Open Journal of Ecology
Vol.4 No.4(2014), Article ID:44352,20 pages DOI:10.4236/oje.2014.44021
Long Term Effect of Major Disturbances on the Northern Mixed Grassland Ecosystem—A Review
Meng Li*, Xulin Guo
Department of Geography, University of Saskatchewan, Saskatoon, Canada
Email: *gismatthew@gmail.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received 24 January 2014; revised 24 February 2014; accepted 2 March 2014
ABSTRACT
Grassland ecosystems in North America have been significantly altered through various disturbances from past to present. This paper has identified and examined major disturbances in the grassland ecosystems, which include grazing, fire and drought. A brief history of each disturbance is reviewed and its impact on the grassland ecosystem is discussed, by synthesizing previous research efforts available in the current literature. The paper intends to understand the grassland ecosystem from a comprehensive perspective and particularly tries to articulate the compound effect of the major disturbances in the same context instead of viewing them in isolations. It is concluded that a holistic understanding of the interaction between disturbances and plant communities is essential for sustainable grassland management, which demands comprehensive research effort in this area. The paper also points out the challenges we are facing in the current researches and suggests potential future improvement.
Keywords
Grassland Ecosystem; Disturbances; Grazing; Fire; Drought

1. Introduction
Grasslands are ubiquitous throughout all continents. As one of major vegetation types they cover approximately 40% of land surface [1] [2] , and are especially significant in the study of terrestrial ecosystems. Grasslands in North America cover about 15% of its continent [1] , and commonly referred to as prairies, stretching from central Canada to southern U.S. on the Great Plains (Figure 1). The Great Plains have different major grassland
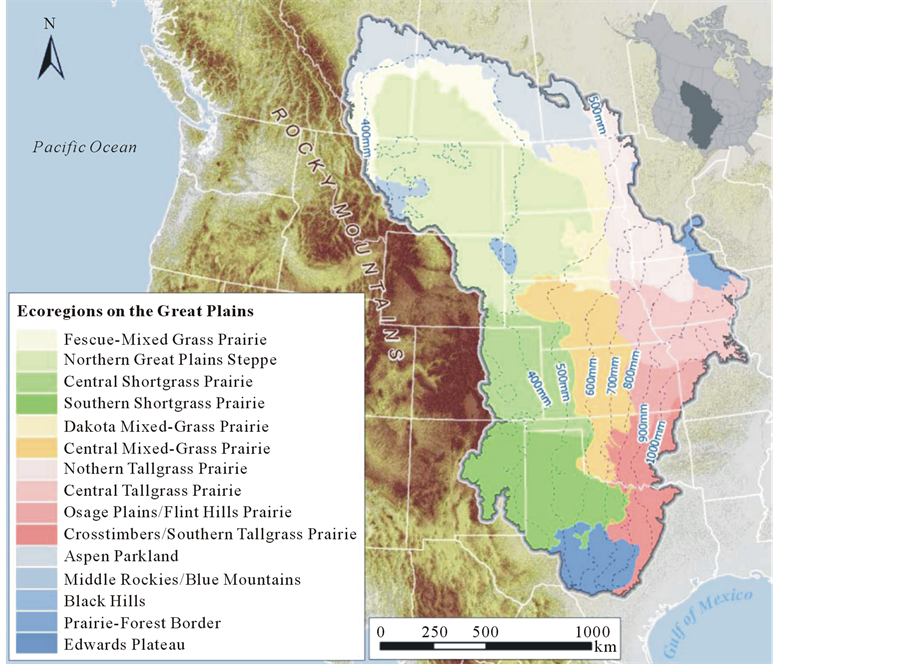
Figure 1. Location and ecoregions of the great plains in North America.
types: shortgrass prairie, northern mixed-grass prairie, tallgrass prairie, and southern mixed-grass prairie.
Grasslands are being fundamentally changed on a global scale, as can be illustrated in their biodiversity. The relationship between biodiversity and the healthy functioning of ecosystems is essential in ecology, which posits that status and rate of ecosystem processes, e.g. productivity and resilience, has strong positive connection with species richness [3] . Biodiversity loss in grassland ecosystems is remarkable, ranking from the most threatened biome to the third most threatened after rainforests and arctic ecosystems [4] . A study [5] shows that by year 2050 another 10% of vascular plant species will be lost. Identification of underlying mechanisms of change in community structure is a pressing and significant task to enable researchers to understand and predict community response to changes in disturbances or climate with a level of certainty useful to management [6] . Composition and structure of grassland ecosystems are constantly subject to many biotic and abiotic factors [7] [8] including the biophysical environment, fire, grazing, and other human activities (e.g. historical and current land-use patterns). Of these diverse factors, there are three which are of great importance and able to override other factors at all trophic levels: rainfall patterns, fire and grazing [8] .
Through evolution history grasslands ecosystems are maintained in conditions appropriate for their productivity and biodiversity in nature, by self-organizing all their biotic and abiotic components in equilibrium [9] . However most of these self-structured, well-evolved mechanisms were fundamentally disrupted and often destroyed by anthropogenic causes (e.g. cultivation and urbanization) before they have been fully understood, mainly because of the intense pressure on grasslands from farming and grazing, resulted from their favorable attributes: flat, treeless, and rich in soil. In fact, North American grasslands are ranked as one of the most extensively disrupted ecosystems on Earth. And in the northern mix-prairie grassland, virtually no major areas of intact habitat remain, with more than 75% of natural habitat being destroyed due to agriculture activities [10] [11] . In USA nearly all land of the Great Plains is cultivated to sustain one of the largest agricultural landscapes [12] . In the northern mixed grasslands, more than half of the area had greater than 80% grass cover in 1880’s before the agricultural settlement [13] . However, current percentage of cultivated land is between 15% in areas with little precipitation and 99% in areas more suitable for growing crops [14] . The Northern Great Plains eco-region is listed as critical and endangered [11] and demands high priority for conservation [15] .
The prairie ecosystem was created and maintained through historical disturbance regimes that sustain its structure, composition, and spatial-temporal dynamics by causing heterogeneity in abundance of individual plant species [16] [17] . It is a well-established theory that disturbances alter various properties of an ecosystem at diverse spatial and temporal scales, most often causing “open space” and patchiness, hence resetting the succession trajectory of the ecosystem [18] , e.g. facilitating re-colonization by previous or other species. In the grassland ecosystem, disturbances such as fire and grazing, or range management (haying) can remove the accumulation of litter content, which in turn positively influence the productivity and plant diversity of the grassland ecosystem [9] [19] . Grazing and burning behaviors are extensively studied in the literature. Because both of them not only have contributed to the evolution history in the past, but also are both forms employed in grassland management practices (cattle grazing and/or prescribed burning). However, for the prairie ecosystem there is in fact a full spectrum of disturbances of different types and scales (Figure 2). Besides grazing and burning, drought is yet another major disturbance on the prairie, which is an under-studied but significant abiotic factor in the grassland ecosystem. In fact, plant growing season, grazing season and fire season, as well as other biotic variables are fundamentally influenced by climate conditions, especially extreme conditions like drought events; mainly because in the prairie available soil moisture functions as the limiting factor and governs the overall physiognomy of the ecosystem in the northern mixed grassland. This makes drought qualified as another significant disturbance and henceforth falls into the scope of this review.
As a result, this paper mainly focuses on the impacts from major disturbances of grazing, fire and drought, even though the authors are aware of other natural and anthropogenic disturbances present in the ecosystem but are regarded less significant in the grassland ecosystem. This review mainly focuses on the mixed prairie in Canada, which is significant for its functioning as the abundant pool of CO2, wildlife and vegetation [20] .
2. Major Disturbances in the Grassland Ecosystem
Disturbances are integral and significant natural components for an ecosystem. For the prairie ecosystem, its dynamics is tightly connected with disturbances. It is not surprising that long-term committed effort has been made to understand the major mechanisms in grassland ecosystems, which has brought us tremendous understanding of each disturbances per se, but unfortunately a systematic view of all major disturbances is not developed, which proves to be a pressing need since grasslands are inherently complex systems. This is also not surprising given the fact of the gradual process of our understanding of grassland ecosystems and their disturbances. It is generally assumed by ecologists and geographers that plant primary productivity and the distribution of biomass are globally determined by climate [21] , and locally modified by disturbances, mainly from consumers
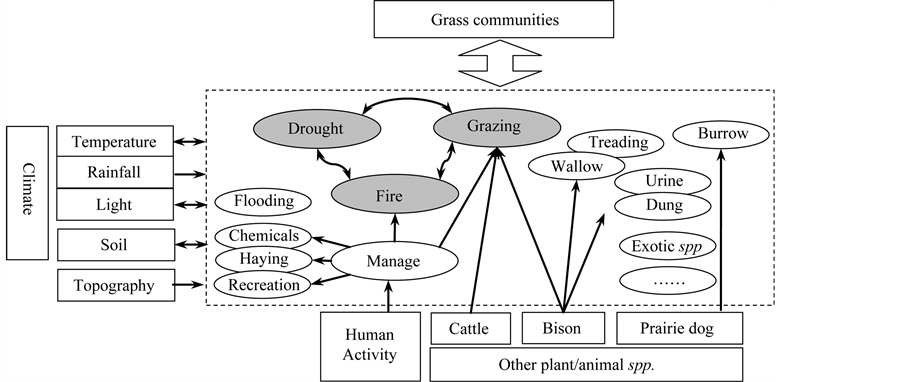
Figure 2. Major disturbances and their interactions in North American mixed-prairie grassland ecosystem (arrow indicates the direction of interactions; oval shapes within the dashed box are disturbances, of which shaded in gray are the main focus of this paper).
and predators [22] . Some critics argue that herbivory and predation fail to depict the whole picture of ecosystems since most plants are inedible, which are not maintained through herbivory but fire. As a result, fire was brought into researchers’ attention and more extensive studies proved fire as an essential force in shaping and maintaining grassland ecosystems. In fact, there are extensive studies on the impact of grazing, fire or drought. Yet most often they are examined separately, having overlooked the fact that in the real ecosystem these disturbances are always closely coupled with one another, leading to both long term and short term effects regarding various properties of disturbances [23] such as distribution, frequency, severity, intensity, time, synergism etc. Disturbances in grasslands rarely occur in isolation, which makes it significant to recognize the interactions between different types of disturbances [24] . Meanwhile, in light of the ubiquitous human impacts on the grass communities, including disturbances such as grazing and fire regimes in the protected prairie, a new level of synthetic and holistic understanding of disturbances on the grassland ecosystem is demanded to be brought into a close attention.
Seeing this, the paper tries to cover major research interests on disturbances of grazing, fire as well as drought, especially their utilities in shaping the mixed-prairie grasslands ecosystem of North America, aiming to offer significant information to better understand the grassland ecosystem and facilitate grassland management practices for diverse stakeholders, researchers and the general public.
The review first examines the well-studied disturbances in grassland ecosystem study: grazing and fire. Then it introduces the role of climate, especially drought, into discussion. The paper then revisits disturbances from the 3 aspects and trying to synthesize them all altogether in the context of the whole ecosystem, with specific attention given to the interaction of all these disturbances.
2.1. Grazing and Its Effect on Grassland Ecosystem
2.1.1. History of Grazing
Grasslands throughout the world are primarily used for livestock production. UNEP report [25] shows that 61% of the arid regions of Earth’s land surface are utilized for ranching. But it still remains a challenge to understand the grazing effect on biophysical factors and the ecosystem processes in grasslands [26] . Before Euro-American’s settlement, the whole prairie region saw the First Nations people living among “buffalo in great droves” [27] , which literally “blackened” the prairie and can be dated back to at least 12,000 years [28] . For tens of thousands of years the history of grazing on the prairie has been centered on bison herds. However starting from 1800 it only took 80 years to change the entire picture, with all the bison wiped out completely from the Canadian Plains, and First Nation people living in reserves. We were responsible for such a calamity ever happened on the prairie history, thanks to the population growth, the buffalo product trading and the construction of the Northern Pacific Railroad. Human predation is the only likely reason of bison’s extinction, which exceedingly eclipsed other possible explanations including drought, disease and competition from other herbivores. Human hunting pushed crowded bison herds to restricted and fragmented habitat which resulted in overgrazing and degradation of the grasslands. To make the situation worse, horses was introduced to the Northern Plains at 1794, which soon were adopted by many First Nations people into their culture, to kill (perhaps overkill) bison effectively, and also to compete with bison for grass. Horse was widely used until 1940s when it was replaced by tractors. During 1880s the population growth led to the increase in domestic livestock production, causing grazing pressure on the prairie. 1900s on the prairie saw the largest ranching company in the world [28] . As settlement increased, most of the prairie was cultivated to produce grain, leaving the rest unsuitable prairie either untouched, or made into large grazing pastures.
Grasslands National Park (GNP) of Canada is one of the pristine grasslands which still stand in the world, with grazing excluded since the park’s foundation in 1986. Later in 2006 grazing was re-introduced to the park area for ecological integrity monitoring purposes. Light and moderate grazing pressure have been carried out in the park, which is lower than the recommended stocking rate of this type of region. Meanwhile, the surrounding areas outside park were still under grazing pressure from livestock. The relative integrity of native pristine prairie in GNP makes it the hotspot for many scientific experiments and researches.
2.1.2. Impact of Grazing
Impact of grazing mainly comes from large herbivores in grasslands. In general herbivores consume 15% to 60% of annual above-ground NPP, and 5% to 15% of below-ground NPP [29] , of which 10% are grazed by small herbivores and the rest by large herbivores [18] . It is proposed that large herbivores as part of native fauna co-evolved with the grasslands, playing a significant role in shaping the structure and composition of the grassland ecosystem in the past as well as the landscape we see today. In the history of North American grasslands, large herbivores included Bison, elk, wild horses, moose and many others. There were probably tens of millions of bison functioning as the keystone species but were later wiped out by massive early European settlers in 19th century. Nowadays most part of the prairie has become managed pasture grazed by domestic grazers.
Grazing is detrimental from the perspective of individual plants in the short term, but becomes more complex when viewed from the perspective of the ecosystem in the long term. A full understanding of the relationship between grazing and vegetation required efforts in both perspectives [30] . At the individual plant level, grazing removes parts of the plant (leaves, flowers, stems, and roots), resulting reduced photosynthetic surfaces, seed production, as well as water, carbohydrate and nutrients supplies etc. This compromises the capacity of the plant to capture solar energy, which leads to decrease in its competiveness and productivity. However, there are species taking advantage of grazing to thrive in grasslands. For example the seed production of scarlet gilia doubled in grazed communities than in ungrazed ones [31] . It’s arguable about the long term relationship between grazing and productivity, given diverse hypotheses and inconsistent results from the literature. Some (e.g. [32] ) claim overgrazing caused nutrient loss, and degradation of the ecosystem, showing strong negative impact of grazing to grasslands. Meanwhile, light to moderate grazing are found to be beneficial to grasslands [33] , or improving herbage production [34] . However, some researches show no benefit of grazing for plants (e.g. [35] ). Meanwhile Milchunas [36] found that production was highest in ungrazed treatments, and decreased as grazing intensity increased in short-grass prairie.
Grazing can also alter the spatial heterogeneity of vegetation, influencing ecosystem processes and biodiversity. Grazing lends itself as a significant tool for ecosystem conservation, given its influence on maintaining habitat structure and biodiversity [37] . Grazing influences plant diversity in many ecosystems [38] , but the mechanism is still not certain. Grazing may promote plant diversity by reducing the pressure of competition between different plant species at fine scale [37] , but it also may cause the invasion of exotic species and extinction of native species [39] . Meanwhile, grazing influences plant diversity by causing heterogeneity in resources at various spatial scales, through either general or selective grazing [40] . Effect of grazing on plant biodiversity depends on the environment. In more productive ecosystem (temperate grasslands or tall grasslands), there’s positive correlation between grazing and biodiversity. Whereas in arid and very saline environments, such relationship does not hold and even becomes negative. Grazing experiment in North American tall grass prairie reduced biodiversity on sites with poor soil conditions, but promoted biodiversity on sites with rich soils [41] . Table 1 shows how the effect of grazing varies across environmental gradients.
2.1.3. Grazing Hypotheses
Hypotheses have been developed to explain the relationship between grazing and biodiversity, by introducing the concepts of local colonization and local extinction [41] . Local colonization is achieved through dispersal and establishment of individuals from species pools at larger spatial scales. Local extinction is caused by competitive exclusion. Local colonization is facilitated by causing gaps in grass communities from grazing, as well as

Table 1. Effects of large herbivore’s grazing on grassland biodiversity across different grassland environments (adapted from [41] ).
aResource competed for by ungrazed plants; b+ indicates increased biodiversity resulted from reduced extinction. − indicates decreased biodiversity resulted from higher extinction rates; c+ indicates increased biodiversity resulted from enhanced local colonization. − indicates decreased biodiversity resulted from reduced local colonization.
other disturbances from herbivores like trampling and digging. Local extinction can be resulted from intense selective grazing, deposition of urine and faeces, wallowing, and soil disturbances from herbivores. It is suggested that higher biodiversity is expected when local extinction rates of species stay lower than colonization rates. Meanwhile these two processes are not necessarily mutually exclusive. In the grazing scenario, dominant species (competitors) is reduced, which meanwhile facilitates the colonization of other plant species in the disturbed community. So the interaction between these processes enhances grazing effects on the biodiversity.
In order to understand the impact of grazing on grassland ecosystem, several theories have been proposed, including range succession model, herbivore optimization hypothesis (e.g., Milchunas-Sala-Lauenroth, or MSL model), and State-and-Transition (ST) Model.
Range succession model [42] hypothesizes that there exists a single equilibrium vegetation composition (climax vegetation) for each grassland ecosystem in the absence of grazing, which supposed to have the highest sustainable productivity and to be the most resistant to disturbance.
Herbivore optimization hypothesis [43] proposes that an intermediate level of grazing shows positive effect on ANPP and biodiversity. However this hypothesis is quite controversial, especially when we observe the fact that grazers can increase their “harvest” by repeated grazing on the “grazing lawns”, one example of this phenomenon can be found in the theory of “pyric-herbivory”. But still this hypothesis is supported [44] in grasslands featuring long evolution history and low productivity.
Grazing effects can be explained by Milchunas-Sala-Lauenroth (MSL) model [45] regarding the relationship between changes in biodiversity caused by grazing along gradients of moisture and historic grazing regime. MSL model is proposed in light of the intermediate disturbance hypothesis, based on research done at 4 sites with distinct extreme conditions along 2 gradients: moisture and historical grazing regime, including the shortgrass prairie in North American Great Plains which was semi-arid grassland with a long grazing history. MSL explains that for semi-arid grasslands, changes in species composition and biodiversity caused by grazing are relatively insignificant or moderate, depending on historic grazing regimes. However in sub-humid grasslands, such changes are relatively more significant regardless of historic grazing regimes. By examining the effects of grazing around the world, Milchunas & Lauenroth [44] showed that changes in species composition caused by grazing are related primarily to Above-ground NPP (ANPP), secondly to the historic grazing regime, and thirdly to the grazing intensity. Species composition change increases with higher ANPP (always connected with higher moisture level) and longer, more intense grazing history. In fact, field experiments confirmed MSL’s hypothesis, recognizing that plant biodiversity was increased by grazing from large herbivores [41] [46] . Such increase in biodiversity can be explained by enhanced local colonization of new species in productive grassland, which was caused by lowered light limitation due to gaps being opened up by grazing in otherwise closed grass canopy.
MSL propose that an ecosystem reach at its climax stage at intermediate levels of disturbance, whereas it shifts away into declined conditions both at low and high levels of disturbance. So the framework of MSL model rejects the potential existence of stable vegetation state alternatives, which may occur diversely when changes in vegetation caused by grazing has surpassed a threshold tipping point which is beyond the resilience of the system.
By proposing potential vegetation state alternatives in multiple equilibria, State-and-Transition (ST) Model was developed [47] , which recognizes the irreversible transitions in grassland, as well as alternative equilibria of the disturbed system. The utility of this model is the power of explaining vegetation’s response from grazing effect in grasslands with relatively short grazing history. For these grasslands, plants have evolved through history to adapt to either productive or non-productive environment, having little or no pre-adaptation to grazing. When grazing is introduced those grazing-sensitive species may not be able to survive and become locally extinct, thus resulting in the decrease in biodiversity. Especially for low productive (e.g. semi-arid) grasslands, high grazing pressure may lead to serious ecosystem shift, which is the exact situation for most part of grasslands in Mongolia since its economic and political shifts in 1990s [48] .
2.2. Fire and Its Effect on Grassland Ecosystems
2.2.1. History of Fire
Fire is one of the oldest natural phenomena on the prairie and functions as an inevitable and essential ecological force throughout the prairie history. Grassland ecosystems not only tolerate fire, but also encourage it and depend on it to survive. Before European settlement, the prairie ecosystem was maintained through fire, caused either by nature or aboriginal people [49] . In fact lightning strokes account for just 10% of fire occurrences on the Great Plains [50] . This makes human the primary source of ignition. Meanwhile mankind has been the vector of its propagation, as well as the most significant modifier of fire regime. Historical journals noted that most Indian fires occurred in valley grasslands and adjacent forests [51] . Native Americans periodically burned grassland intentionally to modify local habitats and to aid in hunting activities by both driving and attracting wild game [51] [52] . A study [27] shows that the extent of grassland reached farther north in 1800s than today, because of the historical fire regime. However, fire occurrence on the prairie has greatly declined ever since, when early Euro-Americans started to populate and cultivate the Great Plains. This post-settlement decline of fire events was mainly caused by active fire suppression and reduction of fuel from grazing by domestic livestock. Absence of recurrent fire since the settlement significantly altered the trajectory of the environmental history of the Great Plains, resulting in accumulated litter, declined productivity, and woody species invasion, etc. Fire regimes were forever modified and vegetation cover was reshaped fundamentally on the Great Plains [53] . People once regarded fire suppression as equal to fire management [54] . Such misconception was carried on through centuries until 1930s, when prescribed fire as a management tool was reintroduced into North America (starting from southeastern USA), backed up by well conducted scientific researches [55] [56] . The role of fire in the structure organization and evolution of grassland ecosystems is widely acknowledged [6] . Nowadays fire is well recognized as a vital component in shaping the structure of grassland ecosystems. It is employed as a management tool throughout all areas of North America in grassland ecosystems [57] .
The prairie’s susceptibility to fire can be mechanistically explained by its natural environment of fuels, weather and topography. The alternating wet-dry landscape of the Great Plains makes itself susceptible to fire. The limiting resource is soil moisture on the Great Plains, especially northern mixed prairie, where droughts are common phenomena following short periods of precipitation. And precipitation is often in forms of lightning storms, concentrated in June and July. Plentiful and extra precipitation received in a wet season makes grasses grow rapidly and abundantly. Then the following dry year and sometimes droughts are able to dry the grasses, making them easily flammable. Meanwhile since the prairie features flat or rolling landscape, when a fire has started it can be carried away by dry winds and spread quickly, with few obstacles to slow down or stop its progress [58] .
2.2.2. Impact of Fire
Fire’s impact on terrestrial ecosystems is documented [59] as: 1) shaping global biome distribution, 2) maintaining the structure and function of fire-prone communities, 3) acting as an evolutionary force, 4) being employed as one of the first tools by humans to reshape the world. Fire has significant impact on the fauna and flora in the grassland ecosystem, in both direct and indirect ways. Direct effects include mortality of individuals, which are short-term. Indirect effects, such as species composition and changes in habitat, are long-term, which are not easy to observe and evaluate but usually more important than short term effects given their profound impact on the ecosystem, especially in the aspects of vegetation recovery and biodiversity.
Fire, coupling with other numerous factors, i.e. topography, soil, fauna (insects, herbivores), together with herbaceous plants [57] [60] , can restrict the encroachment of woody plant (trees, shrubs), release nutrients bound up in organic matter, accelerating the rate of decomposition in the soil, so as to maintain the establishment and the stability of grasslands [6] [57] [61] . Descriptive studies [62] [63] show that fire occurrences decrease herbaceous production for one to three years. Meanwhile, herbaceous response is influenced strongly by precipitation. Fire also reduces woody plant cover and promotes herbaceous dicotyledons. Plant species in semi-arid grasslands are more likely subject to fire season and frequency than fire behavior [64] .
The reestablishment of historical fire regime is fundamental to the ecological restoration of grasslands on the Great Plans [61] . This is especially the case for ecosystems with a long history of fire, such as the prairie ecosystem, where human’s fire suppression caused series of negative consequences. A study [65] [66] in tall grass prairie shows that 50% plant species has been lost due to active fire suppression and the authors argue that this is common in grasslands elsewhere. Taking fire out of grassland ecosystem makes it threatened by the encroachment and dominance of woody plants. Even dramatic species loss may occur when fire suppression results in complete biome change, from grasslands to forests as shown in a study [67] . In the plant level, fire causes various levels of reduction in above ground biomass. In the community level, fire results in potential local species loss, changes in species composition. Meanwhile, fire affects vegetation by lowering the soil albedo, raising soil temperature, as well as releasing nutrients from organic matter. Study of the conifer forests of southwestern North America [59] explicitly describes that: human settlement during the early 20th century, with the policy of total fire suppression, is by itself an experiment on how fire controls vegetation structure, and has resulted in near-total fire extinction. This in turn resulted in major shifts in ecosystem structure and function: 1) tree density increased significantly; 2) major losses in the herbaceous under-storey; 3) losses in species diversity; 4) fuel accumulation, a major potential threat. Besides the use of fire as a management tool in livestock operations, for the purpose of brush control and forage quality improvement, people also employ fire to control external parasites [68] .
Researches show that fire’s seasonality has greater impact on the recovery of vegetation. In the short-term, burning during the growing-season appears to reduce fire severity but exerts greater impact on grass communities (opposite for soil crusts) compared to burning during the dormant-season. Dormant-season fire on the other hand, is less detrimental to grass communities (opposite for soil crusts) than growing-season fire. Shortgrass steppe can recover from fire in 3 to 30 months, dependent on the fire season [61] . Ford et al. also reported that in general, grass cover recovered quickly from the fire treatment, and the long-term effect of fire was neutral. Burning during the dormant-season had little effect on grass cover, even in short term as soon as two months; whereas growing-season fire showed negative impact on grass cover for as long as two years after fire. However, some studies [69] -[71] indicated mixed results when evaluating the impact of spring fire on buffalograss and blue grama, over a time frame of 3 months to 16 years. Often, early-spring burns (March) produce neutral or positive responses; and late-spring burns (May) produce negative results. Fall burns led to more yield than did spring burns. Negative, neutral and positive responses to fire were evident in both season-long grazed areas [69] , and areas protected from domestic livestock grazing [62] . Shortgrass prairie ecosystem recovers relatively quickly from fire disturbance [53] . Vegetation cover, arthropod, mammal species richness treated with dormantseason fire recovered in approximately two months and showed no significant difference from untreated communities. Meanwhile, we must be cautious when interpreting the results from different researchers on the impact of timing of fire on the grassland ecosystem, since impact of fire varies for different types of plants according to their active growth season, with C4 plants least vulnerable to the dormant-season fire and most vulnerable to the growing season fire [61] .
2.2.3. Restoration of Historical Fire Regime
Historical fire regime functioned as an important natural process for the grassland ecosystem, which was irrevocably changed, making study on fire ecology a pressing issue. It is challenging to study historical fire regime, by means of either field experiment or modeling. Mainly because reliable long term historical records of fire occurrences on the prairie are not available, due to at least three reasons: 1) throughout history fire’s role in the grassland ecosystem has been ignored, resulting in little records of fire occurrences, except occasionally sparse burning events can be traced in personal journals or some other sources, however with low data quality; 2) Fire occurrences can be identified accurately based on burned scars on trees [51] ; yet there are not enough trees on the prairie to provide such data; 3) Charcoal fragments is another way of investigating fire’s impact. Umbanhowar [72] sampled some charcoal remains from lake sediment cores to study the fire frequency of preand post-settlement period on the northern Great Plains. However, such research requires tremendous data collection effort and also brings negative disturbances to the ecosystem.
Many researches are focused on the impact of currently manmade fire regimes by introducing prescribed fire, with the aim of revealing and restoring historical fire regimes for the benefit of grassland management. Brockway et al. [73] found that dormant-season fire may be the preferable method for restoring fire in shortgrass steppe ecosystems in the southern Great Plains where fire has been excluded for a long period of time. However, the drawback in their experiment was that post-treatment measurements were taken and analyzed merely 6 months later. More research is needed to investigate the long-term fire regime and vegetation response in the grassland ecosystem. Meanwhile, grasslands show significant variation of heterogeneity in its species composition and physiognomy, which should be taken into account when evaluating the impact of a fire regime. Guo et al. [74] found that aboveground dry biomass, plant moisture, and dominant species together with plant forms are different in cool seasons and warm seasons on the Tallgrass prairie, which demand more investigation on how the senescence stage of grasslands and the seasonality of fire interact with each other within a specific ecosystem like the mixed prairie.
Despite the efforts of restoring historical fire regime, quantifying long-term shift in fire regimes is important to understand fire ecology in the grassland ecosystem. And prescript field experiment proves to be an effective tool to reveal the mechanism of burning in the grassland ecosystem. However, assessing fire is still challenging, especially on the prairie due to: 1) high intraand interannual variation of grass communities, which is determined by a myriad of factors from climate to other disturbances; 2) lack of appropriate satellite imagery or aerial photos for such purposes (for an example of evaluating burned areas in grassland ecosystem using remote sensing technique see [75] ).
Most researches on fire regimes are dealing with particular local landscapes and ecosystems. There is still no clear understanding of the mechanisms behind the interaction of fire and biophysical environment, such as the relative importance of ignition, dry periods, properties of grasses as fuel, and topographic barriers to constrain fire spreading [59] . But we know that different fire regimes favour different plant attributes and similar fire regimes favour similar plant attributes. Grassland ecologists around the world find similar plant attributes with quite similar fire responses [76] . Fire’s impact, its behaviour and its consequences as well as vegetation response in the grassland ecosystem, is to some extent determined by local climate.
2.3. Climate Effect
Despite of influences from grazing and fire, climate variables have fundamental impact on the grassland ecosystem, mainly through the limiting effect of key climate variables such as temperature and precipitation as well as particular climate events, e.g. drought and flood, induced from configurations of these climate variables. In fact, the function of an ecosystem is mainly governed by environmental forces, especially climate, which dictates the photosynthetic process and hence the primary productivity of the ecosystem [77] . Primary productivity in terrestrial ecosystems is a function of temperature and also significantly correlated with precipitation [78] . In fact, the ubiquity of grassland ecosystems can be explained by the adaptation of grasses in localities of a wide and diverse types of climate conditions: where annual precipitation is between 150 and 1,300 mm and average annual temperature between 0˚C and 30˚C [58] [79] . Therefore habitats for grasses can be found along the full precipitation gradients: from dry desert to moist forest (Figure 1). At one extreme, in dry areas with little precipitation grasslands tend to degrade into desert. Moving toward the other extreme, there exists a general trend of increasing cover of trees and bushes as the precipitation increases [80] . Henceforth, climate determines overall attributes of grassland ecosystems. For the mixed prairie on the Great Plains in North America, where changes in elevation are not prominent, precipitation plays the most significant role in the vegetation changes. Indeed, regional precipitation patterns determine the large scale distribution and characteristics of grasslands [18] , shaping mesic tall-grass prairie in the eastern wet areas (625 - 1200 mm) and short grass steppe in the southwestern USA (260 - 375 mm) as well as semi-arid mixed grassland in the Canadian Prairie Provinces (375 - 625 mm), which are classified by ecologists based on the stature of grasses, and tightly related to annual precipitation. The general distribution pattern of different grasses can be well explained by its natural geography, since precipitation on the Great Plains depends on moist air masses originated from the Pacific Ocean in the west coast and Gulf of Mexico in the south interacting with cold air from the Arctic and mountain areas [18] .
Through history native grasses have been highly evolved and adapted to extremely unreliable climate, especially frequent and long drought events. The erratic climate even afflicted High Plains Indians suffering from local environmental degradation caused by its unpredictable precipitation [81] . Drought (“dustbowl”) happened on the Great Plains in early 1930s was another example of illustrating the extent of impact of climate events, which severely damaged the ecology and agriculture throughout the prairie regions [82] . However it was in fact not the most extreme situation happened in the prairie history. Records show that before European settlement, droughts on the prairie were of long duration and much extensive. In northern prairie, fossil records discovered in central Saskatchewan indicate a persistent drought for more than 70 years; whereas in southern part of Great Plains evidence suggests prehistoric drought lasted up to 300 years [18] . In fact grassland ecosystems commonly feature variable climate conditions, where periodic droughts are well known phenomena [58] . Fluctuation in precipitation is the main reason of variability in grassland environments. Research [83] confirms that among other biomes grassland ecosystems have greatest interannual variability in productivity, and are strongly influenced by precipitation. The influence of precipitation on productivity in grasslands and other biomes in North America has been well documented [83] [84] . Particularly precipitation events in the range of 15 - 30 mm can explain most variation in grassland productivity, because this range accounts for most variation in precipitation and effective moistening of the soils to promote productivity.
Even though frequent drought events are widely known and viewed as a normal disturbance for grasslands, long term and severe droughts however may cause severely negative impact. Drought has far-reaching impact on the grassland ecosystem, shown in Figure 3.
Grassland productivity following abnormal dry years shows a clear lag in recovery, even though precipitation after the dry year may be normal or above normal [45] . This lag is proposed to be the result of interactions among grass communities and ecosystem processes [85] , which explains that drought conditions results in mortality in plant tissues which impacts plant’s capacity to recover even in cases of high resource availability [84] [86] .
Furthermore, drought, coupling with other climate and topographic variables, has indirect impact on grassland ecosystem through affecting other disturbances, such as fire regimes [87] . This has been studied extensively in forestry. Significant variability of fire regime existed for forests, with mesic forests governed by mixedto stand-replacing fire regime of 400 to 500 years [88] [89] , while drier forests maintained by lowto mixed-severity fire regime of approximately 15 years [88] . Also it has been found that there exists a gradient in fire regimes from north to south as a function of meteorological variables, i.e., temperature and precipitation patterns [87] .
2.4. Compound Effects of Major Disturbances
2.4.1. Fire and Grazing
Fire and grazing are interacted closely which makes it necessary to investigate the two together. In fact, both actively impact on a large proportion of the earth’s ecosystems [44] , playing critical roles in both establishment and maintenance of grasslands and savannas [1] [45] [90] . Their interaction contributes to the heterogeneity and productivity of grassland ecosystems.
There are intricate connections between fire and grazing. To some extent fire and grazing behave similarly. And fire was proposed as a non-selective, globally-distributed “herbivore” [59] [91] . Though here “non-selective” is clarified as regarding to trait composition instead of species composition [59] . They both act as top-down drivers what take in grasses (complex organic molecules) and convert them into organic and mineral products. And both are selective forces acting upon grass communities, which favor different plant attributes and produce heterogeneity both at landscape level and community level [76] .
Spasojevic [91] specifically investigated long-term (1984-2006) field experiment to compare effects of fire and grazing. They failed to verify whether grazed grass communities and burned communities would converge or not. But they had some interesting conclusions: 1) removing fire from frequently burned grassland may eventually make it more like unburned ones, depending on fire frequency, and with a certain time lag; 2) adding fire into previously unburned grassland may quickly make it like the frequently burned ones, again depending on fire frequency. They found out that fire with intermediate frequency, i.e. once every 4 years, worked best, which confirmed a hypothesis made by Hulbert [92] about the historical fire frequency; 3) both burned and grazed communities at intermediate levels share similarity in high abundances of annual C4 grasses compared to the rest communities. This is consistent with other researches reporting the quick recovery of C4 plants after either disturbance, which proves the competitive advantage of C4 plants over C3 plants in high light environments caused by either of these disturbances [93] .
Even though both fire and grazing put similar pressure on grasslands, they are essentially different. Unlike grazing, fire regularly consumes both dead and living materials, with broad “dietary” preferences, often consuming plant inedible or not favored by herbivores. High frequency burning has more selective pressure on trait
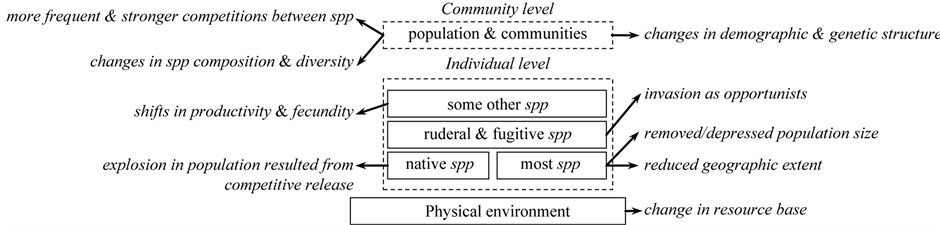
Figure 3. Impact of drought on the grassland ecosystem (adapted from the table by [18] ).
composition of grassland communities than selective grazing [91] . Also, chronic fires tend to have greater impact on species and trait composition than chronic grazing at light to moderate levels, because these two disturbances are different in their selectivity [94] [95] . Meanwhile, unlike grazing, burning or “combustive consumption” by fires acts upon physical properties of available fuels, which may cause rapid and severe impact quickly across large areas [96] . Their impacts can be different spatially, with fire causing relatively uniform impacts and grazing associated with higher spatial heterogeneity [94] [95] . Some researchers reveal that fire and grazing have different effects upon the species composition in grassland ecosystems [16] [65] [97] [98] . A study on the mesic tallgrass prairie [94] shows that frequent burning acts as a non-selective force which decreases biodiversity, with a contrast to selective grazing event which increases biodiversity.
Fire and Grazing are two major disturbances on the grassland ecosystems and more often than not they work together affecting ecosystem processes. Each of the disturbances has been extensively studied individually in the literature (see a review from [99] ). However, the interaction of them may be more ecologically significant than their independent effects in isolation. Given this situation “Pyric herbivory” was proposed recently to evaluate the compound effect of grazing and fire, which is defined as grazing driven by fire [99] [100] . This theory argues that the interaction between burning and grazing involves positive and/or negative feedbacks in a closely coupled fire-grazing system, generating spatial and temporal heterogeneity as well as new dynamics in the ecosystem which are absent when grazing and fire are examined separately.
Pyric-herbivory is being tested and applied [68] [100] -[102] due to its power in explaining the formation of spatial heterogeneity caused by grazing and fire in the ecosystem. The theoretical foundation of pyric-herbivory is the inverse relationship between forage quality and elapsed time since last burning event. For example, Archibald et al. [103] used the term “magnet effect” to refer to the attraction of herbivores to recently burned areas, a phenomenon documented with numerous animal species throughout the world (see the review by [101] ). Allred [101] put the “magnet effect” further by claiming that herbivores prefer recently burned areas and avoid areas with greater time since fire, regardless of the size of landscape, herbivore species or proportion of area burned. A field experiment was conducted [95] , showing that by burning 1/3 of a pasture yearly cattle’s grazing time on burned patches was 3 times that on unburned ones. Meanwhile, McGranahan et al. [104] verified pyricherbivory by investigating the spatial heterogeneity on five grasslands through field experiment. Their result did not consistently confirm the hypothesis claimed by pyric-herbivory, though for some grassland the increase in heterogeneity and biodiversity could be successfully explained using the theory. They suggested that pyric-herbivory work only if fire is the primary driver for grazing site selection of herbivores. That is, pyric-herbivory applies when fire’s influence is strong enough to override other environmental factors contributing to selective grazing at the landscape level [38] [101] .
2.4.2. Coupling with the Climate
There is close connection between grazing and climate variables, particularly precipitation fluctuations. However, the long-term relationship between grazing and climate is still not clear. In the mixed prairie grassland, Yang [105] found that 70% of variation in grassland productivity could be explained by the amount of precipitation during the growing season (from April to August), both for grazed and ungrazed communities. This agrees with the previously mentioned hypothesis on climate being the main determinant of grassland productivity at the global/regional scale. She also observed significant differences in Normalized Canopy Index between grazed and ungrazed communities in years with above normal precipitation in growing seasons (wet years), but not in dry years. This confirms previously mentioned hypothesis on disturbances modifying the general distribution of grass communities at the local scale, with grazing and exceptional precipitation as disturbances in this context. Furthermore, Ellis and Swift [106] showed that climate could override grazing effect completely regarding annual production, which has been confirmed for the northern mixed-grass prairie [107] . However, Fuhlendorf [108] argues that grazing effect establishes the long-term direction of change in vegetation, and that climate explains short-term rate of change.
In mixed-prairie grassland, fire is inherently connected with its climate, which can be easily seen from the concept fire regime. Fire regime was introduced by Gill [109] . Bond et al. [59] modified the concept of fire regime to include 5 aspects: 1) fuel consumption and fire spread patterns, 2) intensity, 3) severity, 4) frequency and 5) seasonality. All these aspects alone or together may have profound impacts on terrestrial ecosystems. But the season of fire is more interesting since it brings climate variables into account explicitly. Fire season is governed by the coincidence of ignition and low moisture in the ecosystem. This condition usually happens to be the driest time of the year. When dry, fuels of semi-arid grasslands may favor high rates of fire spread [110] , whereas green grasses or actively growing grasses promote a discontinuous or patchy fire regime [111] . The high moisture content of green, living plant tissue makes it more difficult to burn, than dry plant or litter which ordinarily promotes fire spread. Therefore, fire intensity and severity during the dormant season tend to be higher, resulting in more widespread damage and mortality to exposed surface crusts than growing-season fire. Whereas, fire during the growing season has been shown to reduce regrowth in vascular plants because large portions of photosynthetically active tissues are killed [112] .
Launchbaugh [63] examined a spring wildfire in shortgrass prairie when the soil moisture was low, and found that fire caused short-term declines in plant biomass. It took three growing seasons for a burned grass community to return to a level comparable to the unburned state. Similar results of burning in prairie were reported in west-central Kansas [86] . Following a wildfire in New Mexico when the moisture balance was more favorable, [62] found that biomass production of blue grama was reduced only by 30% during the first year growing season following the burn. Biomass of the grass community returned to pre-burn status with above-average precipitation the second year after the burning event. Prescribed fire in Texas during years with above-normal winter and spring precipitation showed considerable tolerance of fire for buffalograss and blue grama, with no loss at all in the yield at the end of the first growing season [70] . Research [57] shows that the tolerance of most grass species to fire in the shortgrass prairie, under different moisture regimes, appears to be similar to the species mentioned above.
3. Challenges
Even though significant amount of studies on disturbances have been conducted extensively and thoroughly, controversial findings and unexplainable phenomena are still challenging researchers in grassland ecosystem, including the definition of disturbances and their ecological mechanisms, as well as scaling, timing, complex coupling of disturbances, etc.
The concept of disturbance is not well defined semantically. Fire is an active agent of ecological dynamics in grassland ecosystems, either by being applied or by being withheld [50] . Fire has impact on grassland ecosystems in various ways. In this sense fire is obviously regarded as a disturbance. However given the significant contribution of fire in maintaining grassland ecosystems, it can be argued that the absence of fire rather than fire itself constitutes a disturbance [113] .
Meanwhile, identities of disturbances can be scale-dependent. A factor regarded as disturbance in the local community level may not be true in the landscape scale. Chaneton et al. [114] found that in grasslands in Argentina, grazing increased community richness by promoting the invasion of exotic plant species, yet when investigated at the landscape level it actually reduced compositional heterogeneity. Furthermore, disturbances cause a cascade of effects in an ecosystem for a long period of time and in various trophic levels, which makes it hard to understand its long term effect accurately, especially given the fact that most grasslands available in the world have already been fundamentally altered, being different from its historical disturbance regimes. One promising way of investigating disturbances at various scales is Remote Sensing (RS) analysis. Large scale disturbances such as drought, fire and grazing can be captured by RS imagery, which makes it possible to investigate those disturbances using RS modeling. Yang [105] has tried to monitor impact of grazing at light to moderate level using RS approaches and found that canopy height and the ratio of photosynthetically (PV) to non-photosynthetically (NPV) active vegetation cover were most sensitive biophysical parameters for detecting changes in vegetation cover caused by light to moderate grazing intensities. Seeing this encouraging result, we are convinced that RS will play a significant role in the study of disturbances in near future, given the fact that a great variety of popular remote sensors are in service or being developed sustainably with more advanced features and maintains cost-effective at the same time, especially in the area of Unmanned Aerial Vehicle (UAV) applications.
Long term impact of disturbances is difficult to evaluate. Responses of grass communities under disturbances show both short-term and long-term effect. Short-term effect always tends to cause individual mortality, whereas long term effect is related to the whole ecosystem, e.g. habitat fragmentation, diversity and heterogeneity of species in all trophic levels etc. Long-term effect is always more essential and significant to help us understand the trajectory of the grassland ecosystem but it is always a challenge to investigate, due to limited resources, inappropriate temporal resolution and time span, as well as other reasons. The massive information captured from remote sensors may provide a quality data source yet appropriate and robust data mining and synthesizing techniques also need to be developed accordingly.
Another challenge of evaluating impact of disturbances is the complicated responses from grass communities. It is always arguable trying to define the health status of a grassland ecosystem. Popular proxies such as productivity, biodiversity and heterogeneity etc. all contribute to the system’s health, but at the same time have their own specific limitations, and sometimes even contradicting with one another. So how to reconcile those together or develop more effective response variables remains a challenge.
There is challenge from grass communities, which evolve across time to adapt themselves to fit and even take advantage of disturbance regimes. It is known that through co-evolution process grasses on the prairie often have growing tips beneath soil, which help them escape drought condition, as well as mitigating impact from burning and grazing. Fires on the prairie burn rapidly and the soil lends itself as the insulator to stop the penetration of heat into the soil and cause damage to the growing tips of grasses [9] . Similarly, herbivores feed on aboveground biomass, but leave the growing tips intact which enables grasses to recover quickly once the grazing pressure is removed. It is not easy to find optimum disturbance regimes to maximize the diversity of those native species without looking into their life history as well as the secondary succession of local communities. Meanwhile it is not clear if grass communities are undergoing a different evolution trajectory to fit the current disturbance regimes, especially given the context of climate change and extreme weather conditions brought forth by humans.
In addition, we always try to preserve and restore the grassland to resemble its historic condition. However, few areas of most grasslands today have kept their historical disturbance regimes, given the fact that almost all the grasslands have undergone extensive and abrupt disturbances since human’s activity, with many of them don’t support historical biotic agents of disturbance. This makes it a challenge to understand the historical disturbance regime. Grasslands were once covering most surface of the earth in the past, with their disturbance regimes established through evolution, maintaining the ecosystem health in an effective way. However most grasslands today are either fragmented or managed heavily and extensively, which fundamentally altered the original disturbance regime. Efforts are being made continuously to understand the significant mechanisms of disturbances in grassland ecosystems, without a whole picture of historical conditions as a reference. For example American Bison once roamed in the prairie region in massive herds, functioning as a keystone species; but were driven to extinction by hunting and slaughtering in 19th century [27] . Lots of other native species share similar fate, especially under the pressure of the invasion of exotic species. What is seen in a current grassland ecosystem is a new trajectory significantly different from what was happening in the past where the whole system was in a dynamic equilibrium.
Although we cannot examine the grassland in its historical condition, researchers have devised various ways to hypothesize and deduce the optimum disturbance regimes which resemble the condition of the system in its past. On one hand, fire and grazing experiments have been investigated extensively by researchers to understand the best disturbance regimes. In GNP bison was re-introduced back to the prairie in 1990s, with an effort to resemble the historical disturbances. Though field experiments give us insights on actual details of disturbances, they are always in short term, when compared with the geological time scale with the force of evolution clearly visible. On the other hand, researchers long ago have started to build physical models which are able to simulate the underlying ecological processes supporting the ecosystem. And yet challenges remain as to fine-tuning the parameters to make those physical models reliable.
Finally, the compound effect of grass communities and disturbances is a challenge to investigate. Grassland ecosystem is inherently a complex system which cannot fully understand by reducing it into different separate components. In fact according to holistic theory, a complex system cannot be reduced to the sum of its parts, simply because “the whole is greater than the sum of its parts”, i.e., the information carried by elements taken from the whole is much richer than the elements taken in isolation. Extensive studies have been conducted to single out one or a few disturbances for various reasons; yet real ecosystem has all possible disturbances happening concurrently, closely coupled with one another to shape the ultimate physiognomy of the system. A potential solution may be modeling the grassland ecosystem using complex system theory. Multi-Agent Geosimulation frameworks has been used for this purpose, with their potential to easily simulate spatial processes, and incorporate the interaction of components and stochastic processes. In fact, ABMs have led to a paradigm shift in ecological modeling [115] , but currently they are still in their experimental stage with their own challenges ranging from development to calibration and validation.
4. Conclusions and Future Study
“The systems we isolate mentally are not only included as parts of larger ones, but they also overlap, interlock and interact with one another” [116] . In fact, grasslands are inherently heterogeneous in both spatial and temporal scales, resulting from highly variable vegetation characteristics [117] . The variability is mainly due to other components in the ecosystem, such as climate, soil condition, topography [105] [118] [119] , of which soil accounts for most variation at small scale whereas topography, landforms and climate explains for most variation at large scale [120] [121] .
In North American mixed prairie, disturbances from all these three aspects are interweaved into each other, even coupled with many other components (e.g. prairie dogs, bison wallows etc.), which makes it difficult to evaluate their impact and the mechanism. They work altogether and shape the biodiversity and productivity of the entire grassland ecosystem [122] . This makes it necessary to investigate those disturbances concurrently in the same context. For example, fire is less likely to occur and spread where the available fuel load is insufficient to support it, which is usually caused by disturbances of grazing and/or drought. Drought not only makes vegetation prone to wildfires, but also limits herbivory as well. Meanwhile burned patches on the grassland landscape are more likely to attract herbivores, henceforth promoting the local intensity of grazing [103] . However, there are few researches specifically focusing on all these interacting processes, mainly because the combination of all major disturbances makes it too complicated to design the field experiment or hypothesize a reliable simulation model.
In light of the challenges in understanding impacts from various disturbances in the ecosystem, one promising solution is building a comprehensive, effective and robust simulation framework by synthesizing our current understanding of the grasslands ecosystem and rich dataset from resourceful yet cost-effective remote sensed images. This can not only make scientific contribution in understanding the patterns, processes and mechanisms of the complex ecosystem under disturbances, but also serve as a decision support tool for relevant practitioners such as environmental agencies, range managers and insurance companies etc. Adler et al. [38] developed a Cellular Automata model with spatial simulation capability to explore the critical condition in which grazing whether increase or decrease spatial heterogeneity of the vegetation. Some other computer models have been developed to evaluate and predict vegetation changes in the landscape level [123] , including FETM, LANDSUM, SIMPPLLE and VDDT. VDDT in particular has been tested in the northern mixed prairie with encouraging results. The Vegetation Dynamics Development Tool [124] , VDDT, is a landscape modeling framework for examining the impacts on vegetation change from different disturbances as well as management strategies. It takes into account a variety of natural disturbances, management activities and plant dynamics: human activity, fires, mammals, weather, growth, competition, and others. It is designed to evaluate the complex interaction of various disturbances, especially their combined effects over long periods. VDDT is able to predict the change in land area for vegetation classes (canopy cover, LAI etc.), using disturbance probabilities and pathways data of vegetation. However it is a nonspatial model, since it does not have a GIS component to map vegetation changes geographically and to be aware of spatial context in dynamic simulations. VDDT was later improved into a spatially explicit model and called “Tool for Exploratory Landscape Scenario Analyses” or TELSA [125] [126] . VDDT was originally developed to simulate changes in vegetative composition and structure for Columbia River Basin [127] . And it has been used to evaluate fire regimes for all vegetation types throughout U.S. [128] . TELSA was used by Grasslands National Park (GNP) in Canada to simulate its grasslands ecosystem [129] , evaluating vegetation’s response to a variety of disturbances such as grazing, fire and exotic species invasion in the protected grasslands park area. They reported encouraging result from the modeling, showing the potential of simulation framework in understanding compound effect of various disturbances in the grassland ecosystem. Seeing this, the authors suggest that such framework lends itself well for implementing and synthesizing ecological theories on grassland study, and for testing various hypotheses and scenarios. Meanwhile appropriate field designs are expected to validate the disturbance theories within such simulation framework.
Acknowledgements
Funding for this research is provided by China Scholarship Council and Graduate Scholarship of University of Saskatoon. Much appreciation goes to the reviewers, without their valuable comments this work would never be published.
References
- Anderson, R.C. (2006) Evolution and Origin of the Central Grassland of North America: Climate, Fire, and Mammalian Grazers. The Journal of the Torrey Botanical Society, 133, 626-647. http://dx.doi.org/10.3159/1095-5674(2006)133[626:EAOOTC]2.0.CO;2
- Shantz, H.L. (1954) The Place of Grasslands in the Earth’s Cover of Vegetation. Ecology, 35, 143-145. http://dx.doi.org/10.2307/1931110
- Vitousek, P. and Hooper, D. (1993) Biological Diversity and Terrestrial Ecosystem Biogeochemistry. In: Biodiversity and Ecosystem Function, Springer, Berlin, 3-140.
- Sala, O., Chapin, F.I., Armesto, J., Berlow, E., Bloomfield, J., Dirzo, R., Huber-Sanwald, E., Huenneke, L., Jackson, R., Kinzig, A., Leemans, R., Lodge, D., Mooney, H., Oesterheld, M., Poff, N., Sykes, M., Walker, B., Walker, M. and Wall, D. (2000) Global Biodiversity Scenarios for the Year 2100. Science, 287, 1770-1774. http://dx.doi.org/10.1126/science.287.5459.1770
- Sala, O., van Vuuren, D. and Pereira, H. (2005) Biodiversity across Scenarios. Ecosystems and Human Well Being Scenario, Island Press, Washington DC, 375-408.
- Ford, P.L. and McPherson, G.R. (1996) Ecology of Fire in Shortgrass Prairie of the Southern Great Plains. In: Finch, D.M., Ed., Ecosystem Disturbance and Wildlife Conservation in Western Grasslands—A Symposium Proceedings. Diane Publishing Co., Darby, 20-39.
- White, P.S. (1979) Pattern, Process, and Natural Disturbance in Vegetation. The Botanical Review, 45, 229-299. http://dx.doi.org/10.1007/BF02860857
- Gibson, D.J. and Hulbert, L.C. (1987) Effects of Fire, Topography and Year-to-Year Climatic Variation on Species Composition in Tallgrass Prairie. Vegetatio, 72, 175-185
- Anderson, R.C. (1990) Fire in North American Tallgrass Prairies. University of Oklahoma Press, Norman, 8-18.
- Savage, C (2011) Priaire: A Natural History. 2nd Edition, Greystone Books/David Suzuki Foundation, Vancouver.
- Shay, J., Whelan-Enns, G., Appleby, A., Usher, R., Kavanagh, K., Sims, M., Chaplin, S., Sims, P., Carney, K. and Dinerstein, E. (2013) Northern Mixed Grasslands. http://worldwildlife.org/ecoregions/na0810
- Cunfer, G. (2005) On the Great Plains: Agriculture and Environment. Texas A&M University Press, College Station.
- Archibold, O.W. and Wilson, M.R. (1980) The Natural Vegetation of Saskatchewan Prior to Agricultural Settlement. Canadian Journal of Botany, 58, 2031-2042. http://dx.doi.org/10.1139/b80-236
- Savage, C. (2011) Prairie—A Natural History. D & M Publishers Inc., Vancouver.
- WWF Global (2009) Northern Great Plains. http://wwf.panda.org/?uProjectID=CA0059
- Collins, S.L. (1987) Interaction of Disturbances in Tallgrass Prairie: A Field Experiment. Ecology, 68, 1243-1250. http://dx.doi.org/10.2307/1939208
- Gibson, D.J. (1989) Effects of Animal Disturbance on Tallgrass Prairie Vegetation. American Midland Naturalist, 121, 144-154. http://dx.doi.org/10.2307/2425665
- Gibson, D.J. (2009) Grasses & Grassland Ecology. Oxford University Press Inc., New York.
- Kansas Natural Heritage Inventory (2007) Native Prairie Hay Meadows: A Landowner’s Management Guide. University of Kansas, Lawrence.
- Parton, W.J., Scurlock, J.M.O., Ojima, D.S., Schimel, D.S. and Hall, D.O. (1995) Impact of Climate Change on Grassland Production and Soil Carbon Worldwide. Global Change Biology, 1, 13-22
- Polis, G.A. (1999) Why Are Parts of the World Green? Multiple Factors Control Productivity and the Distribution of Biomes. Oikos, 86, 3-15. http://dx.doi.org/10.2307/3546565
- Hairston, N. (1960) Community Structure, Population Control and Competition. The American Naturalist, 94, 421-425. http://dx.doi.org/10.1086/282146
- White, P.S. and Pickett, S.T.A. (1985) Natural Disturbance and Patch Dynamics: An Introduction. In: The Ecology of Natural Disturbance and Patch Dynamics, Academic Press, Orlando, 3-16. http://dx.doi.org/10.1016/B978-0-08-050495-7.50006-5
- Collins, S.L. and Glenn, S.M. (1988) Disturbance and Community Structure in North American Prairies. In: Diversity and Pattern in Plant Communities, SPB Academic, The Hague, 131-143.
- Arnold, E. (1992) World Atlas of Desertification. UNEP, London.
- Briske, D.D., Fuhlendorf, S.D. and Smeins, F.E. (2003) Vegetation Dynamics on Rangelands—A Critique of the Current Paradigms. Journal of Applied Ecology, 40, 601-614. http://dx.doi.org/10.1046/j.1365-2664.2003.00837.x
- Dobak, W.A. (1996) Killing the Canadian Buffalo. The Western Historical Quarterly, 27, 33-52. http://dx.doi.org/10.2307/969920
- Bruynooghe, J. and Macdonald, R. (2008) Managing Saskatchewan Rangeland. Revised Edition, Agriculture and AgriFood Canada, Saskatoon.
- Detling, J.K. (1988) Grasslands and Savannas: Regulation of Energy Flow and Nutrient Cycling by Herbivores. Concepts of Ecosystem Ecology, 67, 131-148. http://dx.doi.org/10.1007/978-1-4612-3842-3_7
- McNaughton, S.J. (1986) On Plants and Herbivores. The American Naturalist, 128, 765-770. http://dx.doi.org/10.1086/284602
- Paige, K.N. and Whitham, T.G. (1987) Over Compensation in Response to Mammalian Herbivory: The Advantage of Being Eaten. The American Naturalist, 129, 407-416. http://dx.doi.org/10.1086/284645
- Boddey, R.M., Macedo, R., Tarre, R.M., de Oliveira, O.C. and Rezende, C.D. (2004) Nitrogen Cycling in Brachiaria Pastures: The Key to Understanding the Process of Pasture decline. Agriculture, Ecosystems & Environment, 103, 389- 403. http://dx.doi.org/10.1016/j.agee.2003.12.010
- McNaughton, S.J. (1979) Grazing as an Optimization Process: Grass-Ungulate Relationships in the Serengeti. The American Naturalist, 113, 691-703. http://dx.doi.org/10.1086/283426
- Patton, B.D., Dong, X.P., Nyren, E. and Nyren, A. (2007) Effects of Grazing Intensity, Precipitation and Temperature on Forage Production. Rangel Ecology Management, 60, 656-665. http://dx.doi.org/10.2111/07-008R2.1
- Belsky, A.J. (1986) Does Herbivory Benefit Plants? A Review of the Evidence. The American Naturalist, 127, 870-892. http://dx.doi.org/10.1086/284531
- Milchunas, D.G., Forwood, J.R. and Lauenroth, W.K. (1994) Productivity of Long-Term Grazing Treatments in Response to Seasonal Precipitation. Journal of Range Management, 47, 133-139. http://dx.doi.org/10.2307/4002821
- Collins, S.L. and Steinauer, E.M. (1998) Disturbance, Diversity, and Species Interactions in Tallgrass Prairie. In: Grassland Dynamics: Long-Term Ecological Research in Tallgrass Prairie, Oxford University Press, Inc., New York, 140-156.
- Adler, P.B., Raff, D.A. and Lauenroth, W.K. (2001) The Effect of Grazing on the Spatial Heterogeneity of Vegetation. Oecologia, 128, 465-479. http://dx.doi.org/10.1007/s004420100737
- Hobbs, R.J. and Huenneke, L.F. (1992) Disturbance, Diversity, and Invasion: Implications for Conservation. Conservation Biology, 6, 324-337. http://dx.doi.org/10.1046/j.1523-1739.1992.06030324.x
- McNaughton, S.J., Wallace, L.L. and Coughenour, M.B. (1983) Plant Adaptation in an Ecosystem Context: Effects of Defoliation, Nitrogen, and Water on Growth of an African C4 Sedge. Ecology, 64, 307-318. http://dx.doi.org/10.2307/1937078
- Olff, H. and Ritchie, M.E. (1998) Effects of Herbivores on Grassland Plant Diversity. Trends in Ecology & Evolution, 13, 261-265. http://dx.doi.org/10.1016/S0169-5347(98)01364-0
- Dyksterhuis, E.J. (1949) Condition and Management of Rangeland Based on Quantitative Ecology. Journal of Range Management, 41, 450-459.
- Dyer, M.I., Detling, J.K., Coleman, D.C. and Hilbert, D.W. (1982) The Role of Herbivores in Grasslands. In: Grasses and Grasslands, University of Oklahoma Press, Norman, 255-295.
- Milchunas, D.G. and Lauenroth, W.K. (1993) Quantitative Effects of Grazing on Vegetation and Soils over a Global Range of Environments. Ecological Monographs, 63, 327-366. http://dx.doi.org/10.2307/2937150
- Milchunas, D.G., Sala, O.E. and Lauenroth, W.K. (1988) A Generalized Model of the Effects of Grazing by Large Herbivores on Grassland Community Structure. The American Naturalist, 132, 87-106. http://dx.doi.org/10.1086/284839
- Bakker, C., Blair, J.M. and Knapp, A.K. (2003) Does Resource Availability, Resource Heterogeneity or Species Turnover Mediate Changes in Plant Species Richness in Grazed Grasslands? Oecologia, 137, 385-391. http://dx.doi.org/10.1007/s00442-003-1360-y
- Westoby, M., Walker, B. and Noy-meir, I. (1989) Opportunistic Management for Rangelands Not at Equilibrium. Journal of Range Management, 42, 266-274. http://dx.doi.org/10.2307/3899492
- Jamsranjav, C. (2009) Sustainable Rangeland Management in Mongolia : The Role of Herder Community Institutions. Reykjavik, Iceland, 1-36.
- Pyne, S.J. (2001) Fire: A Brief History. University of Washington Press, Seattle.
- Pyne, S.J. (1982) Fire in America: A Cultural History of Wildland and Rural Fire. Princeton University Press, Princeton.
- Barrett, S. and Arno, S. (1999) Indian Fires in the Northern Rockies. In: Indians, Fire L.—Pacific Northwest, Oregon State University Press, Corvallis, 50-62.
- Pyne, S.J. (2001) Fire: A Brief History. University of Washington Press, Seattle.
- Ford, P.L. and McPerson, G.R. (1998) Trillium cernuum, the Rediscovery of the Species and the Ecological Restoration of Its Surrounding Habitat. Proceedings of 15th North American Prairie Conference, Natural Areas Association, Bend, 71-76. econatres.napc15.pford.pdf
- Courtwright, J. (2007) “When We First Come Here It All Looked like Prairie Land Almost”: Prairie Fire and Plains Settlement. The Western Historical Quarterly, 38, 157-179.
- Chapman, H.H. (1936) Effects of Fire in Propagation of Seedbed for Longleaf Pine Seedlings. Journal of Forestry, 34, 852-854.
- Heyward, F. (1937) The Effect of Frequent Fires on Profile Development of Longleaf Pine Forest Soils. Journal of Forestry, 35, 23-27.
- Wright, H.A. and Bailey, A. (1982) Fire Ecology—United States and Southern Canada. John Wiley and Sons, New York.
- Sauer, C. (1950) Grassland Climax, Fire, and Man. Journal of Range Management, 3, 16-21. http://dx.doi.org/10.2307/3894702
- Bond, W.J. and Keeley, J.E. (2005) Fire as a Global “Herbivore”: The Ecology and Evolution of Flammable Ecosystems. Trends in Ecology & Evolution, 20, 387-394. http://dx.doi.org/10.1016/j.tree.2005.04.025
- Grover, H.D. and Musick, H.B. (1990) Shrubland Encroachment in Southern New Mexico, U.S.A.: An Analysis of Desertification Processes in the American Southwest. Climatic Change, 17, 305-330. http://dx.doi.org/10.1007/BF00138373
- Ford, P.L. and Johnson, G.V. (2006) Effects of Dormantvs. Growing-Season Fire in Shortgrass Steppe: Biological Soil Crust and Perennial Grass Responses. Journal of Arid Environments, 67, 1-14. http://dx.doi.org/10.1016/j.jaridenv.2006.01.020
- Dwyer, D.D. and Pieper, R.D. (1967) Fire Effects of Blue Grama-Pinyon-Juniper Rangeland in New Mexico. Journal of Range Management, 20, 359-362. http://dx.doi.org/10.2307/3896405
- Launchbaugh, J.L. (1964) Effects of Early Spring Burning on Yields of Native Vegetation. Journal of Range Management, 17, 5-6. http://dx.doi.org/10.2307/3895539
- Steuter, A.A. and McPherson, G.R. (1995) Fire as a Physical Stress. In: Bedunah, D.J. and Sosebee, R.E., Eds., Wildland Plants: Physiological Ecology and Developmental Morphology, Society for Range Management, Denver, 550- 579.
- Uys, R. (2004) The Effects of Different Fire Regimes on Plant Diversity in Southern African Grasslands. Biological Conservation, 118, 489-499. http://dx.doi.org/10.1016/j.biocon.2003.09.024
- Leach, M.K. and Givnish, T.J. (1996) Ecological Determinants of Species Loss in Remnant Prairies. Science, 273, 1555-1558. http://dx.doi.org/10.1126/science.273.5281.1555
- Peterson, D.W. and Reich, P.B. (2001) Prescribed Fire in Oak Savanna: Fire Frequency Effects on Stand Structure and Dynamics. Ecological Applications, 11, 914-927. http://dx.doi.org/10.1890/1051-0761(2001)011[0914:PFIOSF]2.0.CO;2
- Scasta, J.D., Engle, D.M., Talley, J.L., Weir, J.R., Stansberry, J.C., Fuhlendorf, S.D. and Harr, R.N. (2012) Pyric-Herbivory to Manage Horn Flies (Diptera: Muscidae) on Cattle. Southwestern Entomologist, 37, 325-334. http://dx.doi.org/10.3958/059.037.0308
- Anderson, K.L., Smith, E.F. and Owensby, C.E. (1970) Burning Bluestem Range. Journal of Range Management, 23, 81-92. http://dx.doi.org/10.2307/3896105
- Heirman, A.L. and Wright, H.A. (1973) Fire in Medium Fuels of West Texas. Journal of Range Management, 26, 331-335. http://dx.doi.org/10.2307/3896849
- White, R.S. and Currie, P.O. (1983) Prescribed Burning in the Northern Great Plains: Yield and Cover Responses of 3 Forage Species in the Mixed Grass Prairie. Journal of Range Management, 36, 179-183. http://dx.doi.org/10.2307/3898158
- Umbanhowar, C.E. (1996) Recent Fire History of the Northern Great Plains. American Midland Naturalist, 135, 115-121. http://dx.doi.org/10.2307/2426877
- Brockway, D.G., Gatewood, R.G. and Paris, R.B. (2002) Restoring Fire and an Ecological Process in Shortgrass Prairie Ecosystems: Initial Effects of Prescribed Burning during the Dormant and Growing Seasons. Journal of Environmental Management, 65, 135-152. http://dx.doi.org/10.1006/jema.2002.0540
- Guo, X., Price, K.P. and Stiles, J.M. (2000) Biophysical and Spectral Characteristics of Cooland Warm-Season Grasslands under Three Land Management Practices in Eastern Kansas. Natural Resources Research, 9, 321-331. http://dx.doi.org/10.1023/A:1011513527965
- Dubinin, M., Potapov, P., Lushchekina, A. and Radeloff, V.C. (2010) Reconstructing Long Time Series of Burned Areas in Arid Grasslands of Southern Russia by Satellite Remote Sensing. Remote Sensing of Environment, 114, 1638- 1648. http://dx.doi.org/10.1016/j.rse.2010.02.010
- Scholes, R.J. and Archer, S.R. (1997) Tree-Grass Interactions in Savannas. Annual Review of Ecology and Systematics, 28, 517-544. http://dx.doi.org/10.1146/annurev.ecolsys.28.1.517
- Lieth, H.F.H. (1978) Patterns of Primary Productivity in the Biosphere. Hutchinson Ross, Stroudsberg.
- Rosenzweig, M.L. (1968) Net Primary Productivity of Terrestrial Communities: Prediction from Climatological Data. The American Naturalist, 102, 67-74. http://dx.doi.org/10.1086/282523
- Whittaker, R.H. (1975) Communities and Ecosystems. 2nd Revise Edition, MacMillan Publishing Co., New York.
- Oesterheld, M., Loreti, J., Semmartin, M. and Paruelo, J.M. (1999) Grazing, Fire, and Climate Effects on Primary Productivity of Grasslands and Savanna. In: Walker, L.L., Ed., Ecosystems of Disturbed Grounds, Series Ecosystems of the World, Elsevier, New York, 287-306.
- Sherow, J.E. (1992) Workings of the Geodialectic: High Plains Indians and Their Horses in the Region of the Arkansas River Valley, 1800-1870. Environmental History Review, 16, 61-84.
- NOAA (2014) North American Drought—A Paleo Perspective (Paleoclimatology and Drought). National Climatic Data Center, 1. http://www.ncdc.noaa.gov/paleo/drought/drght_history.html
- Knapp, A.K. and Smith, M.D. (2001) Variation among Biomes in Temporal Dynamics of Aboveground Primary Production. Science, 291, 481-484. http://dx.doi.org/10.1126/science.291.5503.481
- Lauenroth, W.K. and Sala, O. (1992) Long-Term Forage Production of the North American Shortgrass Steppe. Ecological Applications, 2, 397-403. http://dx.doi.org/10.2307/1941874
- Warren, W., Stan, S., William, L., Russell, K. and Milton, S. (1978) Primary Productivity and Water Use in Native Forest, Grassland, and Desert Ecosystems. Ecology, 59, 1239-1247. http://dx.doi.org/10.2307/1938237
- Hopkins, H., Albertson, F.W. and Riegel, A. (1948) Some Effects of Burning upon a Prairie in West-Central Kansas. Transactions of the Kansas Academy of Science, 51, 131-141. http://dx.doi.org/10.2307/3625752
- Knapp, E.E., Estes, B.L. and Skinner, C.N. (2009) Ecological Effects of Prescribed Fire Season: A Literature Review and Synthesis for Managers. Department of Agriculture, Forest Service, Pacific Southwest Research Station, Albany.
- Agee, J.K. (1993) Fire Ecology of Pacific Northwest Forests. Island Press, Covelo.
- Brown, J.K. (2000) Introduction and Fire Regimes. In: Wildland Fire in Ecosystems: Effect of Fire on Flora, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden, 1-7.
- Van Langevelde, F., Van De Vijver, C.A.D.M., Kumar, L., Van De Koppel, J., De Ridder, N., Van Andel, J., Skidmore, A.K., Hearne, J.W., Stroosnijder, L., Bond, W.J., Prins, H.H.T. and Rietkerk, M. (2003) Effects of Fire and Herbivory on the Stability of Savanna Ecosystems. Ecology, 84, 337-350. http://dx.doi.org/10.1890/0012-9658(2003)084[0337:EOFAHO]2.0.CO;2
- Spasojevic, M.J., Aicher, R.J., Koch, G.R., Marquardt, E.S., Mirotchnick, N., Troxler, T.G. and Collins, S.L. (2010) Fire and Grazing in a Mesic Tallgrass Prairie: Impacts on Plant Species and Functional Traits. Ecology, 91, 1651-1659. http://dx.doi.org/10.1890/09-0431.1
- Hulbert, L.C. (1985) History and Use of Konza Prairie Research Natural Area. Prairie Scout, 5, 63-93.
- Ehleringer, J.R. and Monson, R.K. (1993) Evolutionary and Ecological Aspects of Photosynthetic Pathway Variation. Annual Review of Ecology, Evolution, and Systematics, 24, 411-439. http://dx.doi.org/10.1146/annurev.es.24.110193.002211
- Collins, S.L. and Smith, M.D. (2006) Scale-Dependent Interaction of Fire and Grazing on Community Heterogeneity in Tallgrass Prairie. Ecology, 87, 2058-2067. http://dx.doi.org/10.1890/0012-9658(2006)87[2058:SIOFAG]2.0.CO;2
- Fuhlendorf, S.D. and Engle, D.M. (2004) Application of the Fire-Grazing Interaction to Restore a Shifting Mosaic on Tallgrass Prairie. Journal of Applied Ecology, 41, 604-614. http://dx.doi.org/10.1111/j.0021-8901.2004.00937.x
- Whelan, R.J. (1995) The Ecology of Fire. Cambridge University Press, Cambridge.
- Keeley, J.E., Lubin, D. and Fotheringham, C.J. (2003) Fire and Grazing Impacts on Plant Diversity and Alien Plant Invasions in the Southern Sierra Nevada. Ecological Applications, 13, 1355-1374. http://dx.doi.org/10.1890/02-5002
- Trager, M.D., Wilson, G.W.T. and Hartnett, D.C. (2004) Concurrent Effects of Fire Regime, Grazing and Bison Wallowing on Tallgrass Prairie Vegetation. The American Midland Naturalist, 152, 237-247. http://dx.doi.org/10.1674/0003-0031(2004)152[0237:CEOFRG]2.0.CO;2
- Fuhlendorf, S.D., Engle, D.M., Kerby, J. and Hamilton, R. (2009) Pyric Herbivory: Rewilding Landscapes through the Recoupling of Fire and Grazing. Conservation Biology, 23, 588-598. http://dx.doi.org/10.1111/j.1523-1739.2008.01139.x
- Weir, J.R., Fuhlendorf, S.D., Engle, D.M., Bidwell, T.G., Cumminggs, D.C. and Elmore, D. (2007) Patch Burning: Integrating Fire and Grazing to Promote Heterogeneity. Oklahoma State University, Stillwater.
- Allred, B.W., Fuhlendorf, S.D., Engle, D.M. and Elmore, R.D. (2011) Ungulate Preference for Burned Patches Reveals Strength of Fire-Grazing Interaction. Ecology and Evolution, 1, 132-144. http://dx.doi.org/10.1002/ece3.12
- Kagima, B. and Eairbanks, W.S. (2013) Habitat Selection and Diet Composition of Reintroduced Native Ungulates in a Fire-Managed Tallgrass Prairie Reconstruction. Ecological Restoration, 21, 79-88.
- Archibald, S., Bond, W.J., Stock, W.D. and Fairbanks, D.H.K. (2005) Shaping the Landscape: Fire-Grazer Interactions in an African savanna. Ecological Applications, 15, 96-109. http://dx.doi.org/10.1890/03-5210
- McGranahan, D.A., Engle, D.M., Fuhlendorf, S.D., Winter, S.J., Miller, J.R. and Debinski, D.M. (2012) Spatial Heterogeneity across Five Rangelands Managed with Pyric-Herbivory. Journal of Applied Ecology, 49, 903-910. http://dx.doi.org/10.1111/j.1365-2664.2012.02168.x
- Yang, X. (2012) Assessing Responses of Grasslands to Grazing Management Using Remote Sensing Approaches. University of Saskatchewan, Saskatoon.
- Ellis, J.E. and Swift, D.M. (1988) Stability of African pastoral Ecosystems: Alternate Paradigms and Implications for Development. Journal of Range Management, 41, 450-459. http://dx.doi.org/10.2307/3899515
- Biondini, M.E. and Manske, L. (1996) Grazing Frequency and Ecosystem Processes in Northern Mixed Prairie, USA. Ecological Applications, 6, 239-256. http://dx.doi.org/10.2307/2269567
- Fuhlendorf, S.D., Briske, D.D. and Smeins, F.E. (2001) Herbaceous Vegetation Change in Variable Rangeland Environments: The Relative Contribution of Grazing and Climatic Variability. Applied Vegetation Science, 4, 177-188. http://dx.doi.org/10.1111/j.1654-109X.2001.tb00486.x
- Gill, A.M. (1975) Fire and the Australian Flora: A Review. Australian Forestry, 38, 4-25. http://dx.doi.org/10.1080/00049158.1975.10675618
- Rothermal, R.C. (1983) How to Predict the Spread and Intensity of Forest and Range Fires. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station, Ogden.
- Andrews, P L (1986) BEHAVE: Fire Behavior Prediction and Fuel Modeling System/Burn Subsystem, Part I. General Technical Report INT-GTR-194, USDA Forest Service, Intermountain Research Station, Ogden.
- Briske, D.D. (1991) Developmental Morphology and Physiology of Grasses. In: Grazing Management: An Ecological Perspective, Timber Press, Portland, 85-108.
- Evans, E.W., Briggs, J.M., Finck, E.J., Gibson, D.J. and James, S.W. (1988) Is Fire a Disturbance in Grasslands? Proceedings of the North American Prairie Conferences, Lincoln, 7-11 August 1988, 159-161.
- Chaneton, E.J., Perelman, S.B., Omacini, M. and Le, R.J.C. (2002) Grazing, Environmental Heterogeneity, and Alien Plant Invasions in Temperate Pampa grasslands. Biological Invasions, 4, 7-24. http://dx.doi.org/10.1023/A:1020536728448
- Grimm, V. and Railsback, F. (2005) Individual-Based Modeling and Ecology. Princeton University Press, Princeton.
- Tansley, A.G. (1935) The Use and Abuse of Vegetational Concepts and Terms. Ecology, 16, 284-307. http://dx.doi.org/10.2307/1930070
- Ludwig, J.A. and Tongway, D.J. (1995) Spatial Organisation of Landscapes and Its Function in Semi-Arid Woodlands, Australia. Landscape Ecology, 10, 51-63. http://dx.doi.org/10.1007/BF00158553
- Urban, D.L., O’Neill, R.V. and Shugart, H.H.J. (1987) Ecology Landscape Spatial Patterns. BioScience, 37, 119-127. http://dx.doi.org/10.2307/1310366
- He, Y., Guo, X. and Wilmshurst, J. (2006) Studying Mixed Grassland Ecosystems I: Suitable Hyperspectral Vegetation indices. Canadian Journal of Remote Sensing, 32, 98-107. http://dx.doi.org/10.5589/m06-009
- Reynolds, H.L., Hungate, B.A., Chapin, F.S. and D’Antonio, C.M. (1997) Soil Heterogeneity and Plant Competition in an Annual Grassland. Ecology, 78, 2076-2090.
- Sebastian, M.T. (2004) Role of Topography and Soils in Grassland Structuring at the Landscape and Community Scales. Basic and Applied Ecology, 5, 331-346. http://dx.doi.org/10.1016/j.baae.2003.10.001
- Collins, S.L. and Barber, S.C. (1985) Effects of Disturbance on Diversity in Mixed-Grass Prairie. Vegetatio, 64, 87-94. http://dx.doi.org/10.1007/BF00044784
- Barrett, T.M. (2001) Models of Vegetative Change for Landscape Planning: A Comparison of FETM, LANDSUM, SIMPPLLE, and VDDT. General Technical Report, U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Ogden. http://www.fs.fed.us/rm/pubs/rmrs_gtr076.pdf
- ESSA Technologies Ltd. (2013) Vegetation Dynamics Development Tool. ESSA Homepage, 1. http://essa.com/tools/vddt/
- Kurz, W.A., Beukema, S.J., Klenner, W., Greenough, J.A., Robinson, D.C.E., Sharpe, A.D. and Webb, T.M. (2000) TELSA: The Tool for Exploratory Landscape Scenario Analyses. Computers and Electronics in Agriculture, 27, 227- 242.
- ESSA Technologies Ltd. (2008) TELSA : Tool for Exploratory Landscape Scenario Analyses Model Description, Version 3, 6. Vancouver. http://www.essa.com/media/ModelDescription.pdf
- Hemstrom, M.A., Korol, J.J. and Hann, W.J. (2001) Trends in Terrestrial Plant Communities and Landscape Health Indicate the Effects of Alternative Management Strategies in the Interior Columbia River Basin. Forest Ecology and Management, 5504, 1-21.
- LANDFIRE Helpdesk (2013) LANDFIRE. LANDFIRE Homepage, 1. http://www.landfire.gov/index.php
NOTES

*Corresponding author.