S. Imperial, J. J. Centelles
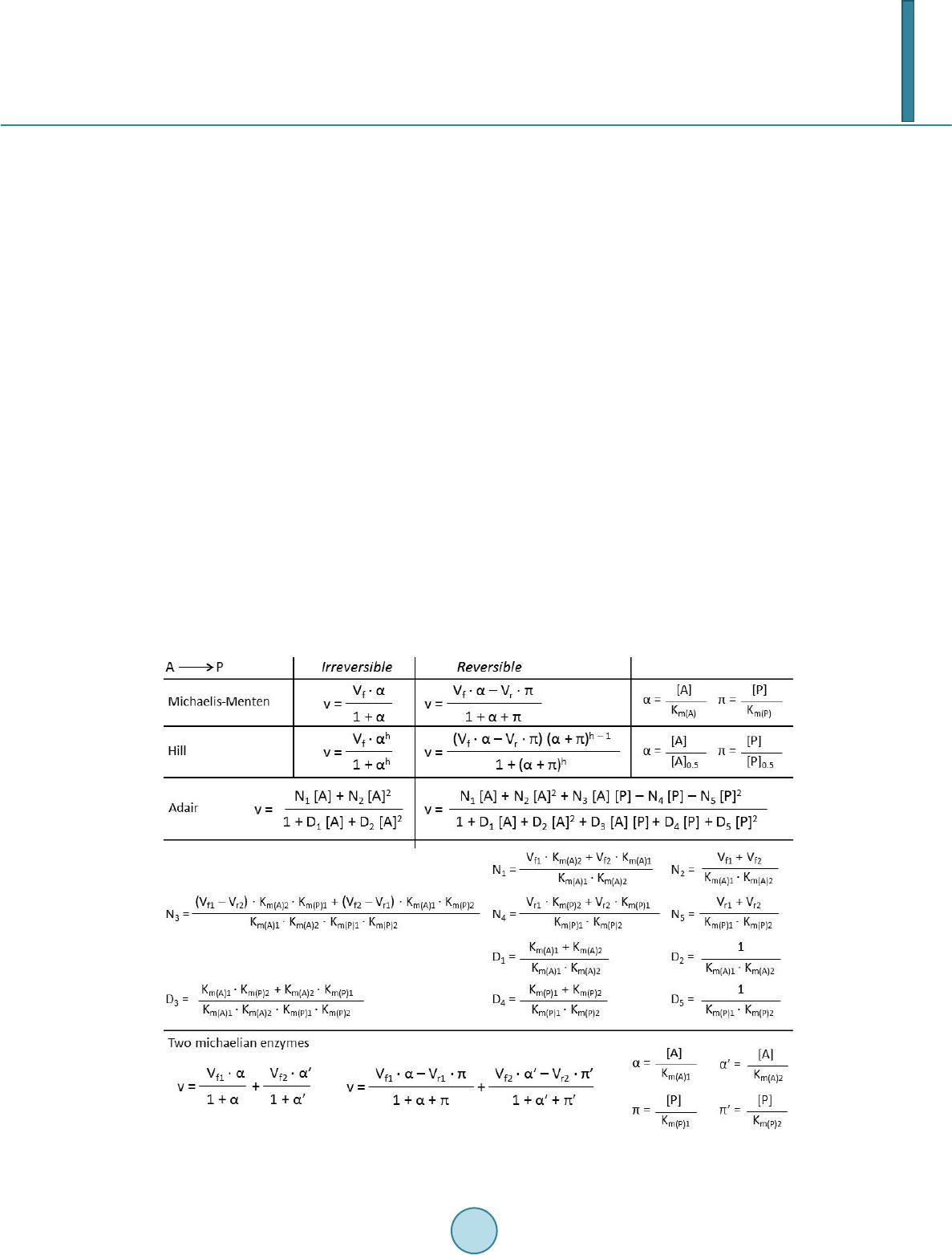
the reversible equation by considering [P] = 0.
Similar equations should be also obtained for the inverse irreversible reaction (P A) by considering [A] = 0
(or α = 0) although in these cases a negative equation is obtained, as the velocity is considered in the inverse
sense.
Haldane relationship from these reversible equations can be solved by considering that equilibrium concentra-
tions would led to v = 0 (for Michaelis-Menten and Hill equations, Vf αeq = Vr πeq), and Keq = [P]eq/[ A] eq. Thus,
resulting Haldane relationship to be: Keq = Vf KmP/Vr KmA.
2.2. Bi-Bi Mechanisms
Although there are several possible mechanisms, the most common bi-bi mechanisms include the ternary com-
plex mechanism (random or ordered bi-bi) and the substituted-enzyme mechanism (ping-pong bi-bi). As it can
be seen in Figure 2, the main difference between both mechanisms is the independent term present in the deno-
minator of the ternary complex mechanism. This independent term is present either for the irreversible or the
reversible equation.
These equations are more complex than those for the uni-uni mechanism, but it should be observed that both
irreversible equations can be simplified to a Michaelis-Menten uni-uni equation by considering saturated the
concentration of one of the substrates. For example, for a high β, v = Vf αβ/(β + αβ), and simplifying, v = Vf α/(1
+ α). Similarly, for a high α, the equation obtained would be v = Vf β/(1 + β).
The reversible equation, when considering high β, is transformed to an irreversible Michaelis-Menten equa-
tion, with inhibitions of products (P and Q). These inhibitions depend on the bi-bi mechanism considered, and
they follow the Cleland laws.
Haldane relationship from these equations can be solved by considering that equilibrium concentrations
would led to v = 0 (in both cases, Vf αeq βeq = Vr πeq ρeq), and Keq = [P]eq[Q]eq/[ A] eq[B]eq. Thus, resulting Haldane
relationship would be: Keq = Vf KmP KmQ/Vr KmA KmB.
2.3. Multisubstrate Mechanisms
We have considered until now uni-uni and bi-bi mechanism. Nevertheless, some reactions can be also uni-bi, or
bi-uni. Irreversible equations from uni-uni and from uni-bi are identical, as both consider only one substrate. But
reversible reactions present other factors in the numerators, as it can be seen in Figure 1 and Figure 2. Thus, for
uni-bi reactions, numerators of the equations can be easily corrected by using the uni positive factor and the bi
negative factor in the numerator: Vf α – Vr π ρ. And in the same way, the bi-uni equation should have the fol-
lowing factor as numerator: Vf α β – Vr π. In general, numerators can be deduced from the previous Figures. In
fact, if the reaction mechanism is known, the King and Altman method [3] can be used to deduce the enzymatic
equation. This method can be easily performed from the web page http://biokin.co m/king-a lt man/i nd ex. ht ml .
3. Some Examples of Reversible Reactions in Metabolism
Equilibrium constant can be used to calculate kinetic parameters by using Haldane relationship. For this reason,
we present in Table 1 a brief summary of some reversible reactions extracted from Barman [4]. Equilibrium
constants from the table are not taken all in the same conditions of pH and temperature, and the substrate or
products concentrations in the cell would indicate whether the reaction is far or near equilibrium. Reversible
Figure 2. Comparison between the reversible and irreversible kinetic equations of a bi-bi reaction.