 Journal of Modern Physics, 2011, 2, 379-391 doi:10.4236/jmp.2011.25047 Published Online May 2011 (http://www.SciRP.org/journal/jmp) Copyright © 2011 SciRes. JMP Thermodynamic Formulation of Living Systems and Their Evolution Luis Felipe del Castillo, Paula Vera-Cruz Departamento de Polímeros, Instituto de Investigaciones en Materiales, Universidad Nacional Autónoma de México, Mexico, Mexico E-mail: lfelipe@servidor.unam.mx Received January 20, 2011; revised March 7, 2011; accepted April 2, 2011 Abstract The purpose of this review article is to present some of the recent contributions that show the use of thermo- dynamics to describe biological systems and their evolution, illustrating the agreement that this theory pre- sents with the field of evolution. Organic systems are described as thermodynamic systems where entropy is produced by the irreversible processes, considering as an established fact that this entropy is eliminated through their frontiers to preserve life. The necessary and sufficient conditions to describe the evolution of life in the negentropy principle are established. Underlining the fact that the necessary condition requires formulation, which is founded on the principle of minimum entropy production for open systems operating near or far from equilibrium, other formulations are mentioned, particularly the information theory, the en- ergy intensiveness hypothesis and the theory of open systems far from equilibrium. Finally suggesting the possibility of considering the lineal formulation as a viable alternative; that is, given the internal constric- tions under which a biological system operates, it is possible that the validity of its application is broader than it has been suggested. Keywords: Entropy, Life, Evolution, Dissipative Structures, Negentropy Principle 1. Introduction The application of thermodynamics [1] to biological systems and their evolution has offered important con- tributions [2] by describing the characteristics of animal evolution [3]. The basic aspect underlined is that the laws of thermodynamics can predict the feasibility of the processes and the relation between the variables [4]. Schrödinger established the first contributions to the field in 1944 [5], offering some ideas about the concept of negentropy or negative entropy to describe the com- mon fact that physiological processes gradually generate an increase of the internal “order” in living organisms. In the sense that the action of living matter opposes the de- gradation of the organic constituents by effect of the ir- reversibilities [6,7]. Currently, the concept of order in relation to negen- tropy is ignored since a quantitative correlation has not been found. Alternatively, in biology the equivalence has been proposed with the concept of “organization” (re- garding structure), originated in information theory as a more appropriate way to quantify the degree of structur- ing (or contained information) of an organic system, as proposed by Brillouin in 1951 [8,9]. Schrödinger also applied the concept of negentropy to describe the evolution of species, which has received much attention [10]. Another related development can be attributed to Prigogine in 1946 [11], who applied the theory of linear non-equilibrium thermodynamics to de- scribe the phenomenon of adaptation of species. He con- sidered the organism as an open system in stationary state [7], which evolves in the direction that entropy production decreases, and reaches a minimum when the adaptation to the environment has concluded. With these first attempts, the phenomenon of adaptation was ex- pressed in a thermodynamic language, involving both chance and necessity [12], thus being characterized as a teleonomic [13] phenomenon. The present work describes the basic facts originated by the irreversibility in biological systems, and the rele- vant advances in animal evolution in terms of structure growth [14] and the introduction of stationary conditions (maturation process) [15]. Particularly, these two aspects are formulated in the negentropy principle, which estab-  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 380 lishes the necessary and sufficient conditions for the sur- vival of a species when the environment has suffered a radical change that endangers its existence. The suffi- cient condition is negentropic growth and the necessary condition observes that the adaptation process (maturity) is the search and achievement of the stationary state in the habitat surrounding a species. Afterward, the contributions to formulate the neces- sary condition for survival are discussed [11]. Particu- larly, the information theory, the energy intensiveness hypothesis, and the theory of self-organization of dissi- pative structures far from equilibrium. 2. Nature of Living Matter as a Thermodynamic System Important efforts have been made to describe biological systems from the point of view of macroscopic sciences, particularly thermodynamics and statistical physics [16,17]. Biological systems are organized internally by con- strictions. These are internal walls (membranes, epithelia, endothelia, interfaces, etc) and their role is to maintain separated two localities, each of them in equilibrium and specified by different local values of the thermodynamic variables; particularly, temperature, pressure or electro- chemical potentials. Generalized flows occur through these constrictions, such as transport of mass, charge, calorific energy and momentum [18]. Since the constric- tions hold the pressure differences, these flows occur under conditions of mechanical equilibrium, which is manifested by the absence of accelerations in them [19]. The present description considers the flows and forces at the local level, such as two entities on both sides of a cellular membrane (see Table 1). This description is known as mesoscopic (at the micron scale) and is gov- erned by the paradigm of mesoscopic systems, charac- terized by the presence of fluctuations formally described by the theory of Brownian motion in systems under elec- tromechanical equilibrium [20]. Assuming that equilibrium thermodynamics condi- tions prevail around the constrictions, such as cellular Table 1. Physiological processes are specified as flows pro- duced by forces. Generalized Flows (Jj) Forces (Xj) Diffusive passive 12 Diffusive active Coupling forces Volumetric movement of fluids 12 PPP Ion transport 12 Advance of chemical reaction Chemical affinity membranes, then the principle of regression of fluctua- tions is valid and the system is stable and behaves ac- cording to the Le Chatelier-Braun principle [21]. Equilibrium thermodynamics prescribes the law of en- tropy growth when generalized flows [22] occur through a membrane from locality 1 to locality 2. 12 ddd 0SS S (1) The differential expression for specific entropy is giv- en by: 11 1 ddd d j c j P Suv n TT T (2) In (2) u, v and nj are the internal energy, volume and number of particles per unit of mass, respectively, asso- ciated to a locality with c different species. Considering the presence of two localities separated by a membrane, the change of total specific entropy is given by the fol- lowing equation: 12 11 12 12 12 1 1 12 11 ddd d jj cj j PP Su v TT TT n TT (3) where the laws of conservation of energy, conservation of volume and conservation of mass have been applied. 12 dd d0uu u (4) 12 dd d0vv v (5) 12 dd d0nn n (6) Equalities (4-6) indicate that the system is operating under conditions of isolation (0U, where Uis the total internal energy). According to classical thermodynamics, the contribu- tion to the variation of entropy of each of the terms in (3) must be positive, as indicated by the Second Law of Thermodynamics. 11 12 11 dd0Su TT (7) 12 11 12 dd0 PP Sv TT (8) 12 11 1 12 dd0 jj cj j Sn TT (9) The validity of conditions (7-9) requires no additional or external actions influencing the flows or forces de- scribed, Therefore, the effects expressed in these equa- tions are considered independent (system regarded as iso- lated). 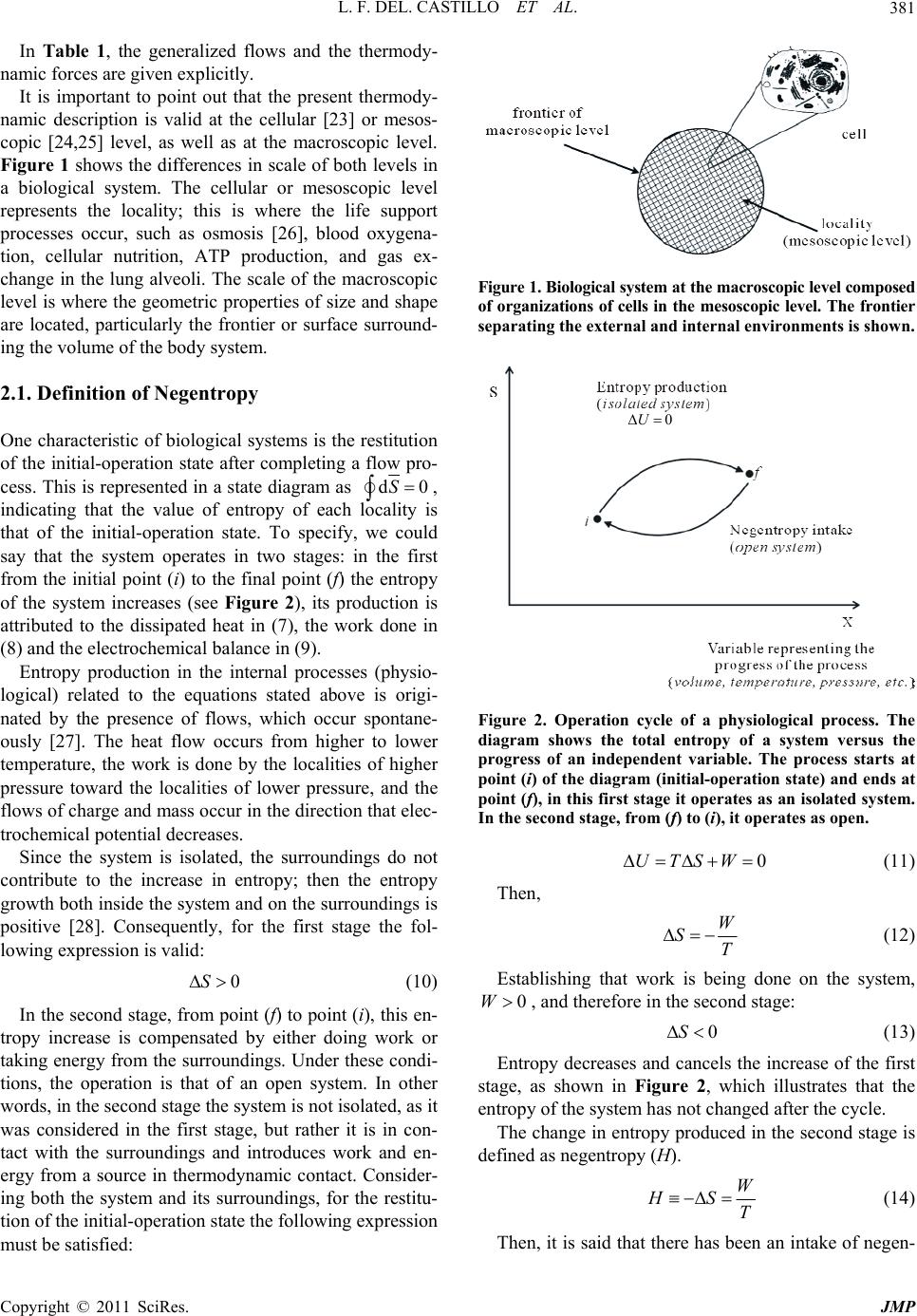 L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 381 In Table 1, the generalized flows and the thermody- namic forces are given explicitly. It is important to point out that the present thermody- namic description is valid at the cellular [23] or mesos- copic [24,25] level, as well as at the macroscopic level. Figure 1 shows the differences in scale of both levels in a biological system. The cellular or mesoscopic level represents the locality; this is where the life support processes occur, such as osmosis [26], blood oxygena- tion, cellular nutrition, ATP production, and gas ex- change in the lung alveoli. The scale of the macroscopic level is where the geometric properties of size and shape are located, particularly the frontier or surface surround- ing the volume of the body system. 2.1. Definition of Negentropy One characteristic of biological systems is the restitution of the initial-operation state after completing a flow pro- cess. This is represented in a state diagram as d0S , indicating that the value of entropy of each locality is that of the initial-operation state. To specify, we could say that the system operates in two stages: in the first from the initial point (i) to the final point (f) the entropy of the system increases (see Figure 2), its production is attributed to the dissipated heat in (7), the work done in (8) and the electrochemical balance in (9). Entropy production in the internal processes (physio- logical) related to the equations stated above is origi- nated by the presence of flows, which occur spontane- ously [27]. The heat flow occurs from higher to lower temperature, the work is done by the localities of higher pressure toward the localities of lower pressure, and the flows of charge and mass occur in the direction that elec- trochemical potential decreases. Since the system is isolated, the surroundings do not contribute to the increase in entropy; then the entropy growth both inside the system and on the surroundings is positive [28]. Consequently, for the first stage the fol- lowing expression is valid: 0S (10) In the second stage, from point (f) to point (i), this en- tropy increase is compensated by either doing work or taking energy from the surroundings. Under these condi- tions, the operation is that of an open system. In other words, in the second stage the system is not isolated, as it was considered in the first stage, but rather it is in con- tact with the surroundings and introduces work and en- ergy from a source in thermodynamic contact. Consider- ing both the system and its surroundings, for the restitu- tion of the initial-operation state the following expression must be satisfied: Figure 1. Biological sy stem at t he macrosc opic level compo sed of organizations of cells in the mesoscopic level. The frontier separating the external and internal environments is shown. 0U Figure 2. Operation cycle of a physiological process. The diagram shows the total entropy of a system versus the progress of an independent variable. The process starts at point (i) of the diagram (initial-operation state) and ends at point (f), in this first stage it operates as an isolated system. In the second stage, from (f) to (i), it operates as open. 0UTSW (11) Then, W ST (12) Establishing that work is being done on the system, 0W, and therefore in the second stage: 0S (13) Entropy decreases and cancels the increase of the first stage, as shown in Figure 2, which illustrates that the entropy of the system has not changed after the cycle. The change in entropy produced in the second stage is defined as negentropy (H). W HS T (14) Then, it is said that there has been an intake of negen-  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 382 tropy to the system [29]. On the other hand, a structure growth could have oc- curred in the system, which can also be described by (10) and (14) establishing two stages, as mentioned above. Then, the first stage features the transformation of food proteins and the ligands produced in the cell, where en- tropy increases. In the second stage, the structuring of the produced components involves work (W). Therefore, there are two types of negentropy intake: one occurs in the common physiological processes (H1) in the irre- versible flows, and the other in structure growth (H2). Thus, the total negentropy intake will be: 12 HH (15) 2.2. Dissipative Structures From the macroscopic point of view, the biological sys- tem behaves essentially as a dissipative structure, but the heterogeneity of its internal structure differentiates it from the physicochemical systems capable of becoming dissipative structures. For example, a metallic rod con- nected to two electric poles, kept under isothermal con- ditions, dissipates to the surroundings an amount of en- ergy defined by the product of the voltage applied and the intensity of the electrical current. At this level, the internal structure of the metal has a microscopic arran- gement at angstrom-level, much smaller than the scale of the mesoscopic structure. Another characteristic that se- parates the dissipative structures of the biological sys- tems from those of the physicochemical systems [30] is the ability to grow with time and reproduce itself without losing this characteristic. For example, the dissipative structures of Marangoni and Bénard [31,32] operate without these attributes. However, the biological dissipa- tive structures [33] can be described by physicochemical methods, as discussed below. The cellular formulation for entropy production at the local level is established according to a balance equation: d dSS S t (16) Equation (16) establishes that the gain or loss of en- tropy per unit volume is equal to the entropy produced minus the net entropy flow through the surfaces of the localities. Here, Sjjj X is the entropy produc- tion given by the sum of the tensor products of flows (Jj) and forces (Xj) present in the localities. The second term of (16) is the divergence of the flow of entropy that cor- responds to its net flow through the cellular surface. Alternatively, considering the macroscopic formula- tion of the biological system at the global level, by inte- gration of (16) we find the following expression for the production and flow of entropy: ddd ddd ie T SSS ttt (17) where, dd d dd T V SSv tt (18) In (18) V is the volume of the biological system. dd d e S S t e (19) where e is a unit vector pointing in the direction of the normal of every point of the frontier, indicated by the surface Σ. dd d i S V Sv t (20) Considering entropy production for a biological dissi- pative structure in the global formulation, from Table 2 we have that: dd dd ie SS tt (21) Equation (21) is the mathematical expression of the negentropy principle in the ontogenic scale of a proto- type. It indicates the requirement for negentropic growth: that the entropy produced in the system by the internal Table 2. Some cases where Equations 17-20 are applied according to the type of system considered. Type of systemConditions Relations of S (cellular for- mulation) Relations of entropy S (global for- mulation) Thermodynamic system Isolated (strict validity of the second law of thermodynam- ics) 0 S J d0 dS S t 0 S reversible process d0 d eS t dd 0 dd i T SS tt Physicochemical dissipative structure Open system with elimination of produced entropy d0 d S t 0 SS J 0 S reversible proc- ess dd dd ie SS tt Biological dissipative structure Open system with elimination of entropy and structure growth d0 d S t SS dd dd ie SS tt 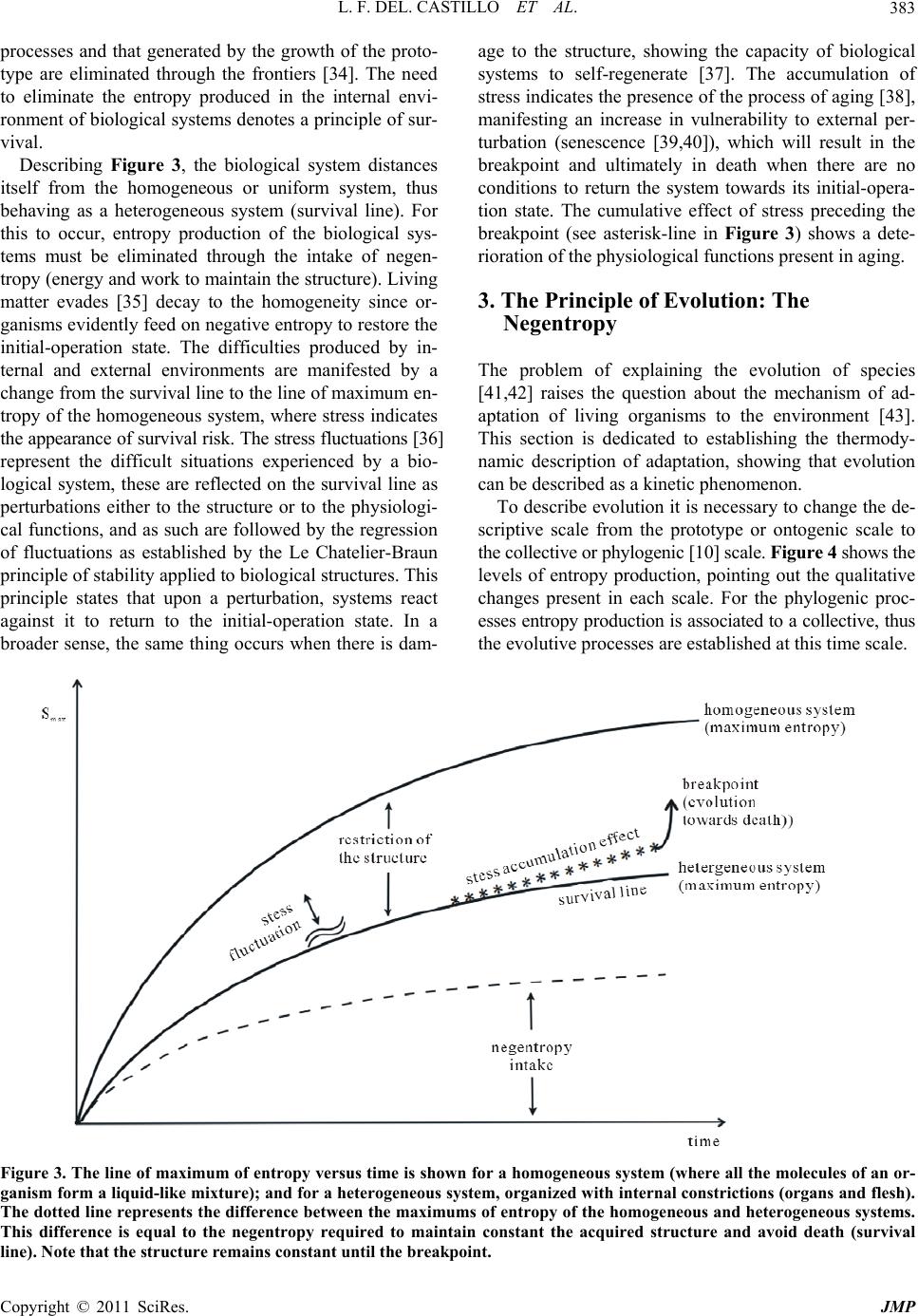 L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 383 processes and that generated by the growth of the proto- type are eliminated through the frontiers [34]. The need to eliminate the entropy produced in the internal envi- ronment of biological systems denotes a principle of sur- vival. Describing Figure 3, the biological system distances itself from the homogeneous or uniform system, thus behaving as a heterogeneous system (survival line). For this to occur, entropy production of the biological sys- tems must be eliminated through the intake of negen- tropy (energy and work to maintain the structure). Living matter evades [35] decay to the homogeneity since or- ganisms evidently feed on negative entropy to restore the initial-operation state. The difficulties produced by in- ternal and external environments are manifested by a change from the survival line to the line of maximum en- tropy of the homogeneous system, where stress indicates the appearance of survival risk. The stress fluctuations [36] represent the difficult situations experienced by a bio- logical system, these are reflected on the survival line as perturbations either to the structure or to the physiologi- cal functions, and as such are followed by the regression of fluctuations as established by the Le Chatelier-Braun principle of stability applied to biological structures. This principle states that upon a perturbation, systems react against it to return to the initial-operation state. In a broader sense, the same thing occurs when there is dam- age to the structure, showing the capacity of biological systems to self-regenerate [37]. The accumulation of stress indicates the presence of the process of aging [38], manifesting an increase in vulnerability to external per- turbation (senescence [39,40]), which will result in the breakpoint and ultimately in death when there are no conditions to return the system towards its initial-opera- tion state. The cumulative effect of stress preceding the breakpoint (see asterisk-line in Figure 3) shows a dete- rioration of the physiological functions present in aging. 3. The Principle of Evolution: The Negentropy The problem of explaining the evolution of species [41,42] raises the question about the mechanism of ad- aptation of living organisms to the environment [43]. This section is dedicated to establishing the thermody- namic description of adaptation, showing that evolution can be described as a kinetic phenomenon. To describe evolution it is necessary to change the de- scriptive scale from the prototype or ontogenic scale to the collective or phylogenic [10] scale. Figure 4 shows the levels of entropy production, pointing out the qualitative changes present in each scale. For the phylogenic proc- esses entropy production is associated to a collective, thus the evolutive processes are established at this time scale. Figure 3. The line of maximum of entropy versus time is shown for a homogeneous system (where all the molecules of an or- ganism form a liquid-like mixture); and for a heterogeneous syste m, organized with internal constrictions (organs and flesh). The dotted line represents the difference between the maximums of entropy of the homogeneous and heterogeneous systems. This difference is equal to the negentropy required to maintain constant the acquired structure and avoid death (survival line). Note that the structure remains constant until the breakpoint.  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 384 Figure 4. The different scales of time and entropy produc- tion in different systems, from the mesoscopic or celular scale to the phylogenic scale. Thermodynamically, the evolution of living matter is described by postulating the validity of the negentropy principle [44], which establishes that the evolution of bio- logical systems occurs in the direction [45] that the structure [46] becomes more complex. That is, the phy- logenic changes occur in the direction that the negen- tropy of the structure increases. However, this postulate represents the sufficient con- dition, since it only shows the need to evolve by gaining structuring; moreover, the adaptation must be established simultaneously, when the system has adjusted to the en- vironment in a stable condition. The negentropy principle can be associated with the inherent difficulty that the growth of the system escalates in size [47] and shape [48-50], and particularly it is found that entropy production increases with volume [51,52,3] (V), as shown by the expression for internal entropy production dd iSt according to (20). In con- trast, the elimination of the entropy of the system dd eSt depends on the area of the frontier (Σ), as seen in (19). Finally, the evolutionary structuring of the sys- tem does not occur in accordance with the volume growth of the species, but rather a balance between size and shape must be maintained for an efficient elimina- tion of produced entropy. Then, it is clear that growth is governed by thermodynamics. Particularly, the rate of entropy elimination with increasing body mass obeys the power law (with 0.75 instead of 1). Figure 5 shows how evolution has made more efficient the negentropy intake and entropy elimination without increasing the surface of the system [53]. One possible formulation of the necessary condition for survival is the adaptation of species to the environment, so as to minimize entropy production [54,55]. This can be established by the condition that the system operates Figure 5. How evolution has made more efficient the negen- tropy intake and entropy elimination without increasing the surface of the system. under stationary conditions. According to the require- ments of thermodynamics, for the fulfillment of this principle it is necessary that, at least for a given period of time, the action of a species be specifically coupled to its ecological surroundings [56-62] in a way that its relation with the environment does not change during this period of time. With this, efficiency is achieved in both energy consumption and entropy elimination: the metabolism reaches maximum efficiency. In the evolution of the species two parallel and inde- pendent events occur, as illustrated in Figure 6. These processes are the growth of the organic structure and the adaptation to the environment. The evolution pathway [65] is guided by natural selec- tion through the active mechanisms of heritage between generations in k steps [66-68]. In Figure 6, the arrows indicate the step from one generation to the next. This process leads to an increase in the demographic survival capacities [17]. It has been established that the evolutive process implies many generations and many events of Figure 6. The negentropy principle in terms of an evolution pathway. The adaptation process is established in k steps, where 1k to produce a stationary state between the prototypes of a species and the environment. The dashed line with white circles shows the separation between the species and the environment that produced the stress [63,64]; for the k generation the stress becomes cero.  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 385 combination and recombination of DNA between mem- bers of a large group. Then, the groups that failed to evolve in k steps, in accordance with the negentropy principle, have disappeared from the environment. In this sense, natural selection relates the two aspects: structure growth and the course of adaptation. Regarding the sufficient condition of the negentropy principle, the evolution appears to be continuous. How- ever, from the molecular point of view [69], it is a se- quence of jumps occurring stochastically. Concerning the formulation of the necessary condition of the negentropy principle, it has been established in two ways: firstly by using the principle of minimum en- tropy of linear non-equilibrium thermodynamics; and se- condly by the self-organization of dissipative structures far from equilibrium. This last point will be discussed in the next section. In relation to the principle of minimum entropy of non-equilibrium thermodynamics, it shows the require- ments for the transport coefficients to be constant and the flow-force relations to be linear. The first requirement is fulfilled by the condition that during a long enough pe- riod of time the physiological capabilities acquired do not change significantly. On the other hand, the linearity is satisfied by the requirement that evolution has mini- mized the gradients under which life operates. This is evidenced by two facts of the operation of phy- siological processes: first that they occur near electro- mechanical equilibrium; and second that they occur in nearly isothermal conditions in the internal environment of the system without taking into account the variations of the external environment, since they are controlled at the frontier (composed of the skin and its coat), as shown in Figure 7. 4. Parallel Formula tions The negentropy principle formulated in the previous T 0 S Figure 7. Production of entropy occurs internally and the entropy flow occurs at the frontier. In average, the station- ary state condition prevails internally. section describes the evolution of life as a dissipative structure regulated by entropy production, its elimination from the system [70], and the adaptation to the environ- ment. Parallel to this formulation, other alternatives pre- senting basic relations with thermodynamics have been established, such as information theory [71,72], the en- ergetic formulation of biological structures, and the the- ory of self-organization of complex systems far from equilibrium. These are not contrasting formulations but rather complementary aspects, since they highlight the kinetic descriptions that thermodynamics does not spec- ify within its own context, which are important aspects to explain the evolution of species. They are described be- low. 4.1. Information Theory According to information theory, entropy is defined by the following summation: 1ln j j Spp (22) where j indicates the number of each element involved in the information set representing the system, with the probability of occurrence pj. The factor is the total number of elements or complexions [8,9] in the set. The difficulty of applying this theory to biological systems lays in having to fully describe the details of the system within a numbered information set [73,74], since gener- ally is very large. The information is a quantity related to a physical state [75]. A similar difficulty is found in statistical physics when defining entropy, which is overcome by using an algo- rithm that counts the number of microstates that are ac- cessible to the system, thanks to the simplifying hy- pothesis that all the elements in the representative set have an equal probability [76]. In that case, if 1 j p then lnS (note that the entropy is a nondimen- sional quantity) [77]. For biological systems all probabilities are not neces- sarily equal, in fact they could even change with time since these systems operate irreversibly. It is thanks to modeling that the information contents of a biological system have been described, providing an insight to the characteristics of the evolution of life [78]. Recent advances in this direction highlight the achievements using evolutionary models that combine mutations and mechanisms of speciation [79,80]. Some bibliographic sources have referred to the feedback be- tween the prototype and the environment as cybernetic [81] aspect of evolutionary mechanisms. Significant progresses have occurred in the analysis of biological entities of small dimensions, as is the case of DNA [82-85] and other cell components, down to the  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 386 description of the cell itself [23,86]. Information theory gives a partial quantitative evalua- tion of the degree of organization [87], preserving the meaning of entropy according to statistical physics [88]. In fact, the increase of S corresponds to a loss of uncer- tainty; the approach dealt here indicates the lack of in- formation. The same occurs with thermodynamic theory, where irreversibility, counted as ΔS, is also associated with the degradation of the useful energy to restore the initial-operation state after spontaneous processes occur, which can be interpreted as a loss of information. Along these lines, based on information theory, the interpreta- tion of entropy-increase is linked to the increase of dis- organization for biological systems. Therefore, the ne- gentropy principle is reflected as an increase in the en- abled organization and system functions, indicating the presence of greater organization. Consequently, evolution is directed (teleonomy [13]) in the direction that organization increases. In this sense, the adaptation to an environment requires an increase in information and a greater efficiency in handling it (that is the meaning of the negentropy [89]). However, the connection between the concepts of en- tropy in information theory and thermodynamics is lack- ing. To achieve a quantitative comparison between these concepts, it is necessary to relate the probabilities used as measures of certainty with the thermodynamic variables of internal energy and the parameters that specify the constriction of the system. What stands out successfully in the theory of evolution is the relationship between organization and negentropy [90,7]. 4.2. Energy Intensiveness Hypothesis The intensiveness of energy consumption of a species can provide an evolutionary pattern as selection criteria in the evolution of species [91,92]. This relates to bio- logical evolution by considering the interactions between species of the same environment, particularly the ability to find, consume and guard resources as the dominant aspect of such evolution. According to this hypothesis it is the capacity to process energy (escalation), which has been a requirement given the critical situations that some species have faced [93]. Rigorously speaking, the negen- tropy principle is not contradicted, since the processing of energy and its conversion to work, as well as the ca- pacity to discard part of the energy, is the way to con- ceive a biotic machine [23] that can operate under condi- tions of irreversibility, thus satisfying this principle. The relation between the energy intensiveness approach and negentropic growth becomes clear from the fact that biological systems obtain energy from the exterior, such as the sun, and store the available energy inside their struc- ture as negentropy [94]. From this point of view, biodi- versity multiplies modes of energy consumption [95], and life evolution manifests itself by increased complexity [96], energy-gathering metabolic systems [97], teleonomic [13] character, as well as its abundance and diversity. 4.3. Self-Organization of Dissipative Structures According to the nonlinear hydrodynamic stability the- ory [98-101] and the description of coupled chemical reactions by chemical kinetics [102-104], dissipative structures as open systems outside equilibrium are capa- ble of self-organization and forming structural patterns [105,106] depending on the boundary conditions im- posed on the system [107]. The hypothesis that the same occurs for biological systems with a higher level of complexity has helped to establish new theoretical developments to explain both the origin of life as well as the diversity of species. Re- ferred to as chaos theory [108,109], it describes dissipa- tive structures capable of transforming themselves through a mechanism of time-space symmetry breaking. The new structures emerge in a point called bifurcation, after critical fluctuations, where several branches or pos- sibilities of pattern formation appear (see Figure 8). Natural selection could choose one of the branches in- fluenced by the environment (adaptation requirement) or simply any possibility of change could present itself (probabilistic requirement for diversity). The continuous transition towards a specific branch is possible, called “transitional bifurcation” [110,111]. This is also a result of self-organization at the edge of chaos [112]. In addition, some systems may show self-organi- zation of the first or second kind depending on boundary Figure 8. The possibilities of evolution of a system near equilibrium affected by an environmental crisis, it goes through long fluctuations and a bifurcation is produced to reach a new evolutionary branch to overcome the need for adaptation.  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 387 and initial conditions [113], they can also exhibit the development of complex hierarchical structures [114,115]. So far, the basic postulation of negentropic growth that negentropy intake is the cause for structure growth is still valid. However, kinetic formulations have sought to re- place the minimum dissipation principle for a broader one that explains not only adaptation but also the causes of evolution. 5. Discussion The thermodynamic description of the vital processes and their evolution is quantitative for the physiological processes in the mesoscopic scale, the global scale for a prototype, but qualitative in the phylogenic scale of the species. In the mesoscopic scale an endless row of phy- siological processes occur that in the ontogenic or global scale appear as a continuum. Then, it makes sense to talk about the entropy associated with a heterogeneous bio- logical system with its maximum compatible with the internal constraints, which are represented by the internal energy barriers established in the organization of the system. This entropy is lower than that which the same molecules would have if they were arranged homogene- ously in a liquid-like mixture. The difference between both amounts is the negentropy associated to the organic structure; it is interpreted as the amount of work or heat energy required to restore the initial-operation state. At the same time, to maintain the organic structure invariant, the entropy produced in the physiological processes when they reproduce the irreversible thermodynamic flows must be eliminated to avoid producing stress and risking life. This is how survival and proximity to death (breakpoint) are defined. On these bases, the thermodynamic image of life is that of a system that develops a cycle, with entropy pro- duction on one part and elimination on the other closing the cycle. Emphasizing the process of elimination, or- ganic systems are regarded as dissipative structures highly organized both mesoscopically and globally. The previous image is centered on the ontogenic scale. Re- garding the phylogenic, the time scale widens to contain biological changes in a time period much longer than the duration of a generation of prototypes of a species. In the preceding thermodynamic terms, the negentropy princi- ple is established with two requirements: an increase in organic structuring over time, and the establishment of a stationary state of high metabolic efficiency (minimum production of wasted energy). The determining factor of the negentropy principle is that one cannot be achieved without the other (necessary and sufficient conditions). The second requirement is set in a state of minimum en- tropy production in relation to a stationary one, thus gu- aranteeing the property of the regression of fluctuations, ensuring the survival of the species. The negentropy principle has been considered ade- quate for describing the general aspects of living matter and its evolution. At present, this principle is established as a biological law due to its general validity. The mechanisms proposed in the linear non-equilibrium thermodynamic description of adapta- tion to an environment through the establishment of a minimum dissipation stationary state are a good proposal. So much so, that attempts have been made to extend their validity to the stationary case far from equilibrium, cre- ating a new insight to search for a nonlinear theory for biological systems. Currently, there are several ideas that treat the evolu- tion of life and they postulate structural growth as the motor of evolution. However, the limitations to growth have been scarcely foreseen except for the criticism to Cope’s rule due to the advantages in volume growth. In this direction, the restrictions due to the surface increase of biological bodies are a topic that could be discussed in the future. The energy intensiveness hypothesis is similar regarding the capacity to process energy from the habitat. Nevertheless, it is not possible to generate or consume energy without wasting some to be removed as excess entropy, but it remains the key to achieve efficiency and even equally important, the survival of the species. The most appropriate critical view is thus established from the thermodynamic point of view, where the two aspects are included in the negentropy principle. In contrast, information theory, using Brillouin’s defi- nition of entropy, gives a new interpretation to thermo- dynamic entropy with its own methodology to measure it for small biological organisms, both at the microscopic and mesoscopic scales. The achievements show the de- scriptive capabilities of the theory. It is expected to achieve a more complete description of the principle of entropy balance on the phylogenic scale with the devel- opments in the techniques of advanced Monte Carlo [116], to verify the negentropy principle using informa- tion theory. The kinetic theory of chaos provides several possibili- ties of results that describe the change of stable dissipa- tive structures far from equilibrium. Two things are im- plicit: that evolution is continuous since the boundary conditions provide the information of mutations, and that the choice of one of the many predictions could be de- termined by natural selection. The image of evolution is that the adaptation process is a branch of a solution of a set of differential equations. This is interesting, since it shows a new insight from the kinetic point of view, which thermodynamics cannot describe. This is a plausible idea and contributions in this direction will be welcome.  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 388 Finally, it has been suggested to consider the social and cultural products of men, both social (work division and language) and intellectual in sciences and arts, as part of the characteristics of animal evolution guided by the principle of negentropic growth. In this sense, the evolutionary properties are transferred to those products in terms of the increase of organizational and hierarchi- cal levels [117] and differentiation [118], highlighting the abilities to transfer information and knowledge [119]. 6. Conclusion In the present work the nature of biological systems was described from a thermodynamic point of view. Two aspects of living matter have been identified. The first refers to mesoscopic aspects, which describe thermody- namically the physiological processes at the level of a cell or local scale, where entropy production is generated by the presence of irreversible flows. The second relates to the macroscopic aspects in the global scale, where the elimination of entropy through the surface of the bodies occurs, thus operating as open systems. At this level, the organization of a biological system is identified by the number of constrictions; which are equivalent to entropy reducers. The total provides the negentropy of the system. Regarding the division of the time scale, two are iden- tified: the ontogenic or the evolution of the prototype, and the phylogenic or the evolution of the species. In the ontogenic description there is a difficulty to restore the initial-operation state at the presence of stress; therefore a regression of fluctuations is required for maintaining the structure. The difficulty arises when stress accumu- lates, since the entropy produced by physiological proc- esses is not being eliminated from the system and there is no negentropy intake, then the open system has been blocked and is operating like a closed system, which could result in death. In the phylogenic description, the need for adaptation is established in the principle of minimum dissipation and maximum metabolic efficiency. Both evo- lutionary aspects are included in the negentropy principle. Moreover, the negentropy principle links two aspects: the necessity of adaptation to prevent the collective death; and the probabilistic aspect where mutations occur ran- domly or induced by the need of survival, enhancing the possibility of adaptation. Regarding the validity of the negentropy principle, in particular with the assumption of minimum energy dis- sipation, several efforts have been made to go beyond lin- ear equilibrium thermodynamics and improve its formu- lation. Encouraging results have been obtained, like those found in information theory, the energy intensiveness hypothesis, and the nonlinear stability theory of dissipa- tive structures far from equilibrium. In all these cases, the description of biological systems and their evolution is a task that is far from being finished. 7. Aknowledgements The authors acknowledge the financial support provided by DGPA-UNAM (Project IN112109). 8. References [1] D. Dix, “Toward a Definition of Life: Semantic and Thermodynamic Considerations,” Journal of Theoretical Biology, Vol. 102, No. 2, 1983, pp. 337-340. doi:10.1016/0022-5193(83)90371-5 [2] J. S. Wicken, “Entropy and Evolution: Ground Rules for Discourse,” Systematic Zoology, Vol. 35, No. 1, 1986, pp. 22-36. doi:10.2307/2413288 [3] D. W. McShea, “Possible Largest-Scale Trends in Organ- ismal Evolution: Eight ‘Live Hypotheses’,” Annual Review of Ecology and Systematics, Vol. 29, 1998, pp. 293-318. doi:10.1146/annurev.ecolsys.29.1.293 [4] J. Kestin, “A Course in Thermodynamics,” 2nd Edition, McGraw-Hill, New York, 1979. [5] E. Schrödinger, “What is Life? Mind and Matter,” Cam- bridge University Press, Cambridge, 1944. [6] L. Demetrius, “Directionality Principles in Thermody- namics and Evolution,” Proceeding of the National Academy of Sciences of the USA, Vol. 94, 1997, pp. 3491-3498. doi:10.1073/pnas.94.8.3491 [7] H. B. Hollinger and M. J. Zenzen, “An Interpretation of Macroscopic Irreversibility within the Newtonian Frame- work,” Philosophy of Science, Vol. 49, No. 3, 1982, pp. 309-354. doi:10.1086/289065 [8] L. Brillouin, “Maxwell’s Demon Cannot Operate: Infor- mation and Entropy. I,” Journal of Applied Physics, Vol. 22, No. 3, 1951, pp. 334-337. doi:10.1063/1.1699951 [9] L. Brillouin, “Physical Entropy and Information. II,” Journal of Applied Physics, Vol. 22, No. 3, 1951, pp. 338-343. doi:10.1063/1.1699952 [10] M. Ruse, “Monad to Man: The Concept of Progress in Evolutionaty Biology,” Harvard University Press, Mas- sachusetts, 1996. [11] I. Prigogine and T. M. Wiame, “Biologie et Thermody- namique del Phénomènes Irréversibles,” Experientia (Ba- sle), Vol. 2, 1946, pp. 451-453. [12] F. Crick, “Life Itself: Its Origin and Nature,” Simon and Schuster, New York, 1981. [13] J. Monod, “Chance and Necessity,” Random, New York, 1972. [14] M. Rossignol, L. Rossignol, R. A. A. Oldeman and S. Bensine-Tizroutine, “Struggle of Life or the Natural His- tory of Stress and Adaptation,” Treemail, The Nether- lands, 1998. [15] B. McDowell, “An Examination of the Ecosystems Per- spective in Consideration of New Theories in Biology  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 389 and Thermodynamics,” Journal of Sociology and Social Welfare, Vol. 21, No. 2, 1994, pp. 49-68. [16] J. G. Miller, “Living Systems: Basic Concepts,” Behav- ioral Science, Vol. 10, No. 3, 1965, pp. 193-237. doi:10.1002/bs.3830100302 [17] M. T. Hannan and J. Freeman, “The Population Ecology of Organizations,” The American Journal of Sociology, Vol. 82, No. 5, 1977, pp. 929-964. doi:10.1086/226424 [18] H. B. Callen, “Thermodynamics and an Introduction to Thermostatistics,” 2nd Edition, John Wiley & Sons, New York, 1960. [19] A. Katchalsky and P. F. Curran, “Nonequilibrium Ther- modynamics in Biophysics,” Harvard University Press, Massachusetts, 1981. [20] A. B. Pippard, “Elements of Classical Thermodynamics,” Cambridge University Press, Cambridge, 1957. [21] L. D. Landau and E. M. Lifshitz, “Statistical Physics,” 3rd Edition, (Part I by E. M. Liftshitz and L. P. Pi- taevskii), Pergamon Press, Oxford, 1980. [22] G. Lebon, D. Jou and J. Casas-Vázquez “Understanding non-equilibrium Thermodynamics. Foundations, Appli- cations, Frontiers”, Springer-Verlag, Berlin, 2008. doi:10.1007/978-3-540-74252-4 [23] E. B. Jacob, Y. Shapira and A. I. Tauber “Seeking the Foundations: From Schrödinger’s Negative Entropy to Latent Information,” Physica A, Vol. 359, 2006, pp. 495-524. doi:10.1016/j.physa.2005.05.096 [24] J. M. Rubi, “The Non-Equilibrium Thermodynamics Approach to the Dynamics of Mesoscopic Systems,” Journal of Non-Equilibrium Thermodynamics, Vol. 29, No. 4, 2004, pp. 315-325. doi:10.1515/JNETDY.2004.058 [25] D. Reguera, J. M. Rubí and J. M. Vilar, “The Mesoscopic Dynamics of Thermodynamic Systems,” Journal of Physical Chemistry B, Vol. 109, No. 46, 2005, pp. 21502- 21515. doi:10.1021/jp052904i [26] L. F. del Castillo, “El Fenómeno Mágico de la Ósmosis,” Colección Ciencia para Todos, Vol. 16, Fondo de Cultura Económica, México, 1996. [27] M. W. Zemansky and R. H. Dittman, “Heat and Thermo- dynamics: An Intermediate Textbook,” 6th Edition, McGraw-Hill, New York, 1981. [28] D. ter Haar and H. Wergeland, “Elements of Thermody- namics,” Addison-Wesley Publishing Company, Massa- chusetts, 1966. [29] L. Brillouin, “The Negentropy Principle of Information,” Journal of Applied Physics, Vol. 24, No. 9, 1953, pp. 1152-1163. doi:10.1063/1.1721463 [30] V. M. Zhukovsky, “Thermodynamics of Environment,” Journal of Mining and Metallurgy B, Vol. 36, No. 1-2, 2000, pp. 93-102. [31] R. Swenson and M. T. Turvey, “Thermodynamic Reason for Perception-Action Cycles,” Ecological Psychology, Vol. 3, No. 4, 1991, pp. 317-348. doi:10.1207/s15326969eco0304_2 [32] M. G. Velarde and C. Normand, “Convection,” Scientific American, Vol. 243, No. 1, 1980, pp. 93-108. [33] C. M. Visser and R. M. Kellogg, “Biorganic Chemistry and the Origin of Life,” Journal of Molecular Evolution, Vol. 11, No. 2, 1978, pp. 163-169. doi:10.1007/BF01733891 [34] H. F. Blum, “Time’s Arrow and Evolution,” 3rd Edition, Princeton University Press, Princeton, 1968. [35] G. Stent, “That was the Molecular Biology that was,” Science, Vol. 160, No. 3826, 1968, pp. 390-395. doi:10.1126/science.160.3826.390 [36] W. Arthur, “Mechanisms of Morphological Evolution: A Combined Genetic, Developmental and Ecological Ap- proach,” John Wiley & Sons, Chichester, 1984. [37] B. Alberts, A. Johnson, J. Lewis, M. Raff, K. Roberts and P. Walter, “Universal Mechanisms of Animal Develop- ment,” Molecular Biology of the Cell, 4th Edition, Gar- land Science, New York, 2002. [38] W. M. Bortz II, “Aging as Entropy,” Experimental Ger- ontology, Vol. 21, No. 4-5, 1986, pp. 321-328. doi:10.1016/0531-5565(86)90039-2 [39] W. Arthur, “Theory of the Evolution of Development,” John Wiley & Sons, New York, 1988. [40] O. Toussaint, P. Dumont, J. F. Dierick, T. Pascal, C. Frippiat, F. Chainiaux, F. Sluse, F. Eliaers and J. Remacle, “Stress-Induced Premature Senescence. Essence of Life, Evolution, Stress, and Aging,” Annals of the New York Academy of Sciences, Vol. 908, 2000, pp. 85-98. doi:10.1111/j.1749-6632.2000.tb06638.x [41] C. Darwin, “On the Origin of Species,” John Murray, London, 1859. [42] D. J. Depew and B. H. Weber, “Darwinism Evolving Systems Dynamics and the Genealogy of Natural Selec- tion,” The MIT Press, Cambridge, 1996. [43] J. Wu and W. Gao, “Spatial Patterns of Species Richness: A Hierarchical Perspective,” Chinese Biodiversity, Vol. 3, 1995, pp. 12-21. [44] R. E. Ulanowicz and B. M. Hannon, “Life and the Pro- duction of Entropy,” Proceedings of the Royal Society of London B, Vol. 232, No. 1267, 1987, pp. 181-192. doi:10.1098/rspb.1987.0067 [45] M. Ziehe and L. Demetrius, “Directionality Theory: an Empirical Study of an Entropic Principle in Life-History Evolution,” Proceedings of the Royal Society B, Vol. 272, No. 1568, 2005, pp. 1185-1194. doi:10.1098/rspb.2004.3032 [46] I. Prigogine, “Time, Structure and Fluctuations,” Science, Vol. 201, No. 4358, 1978, pp. 777-785. doi:10.1126/science.201.4358.777 [47] D. W. Hone and M. J. Benton, “The Evolution of Large Size: How does Cope’s Rule Work?” Trends in Ecology & Evolution, Vol. 20, No. 1, 2005, pp. 4-6. doi:10.1016/j.tree.2004.10.012 [48] J. G. Kingsolver and D. W. Pfennig, “Individual-Level Selection as a Cause of Cope’s Rule of Phyletic Size In- crease,” Evolution, Vol. 58, No. 7, 2004, pp. 1608-1612. doi:10.1111/j.0014-3820.2004.tb01740.x [49] B. van Valkenburgh, X. Wang and J. Damuth, “Cope’s Rule, Hypercarnivory, and Extinction in north American Canids,” Science, Vol. 306, No. 5693, 2004, pp. 101-104.  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 390 doi:10.1126/science.1102417 [50] J. A. Finarelli, “Testing Hypotheses of the Evolution of Encephalization in the Canidae (Carnivora, Mammalia),” Paleobiology, Vol. 34, No. 1, 2008, pp. 35-45. doi:10.1666/07030.1 [51] J. T. Bonner, “The Evolution of Complexity by Means of Natural Selection,” Princeton University Press, New Jer- sey, 1988. [52] M. T. Carrano, “Body-Size Evolution in the Dinosauria,” In: M. T. Carrano, R. W. Blob, T. J. Gaudin and J. R. Wible, Eds., Amniote Paleobiology: Perspectives on the Evolution of Mammals, Birds, and Reptiles, University of Chicago Press, Chicago, 2006, pp. 225-268. [53] D. Jou, J. E. Llebot and C. G. Perez, “Fisica para Cien- cias de la Vida,” 2nd Edition, McGraw Hill Interameri- cana, Madrid, 2008. [54] I. Prigogine, “Introduction to the Thermodynamics of Irreversible Processes,” Wiley & Sons, New York, 1955. [55] S. R. de Groot, “Thermodynamics of Irreversible Proc- esses,” North-Holland Publishing Company, Amsterdam, 1966. [56] F. J. Ayala, “The Concept of Biological Progress,” In: F. J. Ayala and T. Dobzhansky, Eds., Studies in the Phi- losophy of Biology: Reductionism and Related Problems, Macmillan, New York, 1974, pp. 339-354. [57] D. C. Fisher, “Progress in Organismal Design,” In: D. M. Raup and D. Jablonski, Eds., Patterns and Processes in the History of Life, Springer, Berlin, 1986, pp. 99-117. [58] J. S. Huxley, “Evolution: The Modern Synthesis,” Harper, New York, 1942. [59] J. W. Valentine, “Patterns of Taxonomic and Ecological Structure of the Shelf Benthos during Phanerozoic Time,” Palaeontology, Vol. 12, 1969, pp. 684-709. [60] G. G. Simpson, “The Meaning of Evolution,” Yale Uni- versity Press, New Haven, 1967. [61] D. M. Raup, “Testing the Fossil Record for Evolutionary Progress,” In: M. Nitecki, Ed., Evolutionary Progress, Chicago University Press, Chicago, 1988, pp. 293-317. [62] F. J. Ayala, “Can ‘Progress’ be Defined as a Biological Concept?” In: M. Nitecki, Ed., Evolutionary Progress, Chicago University Press, Chicago, 1988, pp. 75-96. [63] H. Selye, “A Syndrome Produced by Diverse noxious Agents,” Nature, Vol. 138, 1936, p. 32. doi:10.1038/138032a0 [64] H. Selye, “The General Adaptation Syndrome and the Diseases of Adaptation,” Journal of Clinical Endocri- nology, Vol. 6, No. 2, 1946, pp. 117-230. doi:10.1210/jcem-6-2-117 [65] R. S. Boardman and A. H. Cheetham, “Degrees of Col- ony Dominance in Stenolaemate and Gymnolaemate Bryozoa,” In: R. S. Boardman, A. H. Cheetham and W. A. Oliver, Eds., Animal Colonies: Development and Func- tion through Time, Dowden, Hutchinson & Ross, Stroudsburg, 1973. [66] S. J. Gould, “Wonderful Life,” Norton, New York, 1989. [67] P. D. Gingerich, “Quantification and Comparison of Evolutionary Rates,” American Journal of Science, Vol. 293, 1993, pp. 453-478. doi:10.2475/ajs.293.A.453 [68] W. Scharloo, “Canalization: Genetic and Developmental Aspects,” Annual Review of Ecology and Systematics, Vol. 22, 1991, pp. 65-93. doi:10.1146/annurev.es.22.110191.000433 [69] R. Lewin, “Patterns in Evolution: The New Molecular View,” W. H. Freeman & Company, New York, 1999. [70] J. S. Wicken, “Thermodynamics and the Conceptual Structure of Evolutionary Theory,” Journal of Theoreti- cal Biology, Vol. 117, No. 3, 1985, pp. 363-383. doi:10.1016/S0022-5193(85)80149-1 [71] C. Shannon, “A Mathematical Theory of Communica- tion,” Bell System Technical Journal, Vol. 27, 1948, pp. 379-423, 623-656. [72] E. T. Jaynes, “Information Theory and Statistical Me- chanics,” In: K. Ford, Ed., Statistical Physics, Benjamin, New York, 1963, pp. 181-218. [73] G. Chaitin, “On the Length of Programs for Computing Finite Binary Sequences,” Journal of the Association for Computing Machinery, Vol. 13, 1966, pp. 547-569. [74] G. Chaitin, “Randomness and Mathematical Proof,” Sci- entific American, Vol. 232, No. 5, 1975, pp. 47-52. doi:10.1038/scientificamerican0575-47 [75] L. Brillouin, “Science and Information Theory,” 2nd Edition, Academic Press, New York, 1962. [76] J. Kestin and J. R. Dorfman, “Course in Statistical Ther- modynamics,” Academic Press, New York, 1971. [77] F. Reif, “Fundamentals of Statistical and Thermal Phys- ics,” McGraw-Hill, Singapore, 1965. [78] D. R. Brooks, P. H. Leblond and D. D. Cumming, “In- formation and Entropy in a Simple Evolution Model,” Journal of Theoretical Biology, Vol. 109, No. 1, 1984, pp. 77-93. doi:10.1016/S0022-5193(84)80112-5 [79] D. R. Brooks and E. O. Wiley, “Evolution as an Entropic Phenomenon,” In: J. W. Pollard, Ed., Evolutionary The- ory: Paths to the Future, John Wiley and Sons, London, 1984, pp. 141-171. [80] J. Campbell, “Grammatical Man: Information, Entropy, Language, and Life,” Simon & Schuster, New York, 1982. [81] N. Wiener, “Cybernetics: Or Control and Communication in the Animal and the Machine,” MIT Press, Massachu- setts, 1948. [82] J. S. Wicken, “Entropy, Information, and Nonequilibrium Evolution,” Systematic Zoology, Vol. 32, No. 4, 1983, pp. 438-443. doi:10.2307/2413170 [83] L. Kari and L. F. Landweber, “Biocomputing in Ciliates,” In: M. Amos, Ed., Cellular Computing, Oxford Univer- sity Press, Oxford, 2003. [84] R. D. Knight, L. F. Landweber and M. Yarus, “How Mi- tochondria Redefine the Code,” Journal of Molecular Evolution, Vol. 53, No. 4-5, 2001, pp. 299-313. doi:10.1007/s002390010220 [85] G. Burger, I. Plante, K. M. Lonergan and M. W. Gray, “The Mitochondrial DNA of the Amoeboid Protozoon, Acanthamoeba Castellanii: Complete Sequence, Gene  L. F. DEL. CASTILLO ET AL. Copyright © 2011 SciRes. JMP 391 Content and Genome Organization,” Journal of Molecu- lar Biology, Vol. 245, No. 5, 1995, pp. 522-537. doi:10.1006/jmbi.1994.0043 [86] C. H. Waddington, “New Patterns in Genetics and Devel- opment,” Columbia University Press, New York, 1966. [87] C. J. Smith, “Problems with Entropy in Biology,” Bio- systems, Vol. 7, No. 2, 1975, pp. 259-265. doi:10.1016/0303-2647(75)90033-7 [88] N. H. Gregersen, “From Complexity to Life: on the Emergence of Life and Meaning,” Oxford University Press, New York, 2003. [89] L. Brillouin, “Thermodynamics and Information Theory,” American Scientist, Vol. 38, 1950, pp. 595-599. [90] P. A. Corning and S. J. Kline, “Thermodynamics, Infor- mation and Life. Revisited, Part I: ‘To Be or Entropy’,” Systems Research and Behavioral Science, Vol. 15, 1998, pp. 273-295. doi:10.1002/(SICI)1099-1743(199807/08)15: 4<273::AID-SRES200>3.0.CO;2-B [91] G. J. Vermeij, “Evolution and Escalation,” Princeton University Press, New Jersey, 1987. [92] G. J. Vermeij, “The Evolutionary Interaction among Spe- cies: Selection, Escalation, and Coevolution,” Annual Re- view of Ecology and Systematics, Vol. 25, 1994, pp. 219-236. doi:10.1146/annurev.es.25.110194.001251 [93] G. J. Vermeij, “Economics, Volcanoes, and Phanerozoic Revolutions,” Paleobiology, Vol. 21, No. 2, 1995, pp. 125-152. [94] M. Tribus and E. C. McIrvine, “Energy and Information,” Scientific American, Vol. 224, 1971, pp. 179-188. doi:10.1038/scientificamerican0971-179 [95] S. N. Salthe, “The Natural Philosophy of Work,” Entropy, Vol. 9, No. 2, 2007, pp. 83-99. doi:10.3390/e9020083 [96] E. Mayr, “Toward a New Philosophy of Biology,” Har- vard University Press, Massachusetts, 1988. [97] A. Pross, “The Driving Force for Life’s Emergence: Ki- netic and Thermodynamic Considerations,” Journal of Theoretical Biology, Vol. 220, No. 3, 2003, pp. 393-406. doi:10.1006/jtbi.2003.3178 [98] S. Chandrasekhar, “Hydrodynamic and Hydromagnetic Stability,” Clarendon, Oxford, 1961. [99] D. D. Joseph, “Stability of Fluid Motions,” Springer, Berlin, 1976. [100] A. Movchan, “The Direct Method of Lyapounov in Sta- bility Problems of Elastic Systems,” Journal of Applied Mathematics and Mechanics, Vol. 23, No. 3, 1959, pp. 483-693. doi:10.1016/0021-8928(59)90161-3 [101] A. J. Pritchard, “A Study of the Classical Problem of Hydrodynamic Stability,” IMA Journal of Applied Ma- thematics, Vol. 4, No. 1, 1968, pp. 78-93. doi:10.1093/imamat/4.1.78 [102] G. Nicoli and I. Prigogine, “Self-Organization in Non- Equilibrium Systems: From Dissipative Structures to Or- der through Fluctuations,” Wiley-Interscience, New York, 1977. [103] P. Glandsdorv and I. Prigogine, “Structure, Stabilité et Fluctuations,” Masson, Paris, 1971. [104] D. Kondepudi, R. J. Kaufman and N. Singh, “Chiral Symmetry-Breaking in Sodium-Chlorate Crystallization,” Science, Vol. 250, No. 4983, 1990, pp. 975-976. doi:10.1126/science.250.4983.975 [105] V. Castets, E. Dalos, J. Boissonade and P. de Kepper, “Experimental Evidence of Sustained Standing Turing- Type Nonequilibrium Chemical Patterns,” Physical Re- view Letters, Vol. 64, No. 24, 1990, pp. 2953-2956. doi:10.1103/PhysRevLett.64.2953 [106] Q. Ouyang and H. L. Swinney, “Transition from a Uni- form State to Hexagonal and Striped Turing Patterns,” Nature, Vol. 352, No. 6336, 1991, pp. 610-612. doi:10.1038/352610a0 [107] A. M. Turing, “The Chemical Basis of Morphogenesis,” Philosophical Transactions of the Royal Society of Lon- don. Series B - Biological Sciences, Vol. 237, No. 641, 1952, pp. 37-72. doi:10.1098/rstb.1952.0012 [108] M. Tabor, “Chaos and Integrability in nonlinear Dynam- ics: An Introduction,” Wiley-Interscience, USA, 1989. [109] P. Davies, “The New Physics: A Synthesis,” In: P. Davies, Ed., The New Physics, Cambrige Unversity Press, Cam- bridge, 1989. [110] J. Swift and P. C. Hohenberg, “Hydrodynamic Fluctua- tions at the Convective Instability,” Physical Review A, Vol. 15, No. 1, 1977, pp. 319-328. doi:10.1103/PhysRevA.15.319 [111] M. C. Cross and P. C. Hohenberg, “Pattern Formation Outside of Equilibrium,” Reviews of Modern Physics, Vol. 65, No. 3, 1993, pp. 851-1112. doi:10.1103/RevModPhys.65.851 [112] K. Ito and Y. P. Gunji, “Self-Organization of Living Sys- tems towards Criticality at the Edge of Chaos,” Biosys- tems, Vol. 33, No. 1, 1994, pp. 17-24. doi:10.1016/0303-2647(94)90057-4 [113] E. Bodenschatz, W. Pesch and G. Ahlers, “Recent De- velopments in Rayleigh-Bénard Convection,” Annual Re- view of Fluid Mechanics, Vol. 32, No. 1, 2000, pp. 709-778. doi:10.1146/annurev.fluid.32.1.709 [114] P. L. Engle, “Conjectures Concerning Complexity and Hierarchy,” Far from Equilibrium, Laurel Highlands Me- dia, Greensburg, 2002. [115] J. Walleczek, “Self-Organized Biological Dynamics and Nonlinear Control: Toward Understanding Complexity, Chaos and Emergent Function in Living Systems,” Cam- bridge University Press, Cambridge, 2000. [116] R. Luzzi, A. R. Vasconcelos and J. G. Ramos, “Predictive Statistical Mechanics: A Nonequilibrium Ensamble For- mulation,” Kluwer Academia Publishers, Dordrecht, 2002. [117] R. A. Eve, S. Horsfall and M. E. Lee, “Chaos, Complex- ity, and Sociology: Myths, Models, and Theories,” Sage Publications, London, 1997. [118] R. Boyd and P. J. Richerson, “Culture and the Evolution- ary Process,” University of Chicago Press, London, 1988. [119] L. L. Cavalli-Sforza and M. W. Feldman, “Cultural Trans- mission and Evolution: A Quantitative Approach,” Prince- ton University Press, New Jersey, 1981.
|