Psychology
2014. Vol.5, No.1, 12-14
Published Online January 2014 in SciRes (http://www.scirp.org/journal/psych) http://dx.doi.org/10.4236/psych.2014.51003
Vection Is Not Determined by the Retinal Coordinate
Takeharu Seno1,2,3
1Faculty of Design, Kyushu University, Fukuoka, Japan
2Institute for Advanced Study, Kyushu University, Fukuoka, Japan
3Research Center for Applied Perceptual Science, Kyushu University, Fukuoka, Japan
Email: seno@design.kyushu-u.ac.jp
Received November 6th, 2013; revised December 5th, 2013; accepted January 3rd, 2014
Copyright © 2014 Takeharu Seno. T his is an open a ccess article dis tributed und er the Creative Co mmons Attri-
bution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the
original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights ©
2014 are reserved for SCIRP and the owner of the intellectual property Takeharu Seno. All Copyright © 2014
are guarded by law and by SCIRP as a guardian.
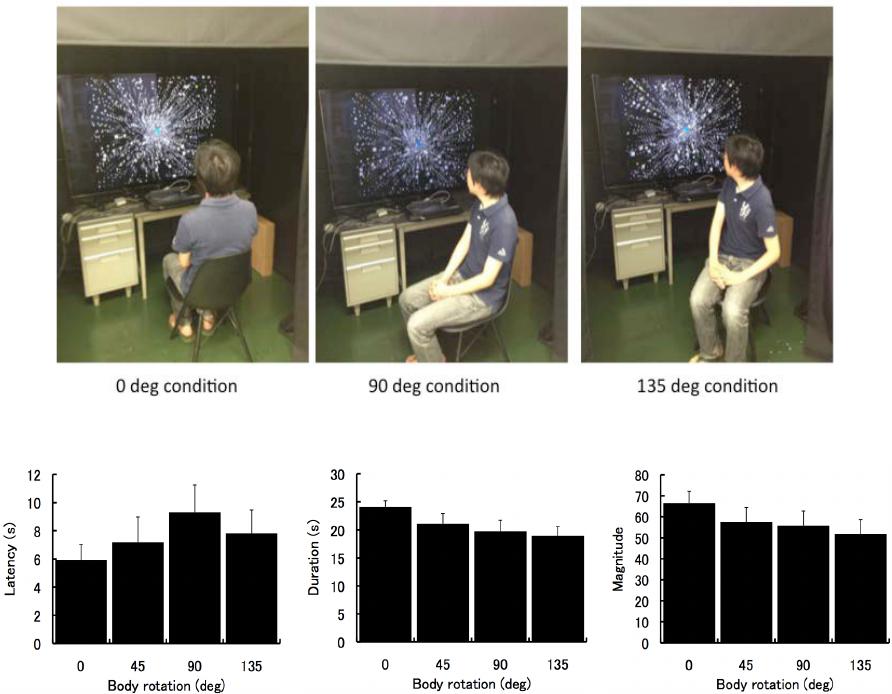
We examined whether vection was determined by the retinal coordinate. The participants sat on a chair
with their body rotated 0, 45, 90 and 135 degrees and with their head facing directly at the stimulus screen.
Vection strength was changed by rotation of the body. Thus vection was affected by the body coordinate
and it was not determined by the retinal coordinate alone.
Keywords: Retinal Coordinate; Head Coordinate; Body Coordinate; Body Rotation
Introduction
When stationary observers are exposed to a large-field visual
motion, simulating the retinal flow generated by self-translation
or self-rotation, they often experience an illusory perception of
self-motion, known as vection (Fischer & Kornmuller, 1930).
In the history of vection research, various stimulus attributes
which are effective or ineffective for vection induction have
been examined (e.g. Seno et al., 2009).
One important attribute is in which coordinate vection is de-
termined, whether in the retinal, body, head and/or world coor-
dinates. Morse & Rieser (2003) reported that vection is strongly
determined by the retinal coordinate. Kano (1991) also reported
that vection is affected both by retinal and world coordinates.
These results suggest that vection is affected by the retinal co-
ordinate.
Other studies reported that vection is affected by vestibular
inputs. Young & Shelhamer (1990) and Mueller & Kornilova
(1994) reported that vection in microgravity condition became
unstable and sometimes could be induced in the opposite direc-
tion to the normal vection. Lepecq et al. (2006) reported that by
adding a vestibular input during vection, the vection direction
could be biased intentionally. These results implied that vection
was not determined by the retinal coordinate alone.
Nakamura & Shimojo (1998) reported that the vertical vec-
tion was modulated by the body posture whereas the horizontal
vection was not modulated by the posture. This result implied
that vection is mediated by both retinal and body coordinates.
In this study, we set four different body postures with the
same retinal optic flow. The body was rotated horizontally 0, 45,
90 or 135 degrees with the head always directly facing the
screen. The participants twisted their head to face the screen. If
the vection is determined by the retinal coordinate, the vection
strength would be constant for each of the different body pos-
tures.
Method
Appara tus
Stimuli were generated and controlled by a computer (MB-
543J/A, Apple) and presented on a plasma display (3D Viera,
50-inch, Panasonic, with 1024 × 768 pixel resolution at a 60 Hz
refresh rate). The experiment was conducted in a dark chamber.
Parti cipa nts
Sixteen naïve volunteers participated in this experiment. Par-
ticipants were graduate or undergraduate students. All partici-
pants reported normal vision and no history of vestibular sys-
tem diseases. None of them was aware of the purpose of the
experiment.
Stimuli
Optic flow displays (101˚ × 80˚; presented for 30 s) consisted
of about 1000 randomly positioned dots per frame and the
global dot motion simulated forwards self-motion (16 m/s). The
screen size was bigger than that in our previous studies (Seno et
al., 2009, 2013). The stimuli were the same as those in Seno
(2013a & 2013b). There was no fixation point in the stimuli.
However, the participants were instructed to fixate on the focus
of the expansion (FOE). We think that the FOE would be the
role of the fixation point.
Procedure
Participants were asked to press a button when they per-
ceived forward self-motion, and keep the button depressed for
the duration of self-mot ion. After each trial, the participants
rated subjective vection strength using a 101-point rating
scale ranging from 0 (no vection) to 100 (very strong vec-
tion).
OPEN ACCESS
12