X. Xu, K. Zmolek
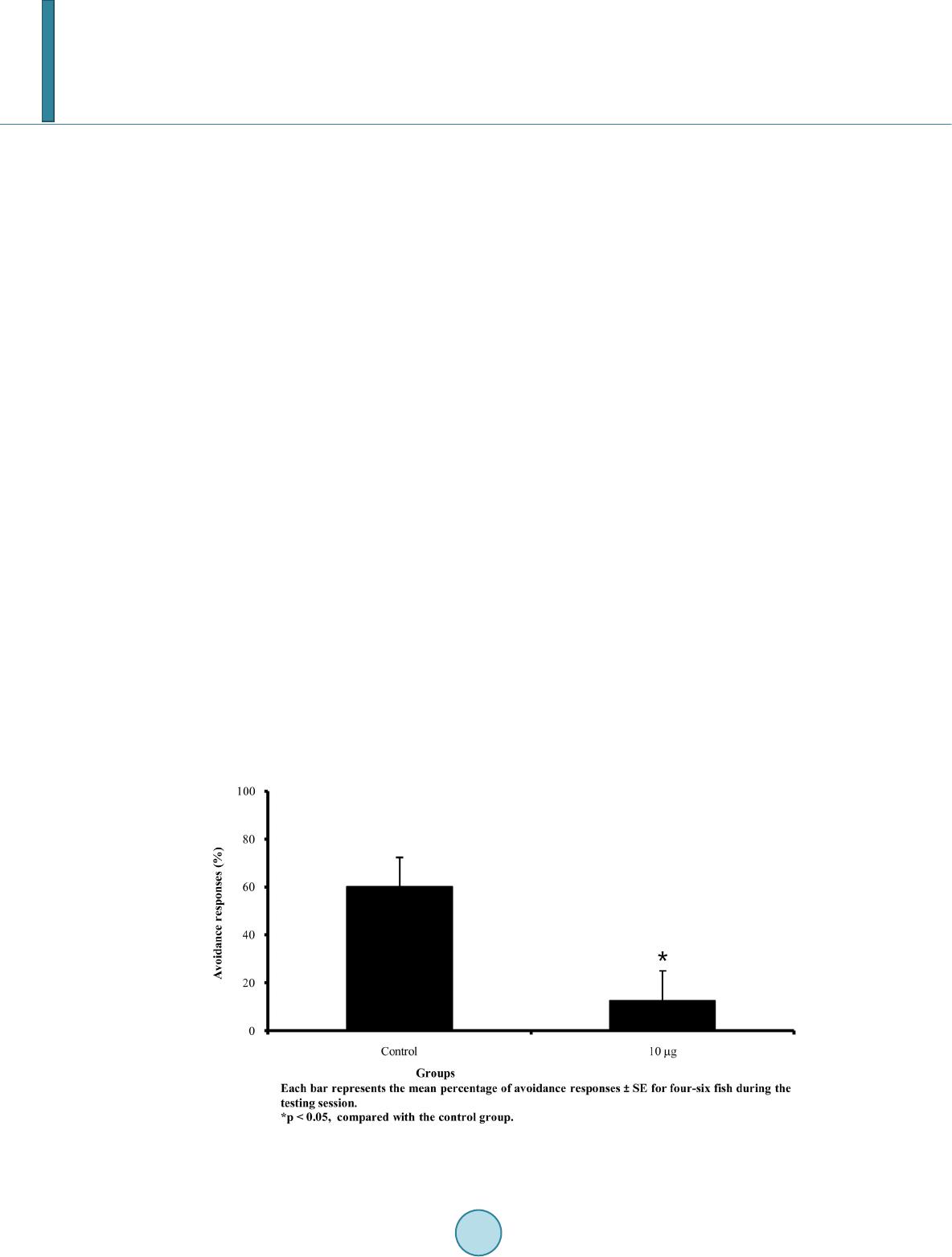
tions and fish that received saline injections immediately following training. Therefore, neither the microinjec-
tion procedure itself nor the microinjection of saline to the goldfish telencephalon impaired active avoidance
conditioning. Microinjections of carboxy-PTIO immediately following training impaired avoidance conditioning,
suggesting that extracellular NO may be necessary for forming memory of avoidance conditioning in goldfish.
The results of the current study confirm the finding that telencephalic NO is involved in memory consolidation
[10]. Thus, our previous results with D-AP5 [9] and current results with carboxy-PTIO together suggest that the
NMDA receptors are involved in learning or the process that is completed during training, whereas the NO is
involved in memory consolidation or the process that is normally completed sometime following the learning
experience.
Acknowledgements
This work was supported in part by a Grand Valley State University grant-in-aid.
References
[1] Gilbert, M.E. and Mack, C.M. (1990) The NMDA Antagonist, MK-801, Suppresses Long-Term Potentiation, Kindling,
and Kindling-Induced Potentiation in the Perforant Path of the Unanethetized Rat. Brain Research, 519, 89-96.
http://dx.doi.org/10.1016/0006-8993(90)90064-I
[2] Cotman, C.W., Monaghan, D.T. and Ganong, A.H. (1988) Excitatory Amino Acid Neurotransmission: NMDA Recep-
tors and Heb b-Type Synaptic Plasticity. Annual Review of Neuroscience, 11, 61-80.
http://dx.doi.org/10.1146/annurev.ne.11.030188.000425
[3] O’Dell, T.J., Hawkins, R.D., Kandel, E.R. and Arancio, O. (1991) Tests of the Roles of Two Diffusible Subtances in
Long-Term Potentiation: Evidence for Nitric Oxide as a Possible Early Retrograde Messenger. Proceedings of the Na-
tional Academy of Sciences of the United States of America, 88, 11285-11289.
http://dx.doi.org/10.1073/pnas.88.24.11285
[4] Lu, Y.-F., Kandel, E.R. and Kawkins, R.D. (1999) Nitric Oxide Signaling Contributes to Late-Phase LTP and CREB
Phosphorylation in the Hippocampus. Journal of Neuroscience, 19, 10250-10261.
[5] Bon, C.M. and Garthwaite, J. (2003) On the Role of Nitric Oxide in Hippocampal Long-Term Potentiation. The Jour-
nal of Neuroscience, 23, 1941-1948.
[6] Lynch, M.A., Errington, M.L., Clements, M.P., Bliss, T.V.P., Redini-Del Negro, C. and Laroche, S. (1990) Increases
in Glutamate Release and Phosphoinositide Metabolism Associated with Long-Term Potentiation and Classical Condi-
tioning. Progress in Brain Research, 83, 251-256. http://dx.doi.org/10.1016/S0079-6123(08)61254-6
[7] Xu, X., Russell, T., Bazner, J. and Hamilton, J. (2001) NMDA Receptor Antagonist AP5 and Nitric Oxide Synthase
Inhibitor 7-NI Affect Different Phases of Learning and Memory in Goldfish. Brain Research, 889, 274-277.
http://dx.doi.org/10.1016/S0006-8993(00)03216-9
[8] Xu, X., et al. (1998) Comparison of the Amnestic Effects of NMDA Receptor Antagonist MK-801 and Nitric Oxide
Synthase Inhibitors: L-NAME and L-NOARG in Goldfish. Behavioral Neuroscience, 112, 892-899.
http://dx.doi.org/10.1037/0735-7044.112.4.892
[9] Xu, X., Bazner, J., Qi, M., Johnson, E. and Freidhoff, R. (2003) The Role of Telencephalic NMDA Receptors in
Avoidance Learning in Goldfish (Carassius auratus). Behavioral Neuroscience, 117, 548-554.
http://dx.doi.org/10.1037/0735-7044.117.3.548
[10] Xu, X., Bently, J., Miller, T., et al. (2009) The Role of Telencephalic Nitric Oxide and cGMP in Avoidance Condi-
tioning in Goldfish (Carassius auratus). Behavioral Neuroscience, 123, 614-623. http://dx.doi.org/10.1037/a0015243