiment was conducted in the laboratory using krill that were incubated under different light, food and temperature regimes. The objectives were to test, and to determine, which of these key environmental factors dictate krill respiration rates between the critical period of maturation in late winter/early spring and sexual regression in late summer/early autumn.
2. Materials and Methods
2.1. Collection of Krill
Live Antarctic krill were collected on 7th February 2005 (66˚15'S, 74˚45'E) and 3rd March 2006 (66˚02'S, 79˚32'E) using a rectangular mid-water trawl net (RMT 8) [40] on board the RSV Aurora Australis. The water temperature during krill capture was −1˚C. Krill were transferred into 200 - L tanks and maintained with a continuous supply of seawater in a cold laboratory (0˚C, dim light and no food) on board the ship. Once north of the Polar Front, the continuous water supply was cut off and 50% of the tank water was exchanged with freshly pre-chilled seawater each day.
2.2. Aquaria Conditions
On return to the research aquarium at the Australian Antarctic Division in Kingston, Tasmania, krill were acclimated to the aquarium conditions [41]. Krill were maintained under light conditions that were adjusted throughout the experimental period to mimic the natural Antarctic seasonal light cycle. Lighting was provided by twin fluorescent tubes. A personal computer controlled-timer system was used to set a natural photoperiod corresponding to that for the Southern Ocean (66˚S at 30 m depth). Continuous light and a maximum of 100 lux light intensity at the surface of the tank (assuming 1% light penetration to 30 m depth) during summer midday (December), a sinusoidal annual cycle with monthly variations of photoperiod and daily variation of light intensity was calculated. At the start of each month, a new photoperiod was simulated by adjusting the timer system [42].
Krill were kept in eight different experimental conditions (natural light cycle or complete darkness, fed or starved conditions and temperatures ranging between −1˚C and 3˚C), as outlined in Table 1. Density of krill in each of the tanks was 0.5 - 2 inds/L. Overall krill mortality in the system is approximately 0.1% of the population per day. An algae mixture of the cultured pennate diatom Phaeodactylum tricornutum, and the flagellates Pavlova sp. and Isochrysis sp., and diatom Thalassiosira sp., which are concentrated bulk feeds of instant algae mixed with seawater (Reed Mariculture, California), and minced clam meat were fed at various regimes summarised in Table 1.
Due to operational reasons (large size of tanks and space constraints) experimental tanks were not replicated to account for “tank effects”.
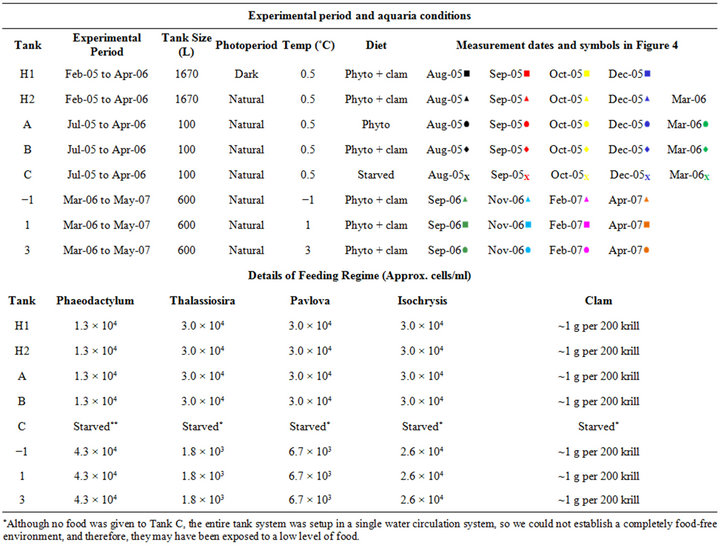
Table 1. Summary of the experimental conditions.
2.3. Measurement of Respiration Rates
Respiration measurements were conducted in filtered seawater (0.1µm pore size). From each of the experimental tanks, four random krill were selected and incubated individually in 1.2 - L glass reagent bottles filled with filtered seawater at the various time points. Krill were rinsed with filtered water that was acclimated to the correct temperature and added to the bottle. Two controls of the same volume without krill were also taken for each experimental treatment. The krill and control bottles were incubated for 24 hours. At the end of the incubation period, oxygen concentrations were measured after immediate fixing for Winkler titrations, as described in Omori and Ikeda [43] and Meyer et al. [44] by the use of a titrator; 716 DMS Titrino (METROHM). The total length (TL) of krill was measured (Standard Length 1) and the dry weight (DW) was calculated using the following empirical Equation:
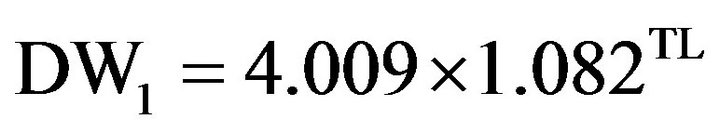
while an alternative Equation is
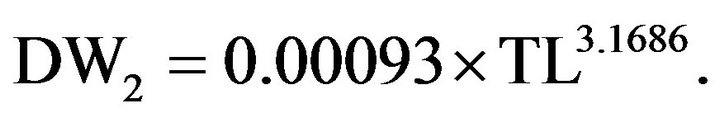 [45]
[45]
For the sample of TL in this study, the correlation between predicted DW for these two equations was 0.999 and the average ratio, DW2/DW1, was 1.16 with a range of 1.06 to 1.17.
2.4. Statistical Analysis
The data was analysed using the GenStat package [46] to provide an ANOVA for each dataset and for the corresponding treatment applied in each. Initially, the TL was also included as a covariate in each ANOVA. A trend analysis with month was also determined by using a decomposition of the month factor sums of squares into linear and quadratic orthogonal polynomial components with the remainder degrees of freedom used to obtain a lack-of-fit statistic. To do this, a continuous version of the month factor was created as a variable using integers 1 (August) through to 9 (April), even though for each treatment only some of these months were measured. For the temperature treatment a similar orthogonal polynomial decomposition was applied, but restricted to linear and lack-of-fit terms since there were only three temperature levels tested. Residual plots and histograms combined with normal quantile-quantile plots for each variable were used to examine whether or not the assumptions of homogeneous variance and normal distribution for the residuals were reasonable.
3. Results
Respiration rates were not significantly different (P > 0.05) between the randomly selected males and females at each of the measured time points so the data was combined for sex in all experimental conditions. There was a significant and strong increasing trend with month, which showed an increase in respiration rate from winter to summer in all experimental treatments (Figures 1-3). Mean respiration rates for krill on a DW basis in the treatments natural light versus complete darkness were 0.22 and 0.35 μL O2 mg∙DW−1∙hr−1, respectively, in August (Figure 1) and gradually increased to December (0.46 and 0.56 μL O2 mg∙DW−1∙hr−1, respectively). Mean respiration rates for krill in the treatments fed versus starved in August ranged between 0.25 - 0.26 μL O2 mg∙ DW−1∙hr−1 (Figure 2), showing a gradual increase into summer. Krill in Tank A (phytoplankton plus clam) reached mean maximal rates in December (0.51 μL O2
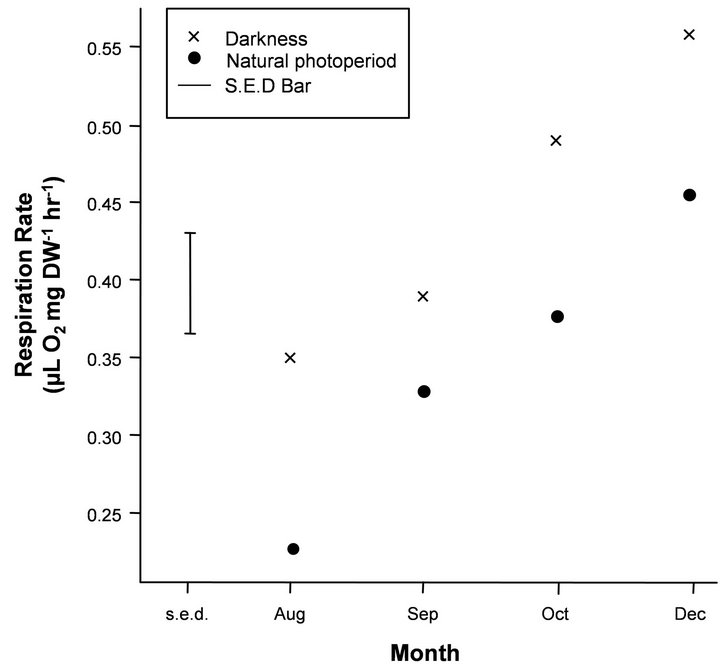
Figure 1. Mean respiration rates on a dry weight (DW) basis (μL O2 mg∙DW−1∙hr−1) for the photoperiod treatment (natural light versus complete darkness). The Standard Error of Difference bar (s.e.d) is based on the residual mean square (m.s.) given in Table 2 and the sample of four krill in each treatment by month combination and gives the standard error of the pairwise differences in means for a given month. Any differences that are more than twice the length of the s.e.d. bar based on a t-statistic of 2.0 can be considered significant at the 95% level.
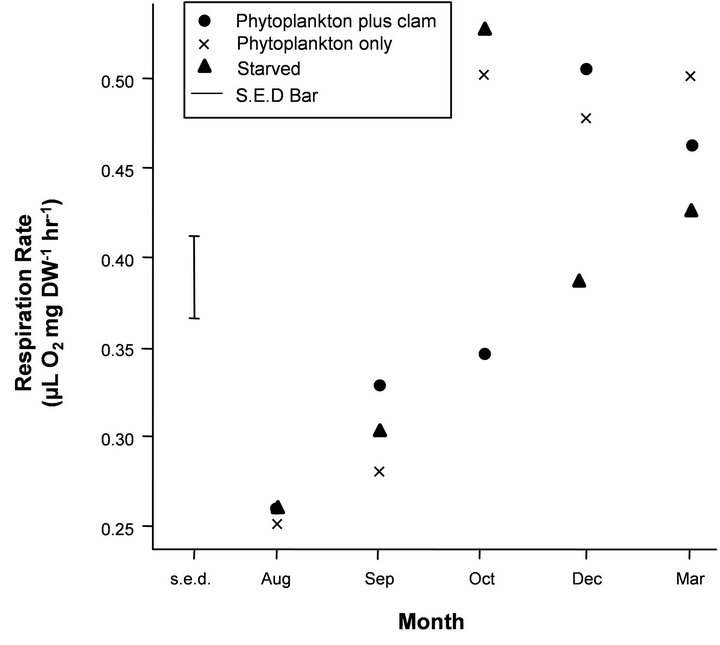
Figure 2. Mean respiration rates on a dry weight (DW) basis (μL O2 mg∙DW−1∙hr−1) for the feeding regime treatment (fed versus starved krill). The Standard Error of Difference bar (s.e.d) is based on the residual mean square (m.s.) given in Table 2 and the sample of four krill in each treatment by month combination and gives the standard error of the pairwise differences in means for a given month. Any differences that are more than twice the length of the s.e.d. bar based on a t-statistic of 2.0 can be considered significant at the 95% level.
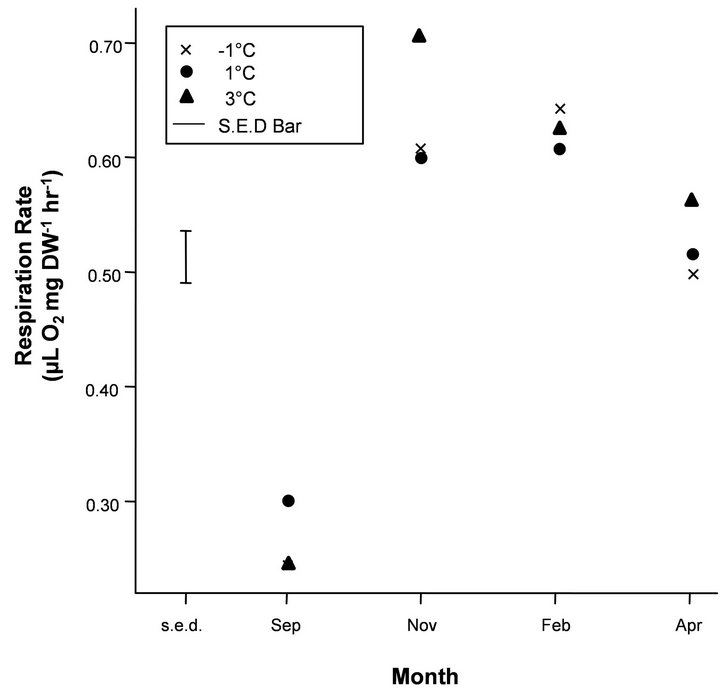
Figure 3. Mean respiration rates on a dry weight (DW) basis (μL O2 mg∙DW−1∙hr−1) for the temperature treatment (−1˚C, 1˚C and 3˚C). The Standard Error of Difference bar (s.e.d) is based on the residual mean square (m.s.) given in Table 2 and the sample of four krill in each treatment by month combination and gives the standard error of the pairwise differences in means for a given month. Any differences that are more than twice the length of the s.e.d. bar based on a t-statistic of 2.0 can be considered significant at the 95% level.
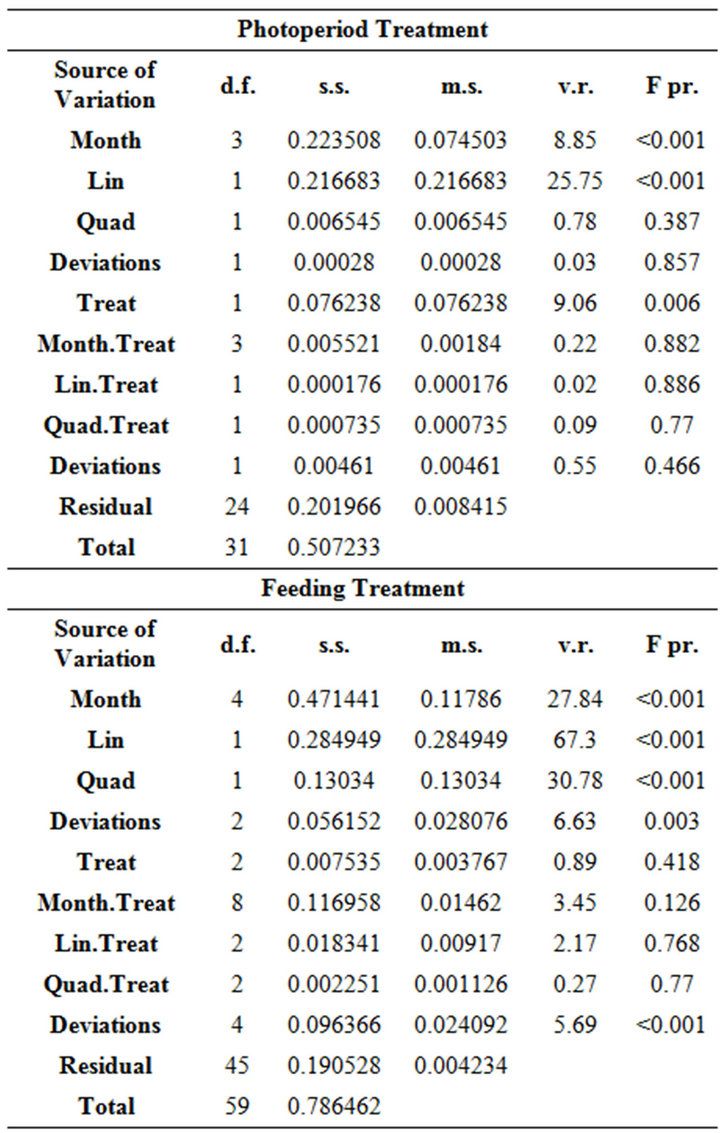
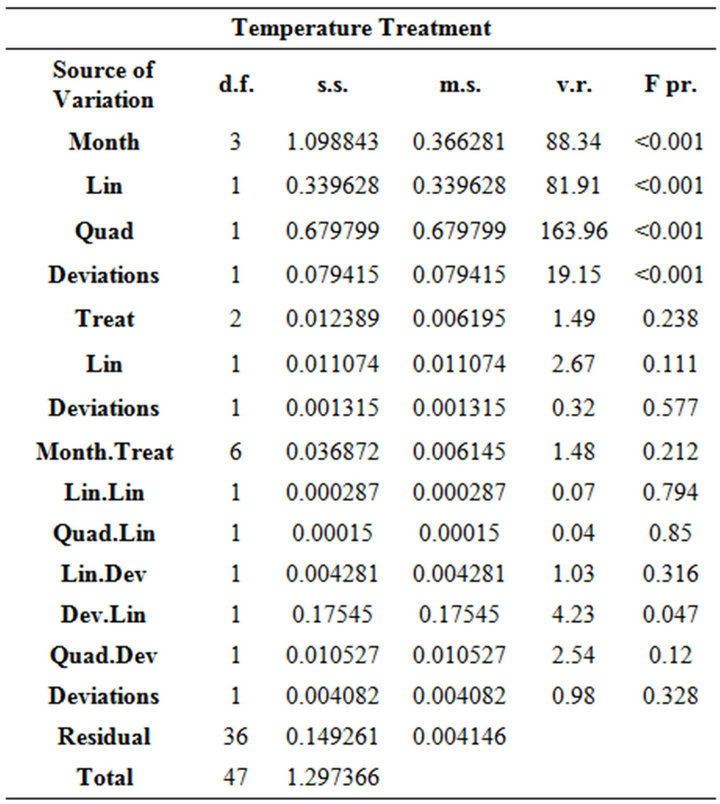
Table 2. ANOVA table for each of the experimental treatments with the linear and quadratic orthogonal polynomial decomposition. The covariate total length (TL) was nonsignificant and therefore not included in this table.
mg∙DW−1∙hr−1), with similar rates measured at the end of the experiment in March. Krill in Tank B (phytoplankton only) reached mean maximal respiration rates in October (0.51 μL O2 mg∙DW−1∙hr−1), and the mean respiration was stabilised at approximately these rates throughout summer until March. Krill in Tank C (starved), however, reached maximal respiration rates in October (0.53 μL O2 mg∙DW−1∙hr−1), which then decreased in December and March (0.39 and 0.43 μL O2 mg∙DW−1∙hr−1, respectively). Under natural photoperiod, mean respiration rates of krill under varying temperature regimes (−1˚C, 1˚C and 3˚C) increased from September (means ranging between 0.24 - 0.30 μL O2 mg∙DW−1∙hr−1) to reach mean maximal rates in November and February (0.60 - 0.71 μL O2 mg∙ DW−1∙hr−1), before a slight decrease in April (0.50 - 0.57 μL O2 mg∙DW−1∙hr−1), under constant food conditions (Figure 3).
The covariate TL was non-significant and there was no significant interaction of treatments with the above trend of month. Residual plots and histograms combined with normal quantile-quantile plots (graphs not shown) showed that the assumptions of homogeneous variance and normal distribution for the residuals were reasonable for each dataset. The mean respiration rates were lower in the natural light treatment compared to complete darkness, and the same trend with month was evident for both treatments (Figure 1). This was determined from the ANOVA since a significant interaction between treatments in this trend was not detected. However, the main effect of treatment was significant (P < 0.01) (Table 2), as expressed in the lower respiration rates for the natural light regime (Figure 1). The corresponding mean difference detected by the main effect term (i.e. averaged over months) was 0.097 with SE of 0.023.
For the food treatments, despite the general increasing trend in respiration rate with month for all treatments (significant “lin” and “quad” terms in Table 2, Figure 2), there was a somewhat strange result for the food treatments in October, which showed respiration rates for animals fed with phytoplankton and clam were significantly lower than those krill fed with phytoplankton only and the starved treatment (Figure 2). This caused the trend analysis (lin + quad) to have a significant lack-offit (“Deviations” terms in Table 2) for both the month main effects and month by treatment interaction.
There was a general increasing trend in respiration rate with month for all temperature treatments, but no significant interaction with treatment or main effect of treatment (Table 2). The significant lack-of-fit term (i.e. “Deviations”) for the month decomposition for the temperature treatment is due to the very sharp increase in respiration rate between September and November, which is difficult to model using a 2nd degree polynomial. The “Dev.Lin” term in the decomposition of the month by treatment interaction represents the difference in the linear trend with temperature of each month’s deviations from the fitted 2nd degree polynomial and is only just significant at the 5% probability level. This is due to the reversal of the order of treatments with respect to respiration rate for September for which the 1˚C regime gave the highest rate which was not the case for the other months. However, considering the length of the S.E.D bar in Figure 3 relative to the differences between temperatures for each month considered separately indicates a lack of statistical significance and combined with this, the marginally significant “Dev.Lin” term, makes any strong inferences drawn from the significance of this term unjustified.
Individual respiration rates, as a function of total length, were compared to a field based study by Quetin and Ross [5] (Figure 4). Laboratory data from this study was generally observed to fall between Quetin and Ross’ winter (July −1.5˚C—not feeding) and summer (January 2˚C—feeding) trend lines, which represented the relationship between total length and oxygen consumption from the wild in Bransfield Strait [5].
When the dry weight equation giving DW2 was applied in place of DW1 the differences in results to those reported in Table 2 were very minor and therefore did not have any effect on the interpretation or conclusions derived from the reported results.
4. Discussion
There was a strong trend of respiration rate with month, with respiration increasing from late winter/early spring to late summer/early autumn, during the important and energy consuming maturation and regression process of krill, under all experimental conditions; natural Antarctic light cycle versus complete darkness, fed versus starved krill and krill subjected to different temperature regimes (−1˚C, 1˚C and 3˚C). The covariate total length of krill was found to be non-significant and there was no significant interaction of experimental treatment with month or main effect of each treatment. However, the power to detect a difference was low, due to the low sample number of four krill in each month and treatment. Due to experimental design it was not possible to examine any interaction among the three possible environmental parameters. Nevertheless, although the power to detect a difference was low the results from this study suggest that the three experimental parameters (light, food and temperature) were not the primary factors directly driving the overall seasonal pattern of respiration rates that have been observed through the experiment between September and April.
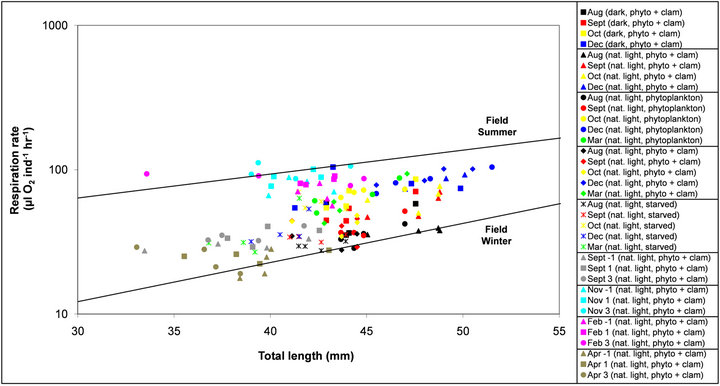
Figure 4. Laboratory and field measurements of respiration rates (µL O2 ind−1∙hr−1, on a logarithmic scale) of Antarctic krill as a function of total length (mm). The two trend lines are an approximate representation of Quetin and Ross (1991) field respiration measurements in the summer (January 2˚C) and winter (July −1.5˚C) at Bransfield Strait for krill between 30 and 55mm in length. Laboratory measurements are represented by the coloured symbols at various time points (combined sex) and experimental conditions (natural light versus complete darkness, fed versus starved krill and temperatures between −1˚C and 3˚C). Refer to Table 1 for details on experimental conditions.
Below we discuss the potential candidate mechanisms that could be related to seasonal changes in metabolism.
4.1. Food Availability
Recruitment and population size of krill are strongly influenced and dictated by the survival success during the dark and less productive winter [5,18,47,48] Krill undertake a variety of strategies to avoid starvation during the season of reduced primary productivity. Krill are known to exploit many types of food resources in the wild so the chances they will actually encounter overall food shortages may be small [49]. Krill almost certainly exploit whatever particulate matter is in the water column including elements of the microbial loop and marine snow [50], as well as ice-algae [19,20] and sea-floor detritus [21,51]. Heterotrophic feeding on protozoans [52] and copepods [8,17,18] are also reported. Lowering overall feeding activity and metabolism are also suggested as overwintering strategies [5,18, 21,22,39,53]. It is difficult to recognize and interpret whether metabolic rates are the result of changes in feeding activity, or vice versa, feeding activity is reflected by the changes in metabolic rates [32]. According to Ikeda and Dixon [29], an increase in metabolic activity in krill was reported to follow the ingestion of food.
Quetin and Ross [5] showed that oxygen consumption in a large size range of adult krill from Bransfield Strait was 33% higher in a summer experimental setup (January, 2˚C) compared to winter (July, −1.5˚C). They suggested that adult krill do not feed in winter, and have considerably lowered metabolic rates and negative or zero growth rates. Meyer et al. [53], through examining size and colour of digestive gland, suggested that krill had either not been feeding or had ingested a heterotrophic and autotrophic diet at low rates during winter. Huntley et al. [17] examined krill west of the Antarctic Peninsula during winter (July-August) and the preceding summer (December-January) in a region similar to that of Quetin and Ross [5]. Huntley et al. [17] demonstrated that krill fed on a carnivorous diet during winter, particularly copepods, which enabled them to sustain growth comparable to that of summer. They further suggested that starvation, body shrinkage and reduced metabolism is not common for krill during winter, but instead, carnivory is sufficient for growth and to meet energy requirements until phytoplankton blooms occur in spring.
Under constant food supply in this study, krill were observed to be feeding year round. The digestive glands were dark green and healthy looking during the experiment (personal observations), indicating that krill were continuously feeding [54-56]. Total lipid and fatty acid content and composition analysis of the digestive gland and the whole body of krill (Brown et al. unpublished)
also showed that krill were actively feeding year round.
Our results may help interpret respiration rates reported in winter krill by Quetin and Ross [5]. Despite constant year round feeding, a reduction in respiration rates was observed in the current study in all experimenttal krill during the non-summer months on a dry weight basis. Respiration rates also increased in krill with month in both fed and starved krill.
At an individual level, the recorded respiration from krill in the laboratory was distributed fully within the whole range of those measured from summer (feeding) and winter (non-feeding) field experiments by Quetin and Ross [5], as shown in Figure 4. This demonstrated the fact that respiration rates of well fed krill under simulated winter conditions were similar to those nonfeeding winter observations from the field. These krill in Bransfield Strait, despite the fact there may be limited sea-ice, may in fact be feeding to some extent and not starving as it has been suggested [5]. Their conclusions came about from observing that both the faecal pellet production and ingestion of phytoplankton in winter were less than 3% of summer rates, suggesting that adult krill did not derive a large proportion of their energy from carnivory. From our observations, we conclude that when food density is low, krill tend to retain their gut content and do not excrete at the same rate as when they are fully fed, which could have also been the case in wild krill from Quetin and Ross [5].
Reasons for why rates under starvation are maximal in October but not at anytime where values are minimal or medium (Figure 2) remains unsolved. This somewhat contradicts our current general understanding. However, October is the month when krill is becoming meta-bolically active due to its rapid maturation and ovary development therefore maturation process could be the reason behind, and warrants further detailed study in order to confirm this deviation from the general trend in October.
4.2. Temperature
Krill is considered to be stenothermal and is sensitive to slight changes in temperature and other environmental changes [57]. Temperature has been shown to influence the frequency of moulting in krill, and thus overall growth rates, which can vary considerably, even within a narrow annual temperature range that is observed in the Southern Ocean [23,24,38,58,59,60]. McWhinnie and Marciniak [38], Rakusa-Suszczewski and Opalinski [23] and Segawa et al. [24] have also suggested that temperature is a key environmental factor influencing the metabolism of krill, with increasing respiration rates with increasing temperature (0˚C to 5˚C, −1˚C to 2.4˚C, and −1.5˚C to 5˚C, respectively). This is generally true up to 5˚C, but above this temperature, krill are probably thermally stressed [59,61]. This relationship between temperature and respiration has been reported for another euphausiid species. Respiration rates in Thysanoessa longipes in the Japan Sea increased exponentially with an increase of temperature from 0˚C to 8˚C [62].
In our study, however, there was no significant interacttion with the temperature treatments (−1˚C to 3˚C) and month or main effect of temperature, indicating that temperature was not a key environmental variable to cue changes in seasonal metabolic activity.
4.3. Photoperiod
Light intensity and duration has strong seasonality in the Southern Ocean, from near-constant light in December to near-constant darkness in June. Photoperiod has been suggested as the likely trigger and possibly has the most important effect on the changes in metabolic activity of krill [32]. The respiration of krill in the starved and fed treatments, as well as krill in the various temperature regimes in our study, increased with month during the stages of maturation and sexual regression under a natural light period. These results may suggest that photoperiod also has a significant influence on krill metabolic rates. However, the respiration rates of krill were higher in the complete darkness compared to a natural light treatment, with the same increasing trend with month. There was no significant interaction between treatments, but when all data was pooled to increase the power of the test, the main effect of treatment was significantly different (P < 0.01). These results oppose those by Teschke et al. [32] who reported a significant decrease in respiretion rates in krill incubated under complete darkness compared to light. Experimental conditions were different between these studies, with a tank size of 100 - L used by Teschke et al. [32], and 1670 - L in the present study. A larger tank may have facilitated an increase in swimming activity inducing higher respiration rates in this study. The fact that maximum respiration rates were recorded under darkness in our study could correspond to a higher swimming activity in darkness, and the larger size tank in our experiment compared to Teschke et al. [32] allowing active swimming leading to higher respiretion in the darkness. Additionally, the abrupt changes in the light regime, and also variation in timing and duration of the experiments in Teschke et al. [32], may account for the differences observed. Our study was a closer emulation of “natural” conditions in the wild compared to those of Teschke et al. [32] which may have influences these results.
4.4. Endogenous Annual Rhythm
Endogenous circadian timing system in Antarctic krill and its likely link to metabolic key processes has recently been reported and these may not only be critical for synchronization to the solar day but also for the control of seasonal events [63]. There are increasing number of studies suggesting adult krill entering a state of winter quiescence in the wild [18,53]. There is evidence on the one hand of continuous feeding throughout the year [17] and, on the other, of reduced respiration and lack of response to food increase [18]. The different conditions experienced by sampled populations undoubtedly underlie these contrasting observations. Published winter based respiration measurements from the field [5,17,21] and long-term controlled experiments in the laboratory (this study), were conducted during the late winter/early spring months so the seasonal timings of krill were in a winter state and comparable. Therefore, reductions in respiration levels were irrespective of effects of photoperiod and feeding conditions within the length of experimental period in this study. The general increasing respiration rates observed from winter to summer in this study, even in starved krill and those kept in complete darkness, were possibly caused by krill changing endogenously to a summer state. This idea of a seasonal endogenous pattern is also evident from the growth and maturation results in Brown et al. [59]. A clear seasonal cycle of growth and maturation for both males and females in the three temperature treatments (−1˚C, 1˚C and 3˚C) were observed under constant and high food concentrations. This is also in agreement with Thomas and Ikeda [64] who showed female krill underwent a regression of external sexual characteristics after spawning, which was accompanied by negative growth and an increase in intermoult period, in both starved and fed krill.
Many biological cycles operate through a combination of endogenous cycles and external environmental cues. If devoid of any external stimuli, free-running endogenous cycles eventually become unsynchronised from the ideal cycle since the mechanism that provide the periodicity are often imperfect [65]. A coordinating cue, or zeitgeber, is required to resynchronise the endogenous cycle. For example, it is believed that Northern krill (Meganyctiphanes norvegica) receive a zeitgeber from the dawn sun immediately prior to their descent to depths that are devoid of any further light cues [66]. Antarctic krill reproduction has successfully been initiated by providing continuous light after prolonged darkness in the aquarium [34]. Further, receiving a cue of darkness or accelerated shortening of day light at the timing of their maturity regression period has been demonstrated to play important role in adjusting their seasonal maturation cycle to the environment [35]. Although the results from our study indicate that the three experimental parameters (light, food and temperature) were not directly involved in dictating general overall pattern of respiration rate of krill, the general seasonal pattern in respiration rate could be result of a free running endogenous annual cycle. This pattern could have been synchronised to the seasonal light regime in the aquarium [42] prior to start of the experiment, but could not be changed due to constant darkness or constant food supply during the experiment [64,67].
5. Conclusion
Krill use a variety of strategies to survive the less productive winter, which highlights the fact that krill is a versatile species and is a contributing explanation for its high biomass in the Southern Ocean [68]. It has been widely accepted that a reduction in metabolic activity can occur during winter, with varying environmental variables considered to be responsible, which can differ between different regions of the Southern Ocean. Under controlled conditions, this long-term study has shown that the environmental variables; light, food availability and temperature, may not necessarily be the major factors in governing the metabolic cycles as general pattern of winter low summer high was observed in all treatments in this study. It is suspected that an endogenous rhythm was operating in krill [64,67], controlling overall general seasonal pattern of metabolic activity [39]. The significance of an “internal clock” mechanism is still relatively unknown and warrants further investigation, particularly examining photoperiod as the possible cue for dictating internal processes [35,39,63,64,67]. Future experiments will need to incorporate measurements over a whole cycle (over one year) with some overlaps to determine when an observed respiration cycle starts to become unsynchronised from the ideal annual pattern.
6. Acknowledgements
We are grateful to the captain and crew of RSV Aurora Australis for their skilful assistance during the collection of krill. We also extend our thanks to R. King and P. Cramp for their general support, and contribution and maintenance of the aquarium facilities at the Australian Antarctic Division. We also thank an anonymous reviewer for constructive comments. This work was supported by the Tasmanian Graduate Research Scholarship and the Australian government’s Cooperative Research Centres Programme through the Antarctic Climate and Ecosystems Cooperative Research Centre (ACE CRC).
REFERENCES
- I. Everson, “Krill Biology, Ecology and Fisheries,” Blackwell Science, Oxford, 2000.
- D. G. M. Miller and I. Hampton, “Biology and Ecology of the Antarctic Krill (Euphausia superba Dana): A Review,” BIOMASS Scientific Series, Vol. 9, 1989, pp. 1-166.
- M. Mangel and S. Nicol, “Krill and the Unity of Biology,” Canadian Journal of Fisheries and Aquatic Science, Vol. 57, Suppl. 3, 2000, pp. 1-5.
- S. H. Alonzo, P. V. Switzer and M. Mangel, “An Ecosystem-Based Approach to Management: Using Individual Behavior to Predict the Indirect Effects of Antarctic Krill Fisheries on Penguin Foraging,” Journal of Applied Ecology, Vol. 40, No. 4, 2003, pp. 692-702. doi:10.1046/j.1365-2664.2003.00830.x
- L. B. Quetin and R. M. Ross, “Behavioral and Physiological Characteristics of the Antarctic Krill, Euphausia superba,” American Zoologist, Vol. 31, No. 1, 1991, pp. 49-63.
- P. Virtue, S. Nicol and P. D. Nichols, “Changes in the Digestive Gland of Euphausia superba during ShortTerm Starvation: Lipid Class, Fatty Acid and Sterol Content and Composition,” Marine Biology, Vol. 117, No. 3, 1993, pp. 441-448.
- W. Hagen, G. Kattner, A. Terbruggen and E. S. Van Vleet, “Lipid Metabolism of the Antarctic krill Euphausia superba and Its Ecological Implications,” Marine Biology, Vol. 139, No. 1, 2001, pp. 95-104. doi:10.1007/s002270000527
- W. Hagen, T. Yoshida, P. Virtue, S. Kawaguchi, K. M. Swadling, S. Nicol and P. D. Nichols, “Effect of a Carnivorous Diet on the Lipids, Fatty Acids and Condition of Antarctic Krill, Euphausia superba,” Antarctic Science, Vo. 19, 2007, 183-188. doi:10.1017/S0954102007000259
- T. Ikeda, and P. Dixon, “Body Shrinkage as a Possible Overwintering Mechanism of the Antarctic Krill (Euphausia superba Dana),” Journal of Experimental Marine Biology and Ecology, Vol. 62, No. 2, 1982, pp. 143-151. doi:10.1016/0022-0981(82)90088-0
- S. Nicol, M. Stolp, T. G. Cochran, P. Geijsel and J. Marshall, “Growth and Shrinkage of Antarctic krill Euphausia superba from the Indian Ocean Sector of the Southern Ocean during Summer,” Marine Ecology Progress Series, Vol. 89, 1992, pp. 175-181. doi:10.3354/meps089175
- A. Clarke, “Lipid content and composition of Antarctic krill, Euphausia superba Dana,” Journal of Crustacean Biology, Vol. 4, No. 1, 1984, pp. 285-294.
- W. Hagen, E. S. Van Vleet and G. Kattner, “Seasonal Lipid Storage as Overwintering Strategy of Antarctic Krill,” Marine Ecology Progress Series, Vol. 134, 1996, pp. 85-89. doi:10.3354/meps134085
- P. Virtue, P. D. Nichols, S. Nicol and G. W. Hosie, “Reproductive Trade-off in Male Antarctic Krill, Euphausia superb,” Marine Biology, Vol. 126, No. 3, 1996, pp. 521- 527. doi:10.1007/BF00354634
- A. Clarke, “The Biochemical Composition of Krill, Euphausia superba Dana from South Georgia,” Journal of Experimental Marine Biology and Ecology, Vol. 43, No. 3, 1980, pp. 221-236. doi:10.1016/0022-0981(80)90049-0
- J. Mauchline, “The Biology of Mysids and Euphausiids (Advanced Marine Biology, 18),” Academic Press, Waltham, 1980, pp. 1-677.
- H. J. Price, K. R. Boyd and C. M. Boyd, “Omnivorous Feeding Behavior of the Antarctic Krill Euphausia superb,” Marine Biology. Vol. 97, No. 1, 1988, pp. 67-77. doi:10.1007/BF00391246
- M. E. Huntley, W. Nordhausen and M. D. G. Lopez, “Elemental Composition, Metabolic Activity and Growth of Antarctic Krill Euphausia superba during Winter,” Marine Ecology Progress Series, Vol. 107, 1994, pp. 23- 40. doi:10.3354/meps107023
- A. Atkinson, B. Meyer, D. Stübing, W. Hagen, K. Schmidt and U. V. Bathmann, “Feeding and Energy Budgets of Antarctic Krill Euphausia superba at the Onset of Winter II. Juveniles and Adults,” Limnology and Oceanography, Vol. 47, No. 4, 2002, 953-966. doi:10.4319/lo.2002.47.4.0953
- H. P. Marschall, “The Overwintering Strategy of Antarctic Krill under the Pack Ice of the Weddell Sea,” Polar Biology, Vol. 9, No. 2, 1988, pp. 129-135. doi:10.1007/BF00442041
- J. J. Stretch, P. P. Hamner, W. M. Hamner, W. C. Michel, J. Cook and C. W. Sullivan, “Foraging Behavior of Antarctic Krill Euphausia superba on Sea Ice Microalgae,” Marine Ecology Progress Series, Vo. 44, No. 2, 1988, pp. 131-139. doi:10.3354/meps044131
- K. Kawaguchi, S. Ishikawa and T. Matsuda, “The Overwintering Strategy of Antarctic Krill (Euphausia superba Dana) under the Coastal Fast Ice off the Ongul Islands in Lützol-Holm Bay, Antarctica,” Memoirs of the National Institute of Polar Research, Vol. 44, 1986, pp. 67-85.
- J. J. Torres, J. Donnelly, T. L. Hopkins, T. M. Lancraft, A. V. Aarset and D. G. Ainley, “Proximate Composition and Overwintering Strategies of Antarctic Micronektonic Crustacea,” Marine Ecology Progress Series, Vol. 113, No. 3, 1994, pp. 221-232. doi:10.3354/meps113221
- S. Rakusa-Suszczewski and K. W. Opalinski, “Oxygen Consumption in Euphausia superba,” Polskie Archiwum Hydrobiologii, Vol. 25, No. 3, 1978, pp. 633-641.
- S. Segawa, M. Kato and M. Murano, “Oxygen Consumption of the Antarctic Krill,” Transactions of the Tokyo University of Fisheries, Vol. 3, 1979, pp. 113-119.
- S. Segawa, M. Kato and M. Murano, “Respiration and Ammonia Excretion Rates of the Antarctic Krill, Euphausia superba Dana,” Transactions of the Tokyo University of Fisheries, Vol. 5, 1982, pp. 177-187.
- T. Ikeda and A. W. Mitchell, “Oxygen Uptake, Ammonia Excretion and Phosphate Excretion by Krill and Other Antarctic Zooplankton in Relation to Their Body Size and Chemical Composition,” Marine Biology, Vol. 71, No. 3, 1982, pp. 283-298. doi:10.1007/BF00397045
- H. J. Hirche, “Excretion and Respiration of the Antarctic Krill Euphausia superba,” Polar Biology, Vol. 1, No. 4, 1983, pp. 205-209. doi:10.1007/BF00443189
- T. Ikeda, “Sequence in Metabolic Rates and Elemental Composition (C, N, P) during the Developmental of Euphausia superba Dana and Estimated Food Requirements during Its Life Span,” Journal of Crustacean Biology, Vol. 4, No. 1, 1984, pp. 273-284.
- T. Ikeda and P. Dixon, “The Influence of Feeding on the Metabolic Activity of Antarctic Krill (Euphausia superba Dana),” Polar Biology, Vol. 3, No. 1, 1984, pp. 1-9. doi:10.1007/BF00265561
- K. W. Opalinski, “Respiratory metabolism and metabolic adaptations of Antarctic krill Euphausia superb,” Polskie Archiwum Hydrobiologii, Vol. 38, No. 2, 1991, pp. 183- 263.
- K. M. Swadling, D. A. Ritz, S. Nicol, J. E. Osborn and L. J. Gurney, “Respiration Rate and Cost of Swimming for Antarctic Krill, Euphausia superba, in Large Groups in the Laboratory,” Marine Biology, Vol. 146, No. 6, 2005, pp. 1169-1175. doi:10.1007/s00227-004-1519-z
- M. Teschke, S. Kawaguchi and B. Meyer, “Simulated Light Regimes Affect Feeding and Metabolism of Antarctic Krill, Euphausia superb,” Limnology and Oceanography, Vo. 52, No. 3, 2007, pp. 1046-1054. doi:10.4319/lo.2007.52.3.1046
- A. Atkinson and R. Snÿder, “Krill-Copepod Interactions at South Georgia Antarctica, I. Omnivory by Euphausia superb,” Marine Ecology Progress Series, Vol. 160, 1997, pp. 63-76. doi:10.3354/meps160063
- Y. Hirano, T. Matsuda and S. Kawaguchi, “Breeding Krill in Captivity,” Marine and Freshwater Behavior and Physiology, Vol. 36, No. 4, 2003, pp. 249-258. doi:10.1080/10236240310001623637
- M. Brown, S. Kawaguchi, R. King, P. Virtue and S. Nicol, “Flexible Adaptation of the Seasonal Krill Maturity Cycle in the Laboratory,” Journal of Plankton Research, Vol. 33, No. 5, 2011, pp. 821-826. doi:10.1093/plankt/fbq123
- G. A. Knox, “Biology of the Southern Ocean,” Cambridge University Press, Cambridge, 1994.
- R. G. Hartnoll, “Growth in Crustacea—Twenty Years on,” Hydrobiologia, Vol. 449, No. 1-3, 2001, pp. 111-122. doi:10.1023/A:1017597104367
- M. A. McWhinnie and P. Marciniak, “Temperature Responses and Tissue Respiration in Antarctic Crustacea with Particular Reference to the Krill Euphausia superba,” Antarctic Research Series, Vol. 1, 1964, pp. 63-72. doi:10.1029/AR001p0063
- B. Meyer, “The Overwintering of Antarctic Krill, Euphausia superba, from an Ecophysiological Perspective,” Polar Biology, Vol. 35, No. 1, 2012, pp. 15-37. doi:10.1007/s00300-011-1120-0
- A. de. C. Baker, M. R. Clarke and M. J. Harris, “The N.I.O. Combination Net (RMT) 1 + 8 and Further Developments of Rectangular Midwater Trawls,” Journal of Marine Biological Association of the United Kingdom, Vol. 53, No. 1, 1973, pp. 167-184. doi:10.1017/S0025315400056708
- R. King, S. Nicol, P. Cramp and K. M. Swadling, “Krill Maintenance and Experimentation at the Australian Antarctic Division,” Marine and Freshwater Behaviour and Physiology, Vol. 36, No. 4, 2003, pp. 271-283. doi:10.1080/10236240310001614457
- S. Kawaguchi, R. King, R. Meijers, J. E. Osborn, K. M. Swadling, D. A. Ritz and S. Nicol, “An Experimental Aquarium for Observing the Schooling Behaviour of Antarctic Krill,” Deep Sea Research Part II: Topical Studies in Oceanography, Vol. 57, No. 7-8, 2010, pp. 683-692. doi:10.1016/j.dsr2.2009.10.017
- M. Omori and T. Ikeda, “Methods in Marine Zooplankton Ecology,” John Wiley and Sons, New York, 1984.
- B. Meyer, A. Atkinson, D. Stübing, B. Oettl, W. Hagen and U. V. Bathmann, “Feeding and Energy Budgets of Antarctic krill Euphausia superba at the Onset of Winter—I. Furcilia III Larvae,” Limnology and Oceanography, Vo. 47, No. 4, 2002, pp. 943-952. doi:10.4319/lo.2002.47.4.0943
- D. J. Morris , J. L. Watkins, C. Ricketts, F. Buchholz and J. Priddle, “An Assessment of the Merits of Length and Weight Measurements of Antarctic Krill Euphausia superba,” British Antarctic Survey Bulletin, Vol. 79, 1988, pp. 27-50.
- Lawes Agricultural Trust, “GenStat Release 10.2 (PC/ Windows) (Rothamsted, UK),” VSN International Ltd., Hemel Hempstead, 2007.
- V. Siegel and V. Loeb, “Recruitment of Antarctic Krill Euphausia superba and Possible Causes for Its Variability,” Marine Ecology Progress Series, Vol. 123, 1995, pp. 45-56. doi:10.3354/meps123045
- V. Siegel, “Krill (Euphausiacea) Demography and Variability in Abundance and Distribution,” Canadian Journal of Fisheries and Aquatic Sciences, Vol. 57, Suppl. 3, 2000, pp. 151-167.
- V. Siegel and S. Nicol, “Population Parameters,” In: I. Everson, Ed., Krill: Biology, Ecology and Fisheries. Blackwell Science, London, 2000, pp. 102-149.
- A. Clarke and P. A. Tyler, “Adult Antarctic Krill Feeding at Abyssal Depths,” Current Biology, Vol. 18, No. 4, 2008, pp. 282-285. doi:10.1016/j.cub.2008.01.059
- K. Schmidt, A. Atkinson, S. Steigenberger, S. Fielding, M. C. M. Lindsay, D. W. Pond, G. A. Tarling, T. A. Klevjer, C. S. Allen, S. Nicol and E. P. Achterberg, “Seabed Foraging by Antarctic Krill: Implications for Stock Assessment, Bentho-Pelagic Coupling, and the Vertical Transfer of Iron,” Limnology and Oceanography, Vol. 56, No. 4, 2012, pp. 1411-1428. doi:10.4319/lo.2011.56.4.1411
- K. Schmidt, A. Akinson, K. -J. Petzke, M. Voss and D. W. Pond, “Protozoans as a Food Source for Antarctic Krill, Euphausia superba: Complementary Insights from Stomach Content, Fatty Acids, and Stable Isotopes,” Limnology and Oceanography, Vol. 51, No. 5, 2006, pp. 2409- 2427. doi:10.4319/lo.2006.51.5.2409
- B. Meyer, L. Auerswald, V. Siegel, S. Spahic, C. Pape, B. A. Fach, M. Teschke, A. L. Lopata and V. Fuentes, “Seasonal Variation in Body Composition, Metabolic Activity, Feeding, and Growth of Adult Krill Euphausia superba in the Lazarev Sea,” Marine Ecology Progress Series, Vol. 398, 2010, pp. 1-18. doi:10.3354/meps08371
- S. Kawaguchi, T. Ichii and M. Naganobu, “Green Krill, the Indicator of Microand Nano-Size Phytoplankton Availability to Krill,” Polar Biology, Vol. 22, No. 2, 1999, pp. 133-136. doi:10.1007/s003000050400
- S. Nicol, P. Virtue, R. King, S. R. Davenport, A. F. McGaffin and P. D. Nichols, “Condition of Euphausia crystallorophias off East Antarctica in Winter in Comparison to Other Seasons,” Deep Sea Research Part II: Topical Studies in Oceanography, Vol. 51, No. 17-19, 2004, pp. 2215-2224. doi:10.1016/j.dsr2.2004.07.002
- C. O’Brien, P. Virtue, S. Kawaguchi and P. D. Nichols, “Aspects of Krill Growth and Condition during Late Winter-Early Spring off East Antarctica (110-130˚E),” Deep Sea Research Part II: Topical Studies in Oceanography, Vol. 58, No. 9-10, 2011, pp. 1211-1221. doi:10.1016/j.dsr2.2010.11.001
- J. Wiedenmann, K. Cresswell and M. Mangel, “Temperature-Dependent Growth of Antarctic Krill: Predictions for a Changing Climate from a Cohort Model,” Marine Ecology Progress Series, Vol. 358, 2008, pp. 191-202. doi:10.3354/meps07350
- S. Nicol, “Understanding Krill Growth and Ageing: The Contribution of Experimental Studies,” Canadian Journal of Fisheries and Aquatic Science, Vol. 57, Suppl. 3, 2000, pp. 168-177.
- M. Brown, S. Kawaguchi, S. Candy and P. Virtue, “Temperature Effects on the Growth and Maturation of Antarctic Krill (Euphausia superba),” Deep Sea Research Part II: Topical Studies in Oceanography, Vol. 57, No. 7-8, 2010, pp. 672-682. doi:10.1016/j.dsr2.2009.10.016
- [61] A. Atkinson, R. S. Shreeve, A. G. Hirst, P. Rothery, G. A. Tarling, D. Pond, R. E. Korb, E. J. Murphy and J. L. Watkins, “Natural Growth Rates in Antarctic Krill (Euphausia superba): II. Predictive Models Based on Food, Temperature, Body Length, Sex, and Maturity Stage,” Limnology and Oceanography, Vol. 51, No. 2, 2006, pp. 973-987. doi:10.4319/lo.2006.51.2.0973
- [62] P. Mayzaud, E. Albessard, P. Virtue and M. Boutoute, “Environmental Constraints on the Lipid Composition and Metabolism of Euphausiids: The Case of Euphausia superba and Meganyctiphanes norvegica,” Canadian Journal of Fisheries and Aquatic Science, Vol. 57, Suppl. 3, 2000, pp. 91-103.
- [63] N. Iguchi and T. Ikeda, “Effects of Temperature on Metabolism, Growth and Growth Efficiency of Thysanoessa longipes (Crustacea: Euphausiacea) in the Japan Sea,” Journal of Plankton Research, Vol. 27, No. 1, 2005, pp. 1-10. doi:10.1093/plankt/fbh146
- [64] M. Teschke, S. Wendt, S. Kawaguchi, A. Kramer and B. Meyer, “A Circadian Clock in Antarctic Krill: An Endogenous Timing System Governs Metabolic Output Rhythms in the Euphausid Species Euphausia superb,” PLoS ONE, Vol. 6, No. 10, 2011, Article ID: e26090. doi:10.1371/journal.pone.0026090
- [65] P. G. Thomas and T. Ikeda, “Sexual Regression, Shrinkage, Re-Maturation and Growth of Spent Female Euphausia superba in the Laboratory,” Marine Biology, Vol. 95, No. 3, 1987, pp. 357-363. doi:10.1007/BF00409565
- [66] E. Gaten, G. Tarling, H. Dowse, C. Kyriacou and E. Rosata, “Is Vertical Migration in Antarctic Krill (Euphausia superba) Influenced by an Underlying Circadian Rhythm?” Journal of Genetics, Vol. 87, No. 5, 2008, pp. 473- 483. doi:10.1007/s12041-008-0070-y
- [67] G. A. Tarling, J. Cuzin-Roudy and F. Buchholz, “Vertical Migration Behaviour in the Northern Krill Meganyctiphanes norvegica Is Influenced by Moult and Reproductive Processes,” Marine Ecology Progress Series, Vol. 190, 1999, pp. 253-262. doi:10.3354/meps190253
- [68] S. Kawaguchi, T. Yoshida, L. Finley, P. Cramp and S. Nicol, “The Krill Maturity Cycle: A Conceptual Model of the Seasonal Cycle in Antarctic Krill,” Polar Biology, Vol. 30, No. 6, 2007, pp. 689-698. doi:10.1007/s00300-006-0226-2
- [69] W. Hagen, “Reproductive Strategies and Energetic Adaptations of Polar Zooplankton,” Invertebrate Reproduction and Development, Vol. 36, No. 1-3, 1999, pp. 25-34. doi:10.1080/07924259.1999.9652674
NOTES
*Corresponding author.