M. Trifonov, V. Rozhkov
4. Conclusions and Future Work
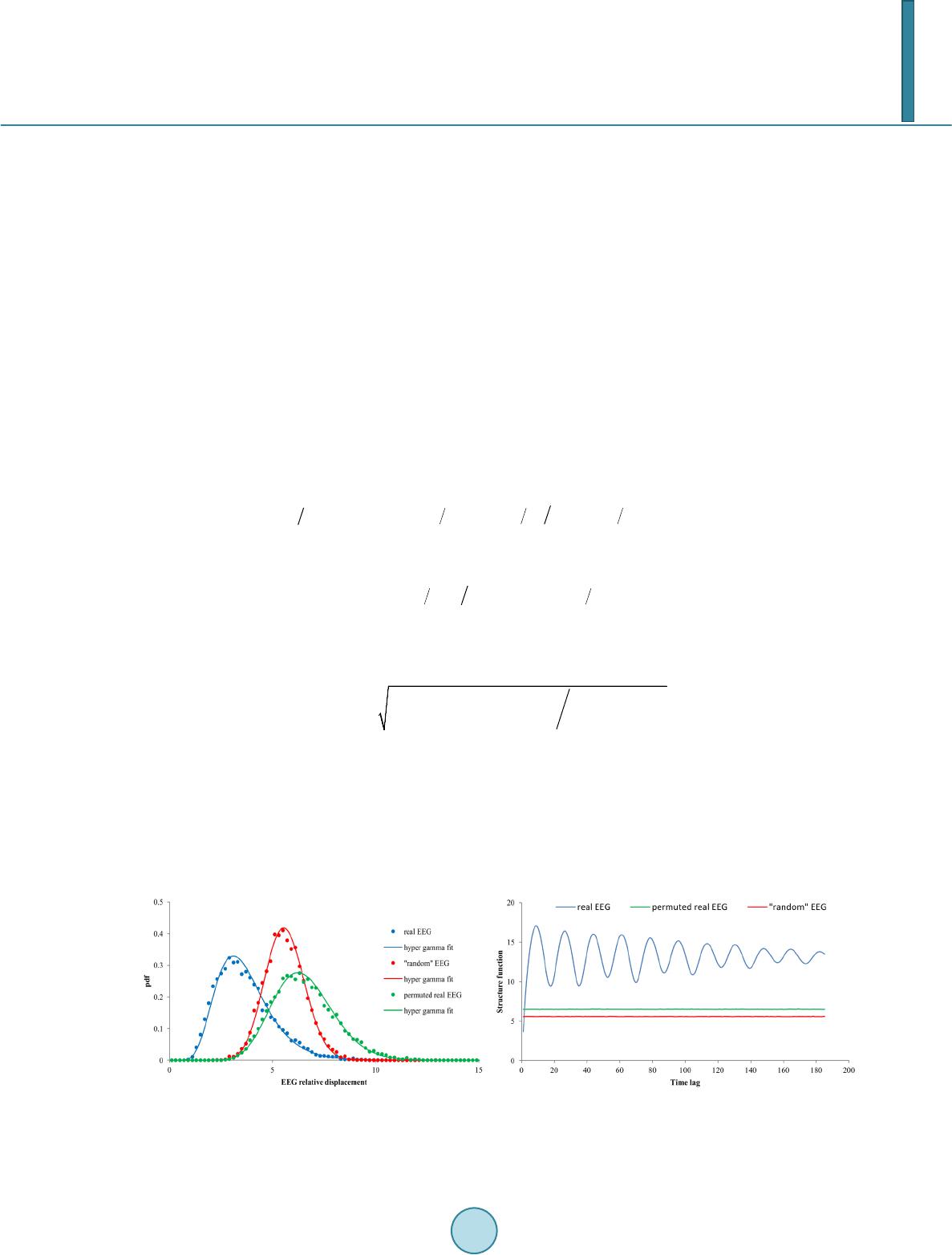
The results of this study allow us to make a preliminary conclusion that one-dimensional pdf’s of EEG relative
displace ments can be used for understanding of the real EEG dynamics in various functional states and different
subjects. To a first approximation, in each case the empirically derived pdf are fitted quite well by the single
hyper gamma distribution. It means that only two parameters (sample mean of EEG relative displacements and
coefficient of variation) may be taken into account. Both these parameters exhibit subject’s individuality. The
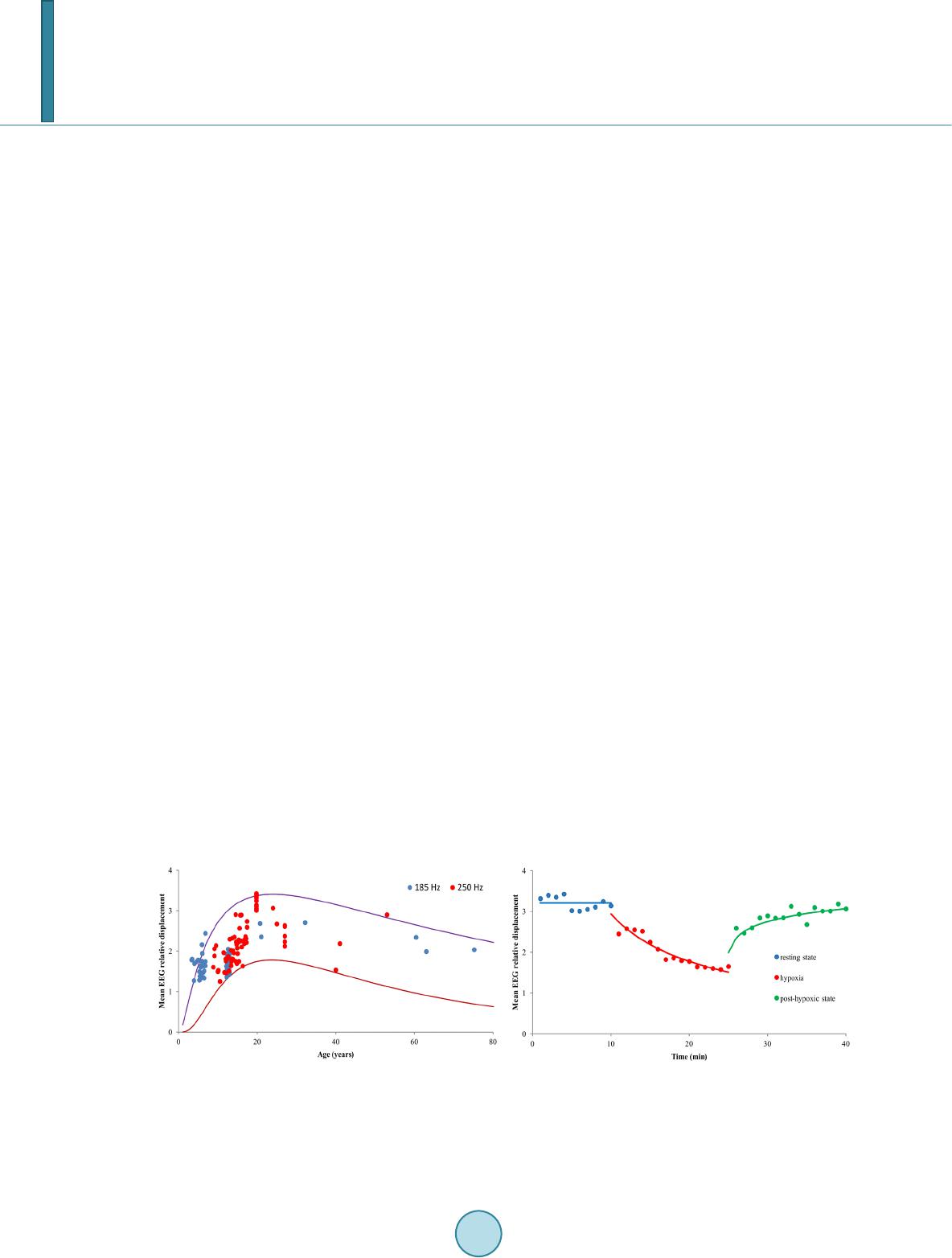
first one reveals age and state dependence while the second one stays rather stable for a given subject over long
period of time except sleep stages. It is interesting to note that age-related dependence of E[ΔY1] looks like as
age-related dependence of the total brain white matter volume given in [9]. In addition the non-linear age effect
on E[ΔY1] is consistent with the suggestion that during late childhood period there is a shift of topological or-
ganization of brain white matter toward a more randomized configuration [10].
In our future research we are going to analyze much more EEG records to investigate in details the age and
time-specific dependence of the parameters mentioned above. The next interesting aspect of our future work is
the improvement of the fitting quality of the empirically derived pdf of EEG relative displacements by using
distribution that consists of a mixture of hyper gamma distribution.
Acknowledgements
The authors would like to express their sincere thanks to Prof. M. N. Tsitseroshin, Dr. E. A. Panasevich, and Dr.
S. S. Bekshaev for providing EEG data.
References
[1] Gao, J., Hu, J. and Tung, W. (2011) Complexity Measures of Brain Wave Dynamics. Cognitive Neurodynamics, 5,
171-182. http://dx.doi.org/10.1007/s11571-011-9151-3
[2] Masquelier, T. (2013) Neural Variability, or Lack There of. Frontiers in Computational Neuroscience, 7, 1-7.
http://dx.doi.org/10.3389/fncom.2013.00007
[3] Osada, R., Funkhouser, T., Chazelle, B. and Dobkin, D. (2002) Shape Distributions. ACM Transactions on Graphics,
21, 807-832. http://dx.doi.org/10.1145/571647.571648
[4] Nikolova, N.D., Toneva-Zheynova, D., Kolev, K. and Tenekedjiev, K. (2013) Monte Carlo Statistical Tests for Identity
of Theoretical and Empirical Distributions of Experimental Data. In: Chan, V., Ed., Theory and Applications of Monte
Carlo Simulations, InTech, 1-26. http://dx.doi.org/10.5772/53049
[5] Suzuki, E. (1964) Hyper Gamma Distribution and Its Fitting to Rainfall Data. Papers in Meteorology and Geophysics,
15, 31-51. http://www.mri-jma.go.jp/Publish/Papers/DATA/VOL_15/15_031.pdf
[6] Izenman, A.J. (1991) Recent Developments in Nonparametric Density Estimation. Journal of the American Statistical
Association, 86, 205-224. http://www.jstor.org/stable/2289732
[7] Scott, D.W. (2004) Multivariate Density Estimation and Visualization. P apers / Humboldt-Universität Berlin, Center for
Applied Statistics and Economics (CASE), No. 2004, 16. http://hdl.handle.net/10419/22190
[8] Alfaouri, M., Daqrouq, K., Abu-Isbeih, I.N., Khalaf, E.F., Al-Qawasmi, A-R. and Al-Sawalmeh, W. (2009) Quality
Evaluation of Reconstructed Biological Signals. American Journal of Applied Science, 5, 187-193.
http://dx.doi.org/10.3844/ajas.2009.187.193
[9] Sowell, E.R., Peterson, B.S., Thompson, P.M., Welcome, S.E., Henkenius, A.L. and Toga, A.W. (2003) Mapping Cor -
tical Change across the Human Life Span. Nature Neuroscience, 6, 309-315. http://dx.doi.org/10.1038/nn1008
[10] Chen, Z., Liu, M., Gross, D.W. and Beaulieu, C. (2013) Graph Theoretical Analysis of Developmental Patterns of the
White Matter Netwo rk. Frontiers in Human Neuroscience, 7, 1-13. http://dx.doi.org/10.3389/fnhum.2013.00716