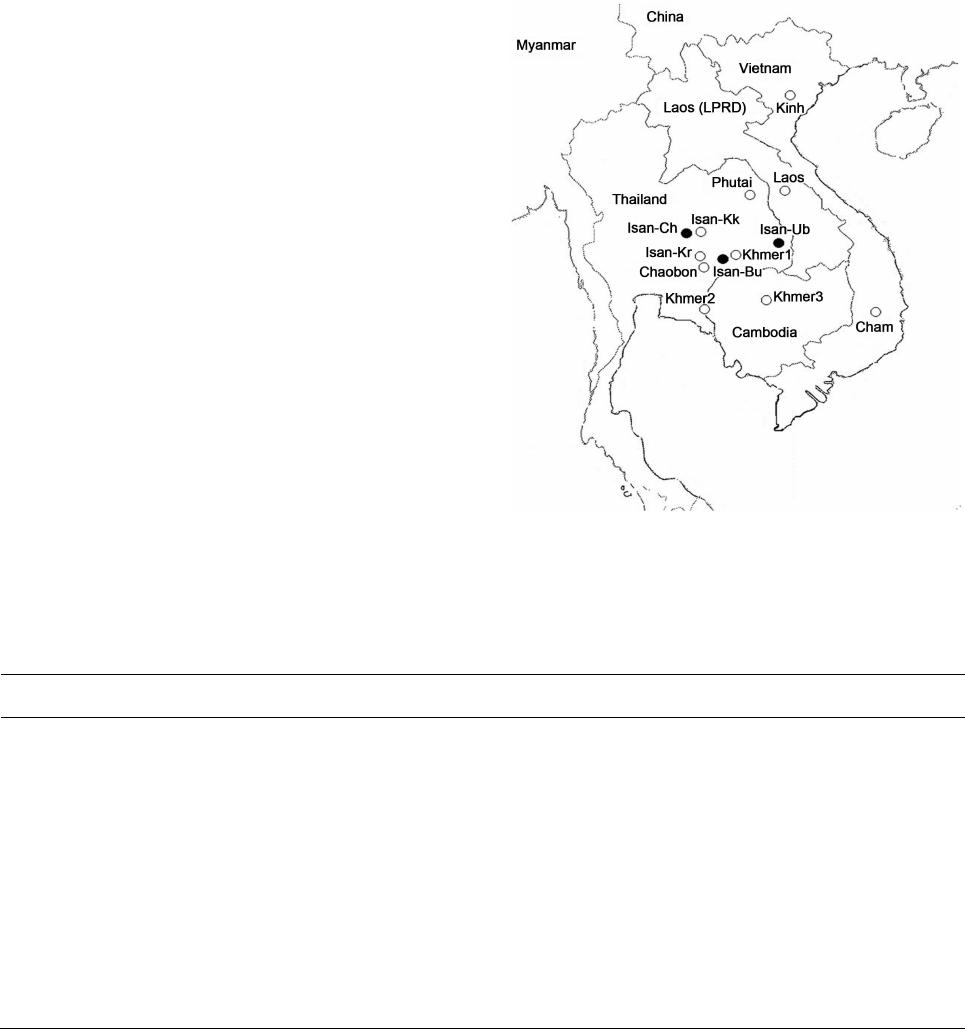
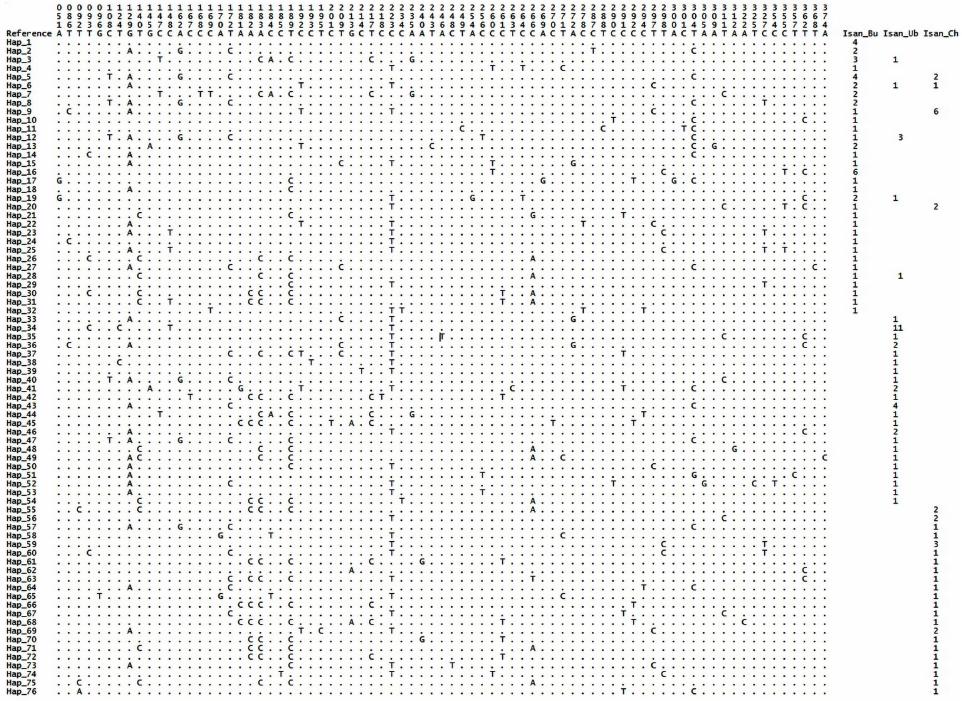
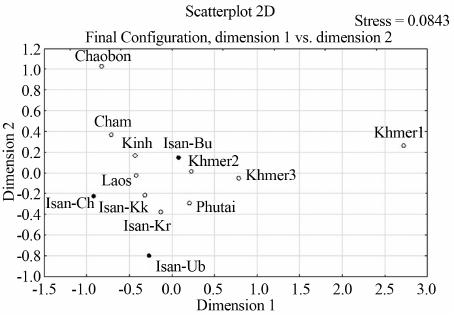
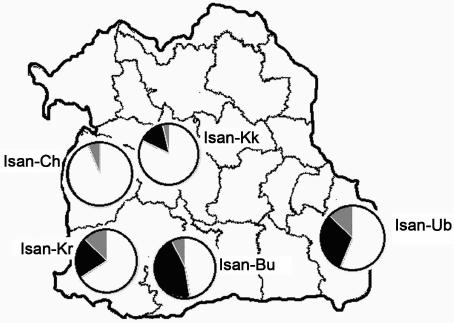
W. KUTANAN ET AL.
REFERENCES
Andrews, R. M., Kubacka, I., Chinnery, P. F., Lightowlers, R. N.,
Turnbull, D. M., & Howell, N. (1999). Reanalysis and revision of the
Cambridge reference sequence for human mitochondrial DNA. Na-
ture Genetics, 23, 147. http://dx.doi.org/10.1038/7727
Bertorelle, G., & Excoffier, L. (1998). Inferring admixture proportions
from molecular data. Molecular Biology and Evolution, 15, 1298-
1311. http://dx.doi.org/10.1093/oxfordjournals.molbev.a025858
Black, M. L., Dufall, K., Wise, C., Sullivan, S., & Bittles, A. H. (2006).
Genetic ancestries in n orthwest Ca mbodia. Annals of Human Biology,
33, 620-627. http://dx.doi.org/10.1080/03014460600882561
Bodner, M., Zi mmermann, B., Röck , A., Kloss-Brandstätter, A., Horst,
D., Horst, B., Sen gchanh, S., Sanguansermsri, T., Horst, J., Krämer,
T., Schneider, P. M., & Pa rson, W. (20 11). Southeast Asian diversity:
First insights into the co mplex mtDNA structure of Laos. BMC Evo-
lutionary Biol ogy, 11, 49. http://dx.doi.org/10.1186/1471-2148-11-49
Bonnie Pacala, B., & Somroay, Y. (2010). Buddhist murals of northeast
Thailand: Reflection of the Isan heartland. Chiangmai: Silkworm
Books.
Boonsoda, P., Srithawong, S., Srikuka, S., & Kutanan, W. (2013). Mi-
tochondrial DNA variation of the Khmer in Surin Provin ce, Thailand.
Thai Journal of Genetics, 6, 40-48.
Cavalli-Sforza, L. L., & Feldman M. W. (2003). The application of
molecular genetic approaches to the study of human evolution. Na-
ture Genetics, 33, 266-275. http://dx.doi.org/10.1038/ng1113
Davis, M. C., Novak, S. J., & Hampikian, G. (2011). Mitochondrial
DNA Analysis of an immigrant Basque p opulation: Loss of diversity
due to founder effects. American Journal of Physical Anthropology,
144, 516-525. http://dx.doi.org/10.1002/ajpa.21432
Dupanloup, I., & Bertorelle, G. (2001). Inferring admixture proportions
from molecular data: Extension to any number of parental popula-
tions. Molecular Biology and Evolution, 18, 672-675.
http://dx.doi.org/10.1093/oxfordjournals.molbev.a003847
Excoffie r, L., & Li sch er, H . E. L. (2010). Arleq uin su ite ver 3 .5: A n ew
series of programs to perform population genetics analyses under
Linux and Windows . Molecular Ecology Resources, 10, 564-567.
http://dx.doi.org/10.1111/j.1755-0998.2010.02847.x
Excoffier, L., Smouse, P., & Wuattro, J. (199 2). Analysis of molecular
variance inferred fro m metric distance a mong DNA ha plotypes: Ap-
plication to human mitochondrial DNA restriction data. Genetics,
131, 479-491.
Fu, Y. X. (1997). Statistical tests of n eutrality of mutations against po-
pulation growth, hitchhiking and background selection. Genetics,
147, 915-925.
Fucharoen, G., Fucharoen , S., & Horai, S. (2001). Mitoch ondrial DNA
polymorphisms in Thailand. Journal of Human Genetics, 46, 115-
125. http://dx.doi.org/10.1007/s100380170098
Harpending, H. C. (1994). Signature of ancient population growth in a
low-resolution mitochondrial DNA mismatch distribution. Human
Biology, 66, 591-600.
Hurles, M. E., Nicholson, J., Bosch, E., Renfrew, C., Sykes, B. C., &
Jobling, M. A. (2002). Y chromosomal evidence for the origins of
Oceanic-speaking peoples. Genetics, 160, 289-303.
Kampuansai, J., Bertorelle, G., Castri, L., Nakbunlung, S., Seielstad, M.,
& Kangwanpong, D. (2007). Mitochondrial DNA variation of Tai
speaking peoples in northern Thailand. Science Asia, 33, 443-448.
http://dx.doi.org/10.2306/scienceasia1513-1874.2007.33.443
Keyes, C. (1967) Isan: Regionalism in northeastern Thailand. New
York: Department of Asian Stu dies , Southeast Asia Pro gra m, Cornell
University.
Keyes, C. (1974). A note on the ancient towns and cities of northeastern
Thailand. Southeast Asian St udies, 11, 497-506.
Khanittanan, W. (20 01). Khmero-Thai: The great change in the histo ry
of Thai language in the Chao Praya basin. Journal of Language and
Linguistics, 19, 35-50.
Lertrit, P., Poolsuwan, S., Thosarat, R., Sanpach udayan, T., Boonyarit,
H., Chinpaisal, C., & Suktitipat, B. (200 8). Genetic history of south-
east Asian populations as revealed by ancient and modern human
mitochondrial DNA analysis. American Journal of Physical Anthro-
pology, 137, 425-440. http://dx.doi.org/10.1002/ajpa.20884
Lewis, M. P. (2009). Ethnologue: Languages of the world (16th ed.).
Dallas: SIL International. http://www.ethnologue.com/
Librado, P., & Rozas, J. (2009) DnaSP v5: A software for comprehen-
sive analysis of DNA polymorphism data. Bioinformatics, 25, 1451-
1452. http://dx.doi.org/10.1093/bioinformatics/btp187
Malyarchuk, B. A., Perkova, M. A., Derenko, M. V., Vanecek, T.,
Lazur, J., & Gomolcak, P. (2008 ). Mitochondrial DNA variability in
Slovaks, with application to the Roma origin. Annals of Human Ge-
netics, 72, 228-240.
http://dx.doi.org/10.1111/j.1469-1809.2007.00410.x
McCargo, D., & Hongladarom, K. (2004). Contesting Isan-ness: Dis-
courses of Politics and Identity in Northeast Thailand . Asian Ethnici-
ty, 5, 220-234. http://dx.doi.org/10.1080/1463136042000221898
Peng, M. S., Quang, H. H., Dang, K. P., Trieu, A. V., Wang, H. W.,
Yao, Y. G., Kong, Q. P., & Zhang, Y. P. (2010). Tracing the Aus-
tronesian footprint in mainland southeast Asia: A perspective from
mitochondrial DNA. Molecular Biology and Evolution, 27, 2417-
2430. http://dx.doi.org/10.1093/molbev/msq131
Premsrirat, S. (1999). Language and ethnicity on Kh o rat Plateau. In The
Princess Maha Chakri Sirindhorn Anthropology Centre, Thailand:
Culture and Society (pp. 267-284). B angkok: Ruen Kaew Printing.
Rogers, A. R., & Harpending, H. (1992). Population growth makes
waves in the distribution of pairwise genetic differences. Molecular
Biology and Evol ution, 9, 552-569.
Schliesinger, J. (2000). Ethnic groups of Thailand: Non-Tai-speaking
peoples. Bangkok: White Lotus Press.
Schliesinger, J. (2001). Tai Group of Thailand, Volume 1: Introduction
and overview. Bangkok: White Lotus Press.
Schurr, T. G., Sukernik, R. I., Starikovskaya, Y. B., & Wallace, D. C.
(1999). Mitocho ndrial DNA variation in Koryaks and It el’men: Pop-
ulation replacement in th e Okhotsk sea-Bering sea region during the
Neolithic. American Journal of P hysical Anthr opology, 108, 1-39.
http://dx.doi.org/10.1002/(SICI)1096-8644(199901)108:1<1::AID-A
JPA1>3.0.CO;2-1
Smalley, W. A. (1988 ). Multilingualis m in the Northern Kh mer popu la-
tion of Thailand. Language Sciences, 10, 395-408.
http://dx.doi.org/10.1016/0388-0001(88)90023-X
Smalley, W. A. (1994). Linguistic diversity and national unity: Lan-
guage ecology in Thailand. Chicago: University of Chicago Press.
Tajima, F. (1989). Statistical method for testing the neutral mutation
hypothesis by DNA polymorphism. Genetics, 123, 585-595.
Talbot, S., & Janthed, C. (2002). Northeast Thailand before Angkor:
Evidence from an Archaeological Excavation at the Prasat Hin Phi-
mai. Asia Perspectives, 40, 179-194.
http://dx.doi.org/10.1353/asi.2001.0027
Vail, P. (2006). Can a language of a million speakers be endangered?
Language shift and apathy among northern Khmer speakers in
Thailand. International Journal of the Sociology of Language, 178,
135-147.
Vail, P. (2007). Thailand’s Khmer as “invisible minority”: Language,
ethnicity and cultural politics in north-eastern Thailand. Asian Eth-
nicity, 8, 111-130. http://dx.doi.org/10.1080/14631360701406247
Welch, D. J. (1998). Archaeology of northeast Thailand in relation to
the pre-Khmer and Khmer historical records. International Journal
of Historical Archaeology, 2, 205-233.
http://dx.doi.org/10.1023/A:1027320309113
Winichakul, T. (2000). The quest for “Siwilai”: A geographical dis-
course of civilizational thinking in the late nineteenth and early
twentieth-century Siam. The Journal of Asian Studi es, 59, 528-549.
Wongtaed, S. (1999). Explore Isan society and cultures. Bangkok: Art
and culture Press.
Wyatt, D. K. (1969). The politics of reform in Thailand. New Haven:
Yale University Press.
Wyatt, D. K. (1984). Thailand: A short history. New Haven: Yale Uni-
versity Press.
OPEN ACCESS
12