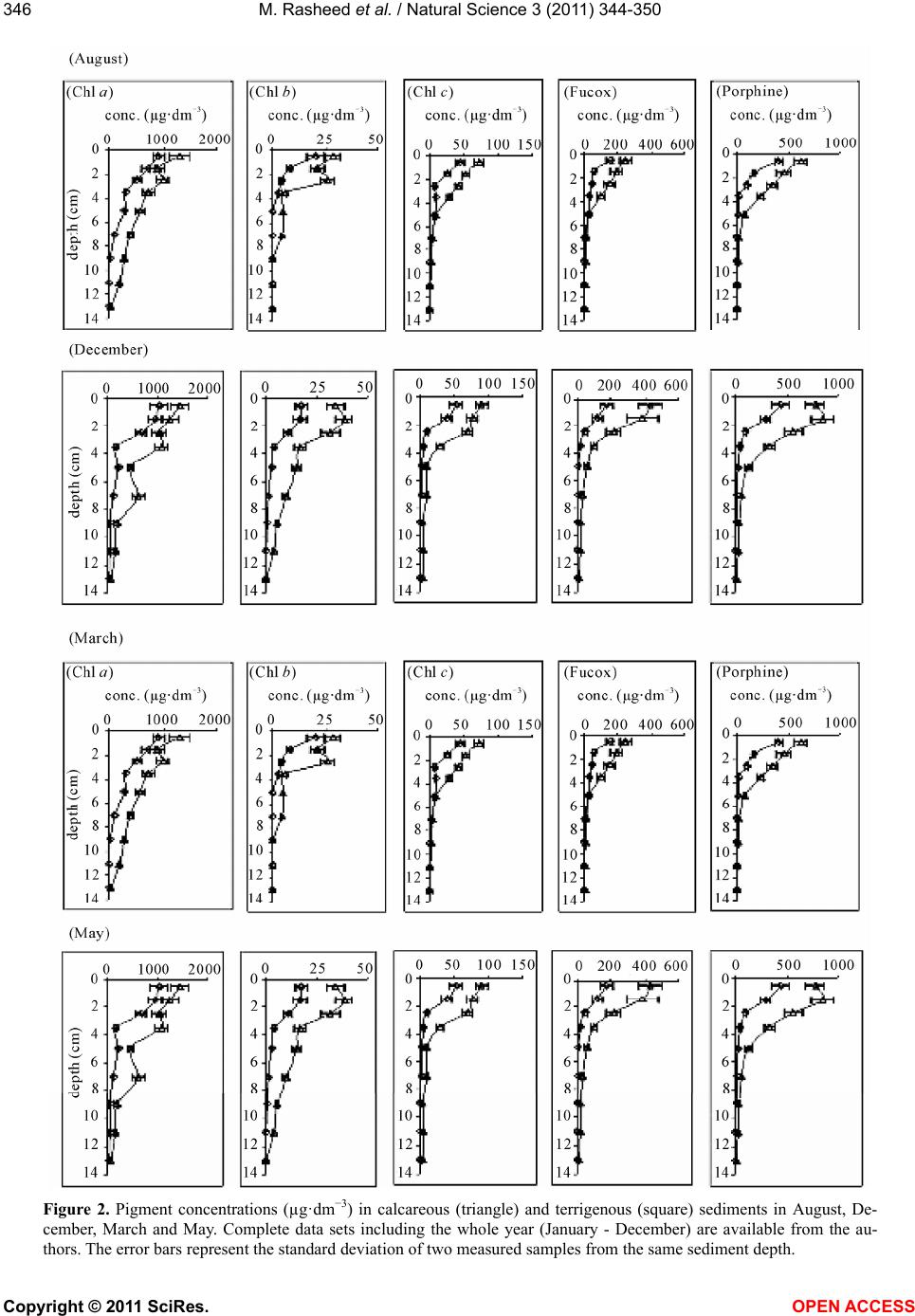
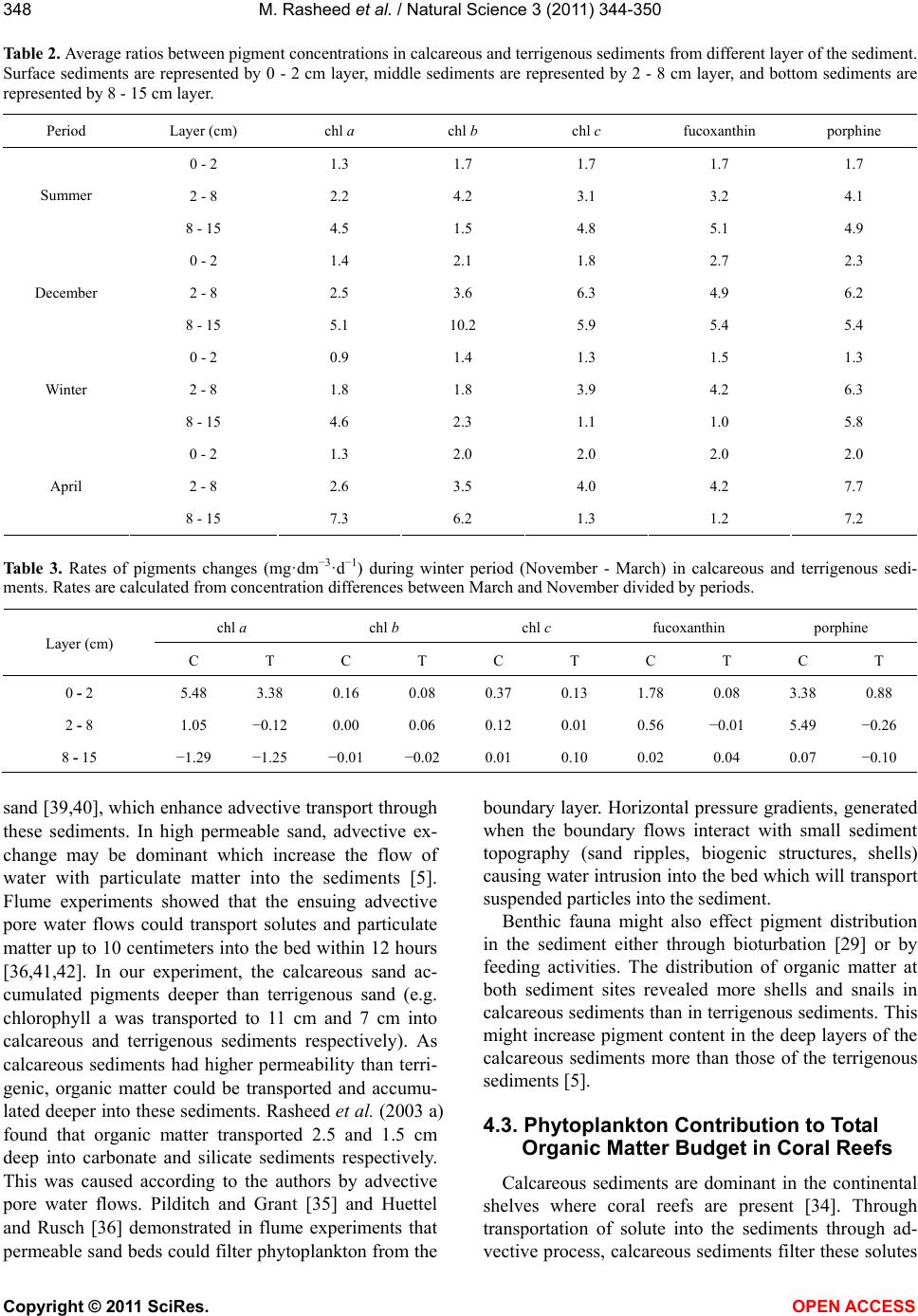
M. Rasheed et al. / Natural Science 3 (2011) 344-350
Copyright © 2011 SciRes. OPEN ACCESS
349
and the trapped particulate matter present. Phytoplank-
ton detritus can be also trapped in the sediments. In or-
der to calculate the contribution of phytoplankton detri-
tus in total organic matter deposited in calcareous and
terrigenous sediments, the conservative calculations us-
ing the accumulative rates of chl a and fucaxanthin in
the sediment and the pigment concentration percentage
for the total phytoplankton in the Gulf of Aqaba during
winter season were 28 and 12% respectively [20], how-
ever, we found phytoplankton contributions of 8 ± 3 and
6 ± 2% of organic matter to calcareous and terrigenous
sediments respectively. The accumulation of phyto-
plankton detritus and the subsequent degradation of
these detritus to inorganic nutrients may partly support
the productivity of the coral reef community which lives
in nutrient-poor environments.
5. ACKNOWLEDGEMENTS
Thanks are due to Khalid Altrabeen for his help in the lab. We would
like to thank Tariq Al-Salaman and Ali Hammad for helping in the
collection of the samples. This work has been funded by the Marine
Science Station, Aqaba-Jordan.
REFERENCES
[1] Charpy-Roubaud, C.J., Charpy, L. and Cremoux, J.L.
(1990) Nutrient budget of the lagoonal waters in an open
central South-Pacific atoll (Tikehau, Tuamotu, French
Polynesia). Marine Biology, 107, 67-73.
doi:10.1007/BF01313243
[2] Rysgaard, S., Thamdrup, B., Risgaard-Petersen, N.,
Fossing, H., Berg, P., Christensen, P.B. and Dalsgaard, T.
(1998) Seasonal carbon and nutrient mineralization in a
high-Arctic coastal marine sediment, young sound, North-
east Greenland. Marine Ecology Progress Series, 175,
261-276. doi:10.3354/meps175261
[3] Ciceri, G., Ceradini, S. and Zitelli, A. (1999) Nutrient
benthic fluxes and pore water profiles in a shallow brack-
ish marsh of the lagoon of Venice. Annali Di Chimica, 89,
359-375.
[4] Wild, C., Rasheed, M., Werner, U., Franke, U., Johnston,
R. and Huettel, M. (2004) Degradation and mineraliza-
tion of coral mucus in reef environments. Marine Ecol-
ogy - Progress Series, 267, 159-171.
doi:10.3354/meps267159
[5] Rasheed, M., Badran, M.I. and Huettel, M. (2003) Par-
ticulate matter filtration and seasonal nutrient dynamics
in permeable carbonate and silicate sands of the Gulf of
Aqaba, Red Sea. Coral Reefs, 22, 167-177.
doi:10.1007/s00338-003-0300-y
[6] Rasheed, M., Wild, C., Jantzen, C. and Badran, M. (2006)
Mineralization of particulate organic matter derived from
coral-reef organisms in reef sediments of the Gulf of
Aqaba. Chemistry & Ecology, 22, 13-20.
doi:10.1080/02757540500456823
[7] Wild, C., Naumann, M., Haas, A., Struck, U., Mayer, F.,
Rasheed, M. and Huettel, M. (2009) Coral sand O2 up-
take and pelagic-benthic coupling in a subtropical fring-
ing reef, Aqaba, Red Sea. Aquatic Biology, 6, 133-142.
doi:10.3354/ab00181
[8] Jorgensen, B.B. (1983) Processes at the sediment-water
interface. In: Bolin, B. and Cook, R.B. Eds., The Major
Biochemical Cycles and Their Interactions, John Wiley,
New York, 477- 515.
[9] Jorgensen, B.B. (1996) Material flux in the sediment. In:
Jorgensen, B.B. and Richardson, K. Eds., Eutrophication
in Coastal Marine Ecosystems, American Geophysical
Union, Washington, 115-135.
[10] Kristensen, E., Jensen, M.H., Jensen, K.M. (1997) Tem-
poral variations in microbenthic metabolism and inor-
ganic nitrogen fluxes in sandy and muddy sediments of a
tidally dominated bay in the northern Wadden Sea. Hel -
goland Marine Research, 51, 295-320.
[11] Wollast, R. (1991) The coastal organic carbon cycle:
Fluxes, sources, and sinks. In: Mantoura, R.F.C., Martin,
J.M. and Wollast, R. Eds., Ocean Margin Processes in
Global Change, Wiley, Chichester, 365-381
[12] Rasheed, M., Badran, M.I. and Huettel, M. (2003) Influ-
ence of sediment permeability and mineral composition
on organic matter degradation in three sediments from
the Gulf of Aqaba, Red Sea. Estuarine, Coastal and Shelf
Science, 57, 369-384.
doi:10.1016/S0272-7714(02)00362-1
[13] Friedman, G.M. (1968) Geology and geochemistry of
reefs, carbonate sediments, and waters, Gulf of Aqaba
(Elat), Red Sea. Journal of Sedimentary Research, 38,
895-919.
[14] Klute, A. and Dirksen, C. (1986) Hydraulic conductivity
and diffusivity: Laboratory methods. In: Klute, A. Ed.,
Methods of Soil Analysis - Part 1 - Physical and Minera-
logical Methods, American Society of Agronomy, Madi-
son, 687-734.
[15] Muller, G. (1967) Methods in sedimentary petrology.
Hafner Publishing Company, New York, 255.
[16] Sandstrom, M.W., Tirendi, F. and Nott, A. (1986) Direct
determination of organic carbon in modern reef sediments
and calcareous organisms after dissolution of carbonate.
Marine Geology, 70, 321-329.
doi:10.1016/0025-3227(86)90009-5
[17] Rusch, A., Forster, S. and Huettel, M. (2001) Bacteria,
diatoms and detritus in an intertidal sandflat subject to
advective transport across the water-sediment interface.
Biogeochemistry, 55, 1-27.
doi:10.1023/A:1010687322291
[18] Karsten, U. and Garcia-Pichel, F. (1996) Carotenoids and
mycosporine-like amino acid compounds in members of
the genus Microcoleus (cyanobacteria): A chemosys-
tematic study. Systematic and Applied Microbiology, 19,
285-294.
[19] Badran, M.I. and Foster, P. (1998) Environmental quality
of the Jordanian coastal waters of the Gulf of Aqaba, Red
sea. Aquatic Ecosystem Health & Management, 1, 75-89.
[20] Al-Najjar, T. and Rasheed, M. (2005) Zooplankton bio-
mass in the most northern tip of the Gulf of Aqaba, a case
study. Lebanese Science Journal, 6, 3-10.
[21] Rasheed, M., Badran, M.I., Richter, C. and Huettel, M.
(2002) Effect of reef framework and bottom sediment on
nutrient enrichment in a coral reef of the Gulf of Aqaba,
Red Sea. Marine Ecology-Progress Series, 239, 277-285.
doi:10.3354/meps239277