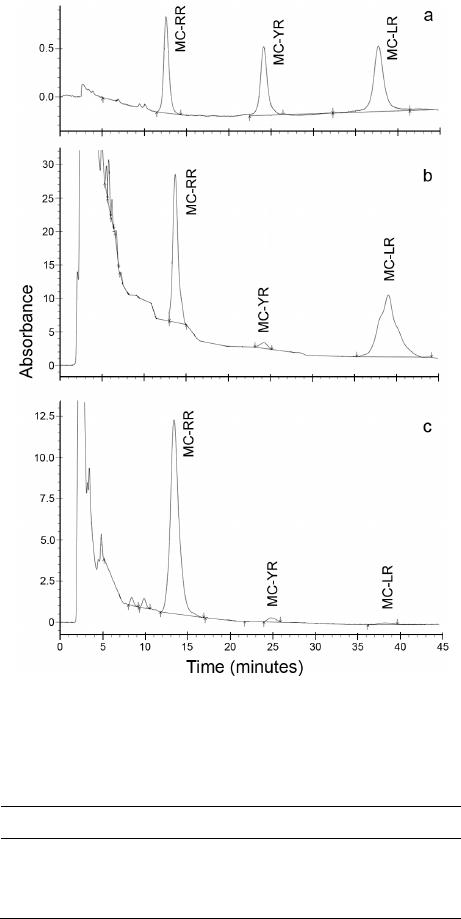
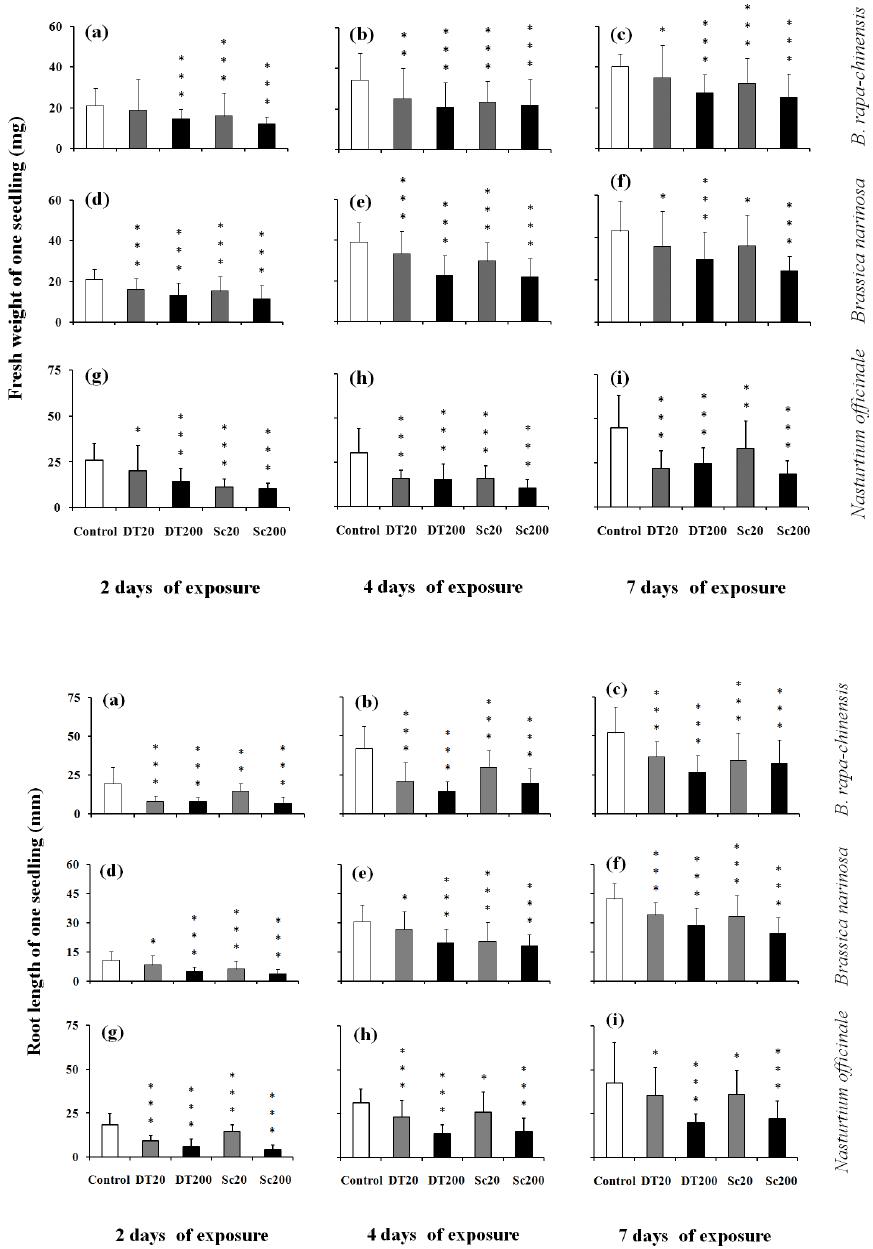
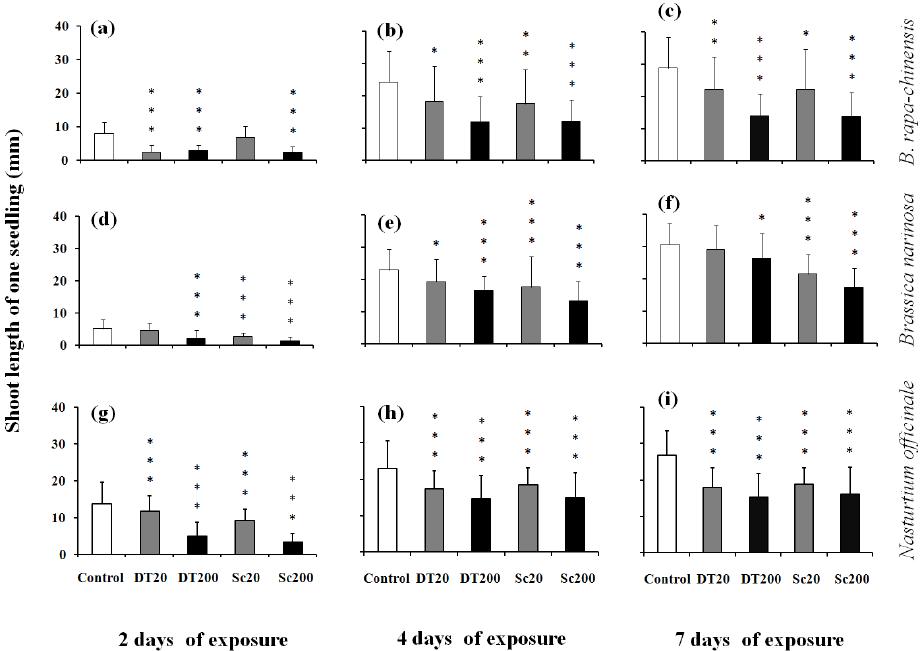
Influences of Cyanobacterial Toxins Microcystins on the Seedling of Plants
Figure 5. Normal leaves from control (a) and abnormal
leave s from M C ex posure (b) of Brassica rapa -chinensis at 7
days of incubation. Arrows indicate the difference in size
and the brown c olor at margin of the yo ung leaves.
be attributed to the impact of MC on chlorophyll content.
The results of this study confirm the potent toxicity of
cyanobacterial toxins from Dau Tieng Reservoir on
plants. To our knowledge, this is the first report on the
effects of MC on seedlings of B. rapa-chinensis, B. na-
rinosa and N. officinale.
Ackno wledgements
Thi s stud y is fund ed b y the Vietnam National University—
Hochiminh City under the granted projects numbers
B2012-24-01TD and A2013-48-01.
REFERENCES
[1] K. Sivonen and G. Jones, “Cyanobacterial Toxins,” In: I.
Chorus andJ. Bartram, Eds., Toxic Cyanobacteria in
Water—A Guide to Their Public Health Consequences,
Monitoring and Management, E & FN Spon, London,
1999, pp . 41-111.
[2] S. Pflugmacher, C. Wiegand, K. A. Beattie, G. A. Codd
and C. E. W. Steinberg, “Uptake of the Cyanobacterial
hepatotoxin Microcystin-LR by Aquatic Macrophytes,”
Journal of Applied Botany, Vol. 72, No. 5-6, 1998, pp.
228-232.
[3] S. Pflugmacher, C. Wiegand , K. A. Beat tie, E. Krause, C .
E. W. Steinberg and G. A. Codd, “Uptake, Effects and
Metabolism of Cyanobacterial Toxins in the Emergent
Reed Plant Phragmites australis (Cav.) Trin. ex Steud,”
Environmental Toxicology and Chemistry, Vol. 20, No. 4,
2001, pp . 846-852.
[4] K. Kurki-Helasmo and J. Meriluoto, “Microcystin Uptake
Inhibits Growth and Protein Phosphatase Activity in
Mustard (Sinapis alba L.) Seedlings,” Toxicon, Vol. 36,
No. 12, 1998, pp. 1921-1926.
http://dx.doi.org/10.1016/S0041-0101(98)00114-7
[5] S. M. Mitrovic, O. Allis, A. Furey and K. J. James, “Bio-
accumulation and Harmful Effects of Microcystin-LR in
the Aquatic Plants Lemna minor and Wolffia arrhiza and
the Filamentous Alga Chladophora fracta,” Exotoxicolo-
gy and Environmental Safety, Vol. 61, No. 3, 2005, pp.
345-352. http://dx.doi.org/10.1016/j.ecoenv.2004.11.003
[6] S. Jarvenpaa, C. Lundberg-Niinisto, L. Spoof, O. Sjovall,
E. Tyystjarvi and J. Meriluoto, “Effects of Microcystins
on Broccoli and Must ard , and Analysis of Accumulated
Toxin by Liquid Chromatography-Mass Spectrometry,”
Toxicon, Vol. 49, No. 6, 2007, pp. 865-874.
http://dx.doi.org/10.1016/j.toxicon.2006.12.008
[7] G. A. Codd, J. S. Metcalf and K. A. Beattie, “R etentio n of
Microcystis aeruginosa and Microcystin b y Salad Lett uce
(Lactuca sativa) after Spray Irrigation with Water Con-
taini ng Cyano bact eria, ” To xicon, Vol. 37, No. 8, 1999, pp.
1181-1185.
[8] J. McElhiney, LA. Lawton and C. Leifert, “Investigations
into the Inhibitory Effects of Microcystins on Plant
Growt h , and the Toxicity of Plant Tissues Following Ex-
posure,” Toxicon , Vol. 39, No. 9, 2001, pp. 1411-1420.
http://dx.doi.org/10.1016/S0041-0101(01)00100-3
[9] J. Chen, L. Song, J. Dai and Z. Liu, “ Effect s o f M icro cys -
tins on the Growth and the Activity of Superoxide Dis-
mutase and Peroxidase of Rape (Brassica napus L. ) and
Rice (Oryza sativa L.),” Toxicon, Vol. 43, No. 4, 2004,
pp. 393-400.
http://dx.doi.org/10.1016/j.toxicon.2004.01.011
[10] S. Pflugmacher, M. Aulhorn and M. Grimm, “Influence
of a Cyanobacterial Crude Extract Containing Microcys-
tin-LR on the Physiology and Antioxidative Defence
Systems o f Differen t Spi nach V ariants, ” New Phyco logist,
Vol. 175, No. 3, 2007, pp . 482-489.
http://dx.doi.org/10.1016/j.toxicon.2004.01.011
[11] J. Weiss, H. P. Liebert and W. Braune, “Influence on
Microcystin-RR on Growth and Photosynthetic Capacity
of the Duckweed Lemma minor L.,” Journal of Applied
Botany, Vol. 74, No. 3-4, 2000, pp. 10-105.
[12] M. H. Ha and S. Pflugmacher, “Phytotoxic Effects of the
Cyanobacterial Neurotoxin Anatoxin-a: Morphological,
Physiological and Biochemical Responses in Aquatic
Macrophyte, Ceratophyllum demersum,” Toxicon, Vol.
70, 2013 , pp. 1-8.
http://dx.doi.org/10.1016/j.toxicon.2013.03.021
[13] C. Pietsch, C. Wiegand, M. V. Ame, A. Nicklisch, D.
Wunderlin and S. Pflugmacher, “The Effects of Cyano-
bacterial Crude Extract on Different Aquatic Organisms:
Evidence for Cyanobacterial Toxin Modulating Factors, ”
Environmental Toxicology, Vol. 16, No. 6, 2 00 1, pp . 535-
542. http://dx.doi.org/10.1002/tox.10014
[14] C. Wiegand, A. Peuther t, S. Pflug macher and S. Carmeli,
“Effects of Microcin SF608 and Microcystin-LR, Two
Cyanobacterial Compounds Produced by Microcystis sp.,
on Aquatic Organisms,” Environmental Toxicology, Vol.
17, No. 4, 2002, pp . 400-406.
http://dx.doi.org/10.1002/tox.10065
[15] J. Stuven and S. Pflugmacher, “Antioxidative Stress Re-
sponse of Lepidium sativum Due to Exposure to Cyano-
bacterial Seco ndar y Metabo li tes,” Toxicon, Vol. 50, No. 1,
2007, pp . 85-93.
OPEN ACCESS JEP