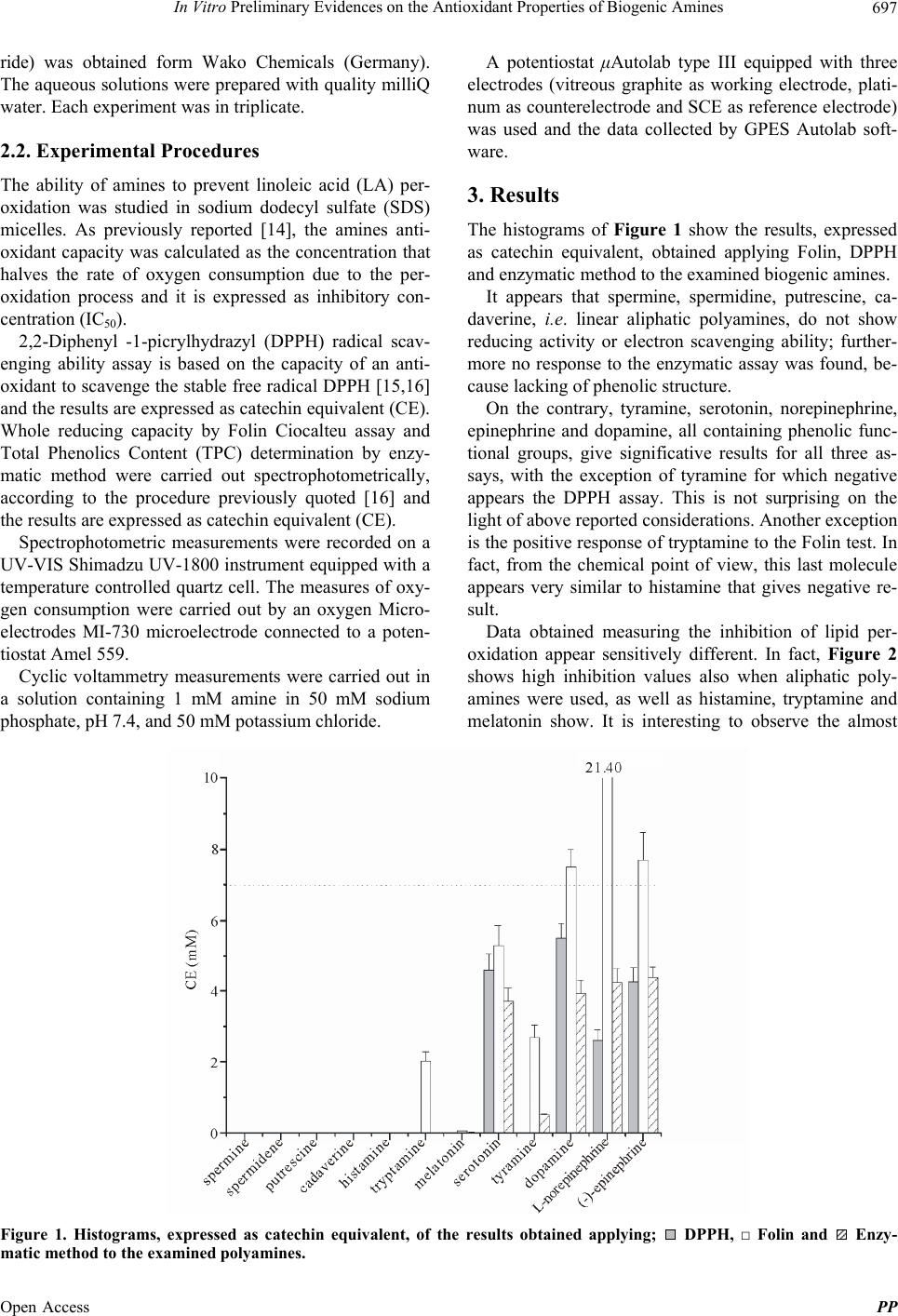
In Vitro Preliminary Evidences on the Antioxidant Properties of Biogenic Amines 699
Table 1. Discharge potential vs SCE of biogenic amines. A
vitrous graphite as working electrode was used.
Amine E(V)
spermine 1
spermidine 1
putrescine 1
cadaverine 1
histamine 0.765 ± 0.001
tryptamine 0.805 ± 0.003
melatonin 0.680 ± 0.001
serotonin 0.430 ± 0.005
tyramine 0.692 ± 0.006
dopamine 0.196 ± 0.008
L-norepinephrine 0.302 ± 0.007
(-)-epinephrine 0.311 ± 0.007
4. Discussion
As previously reported [14], no analytical method can
give a reliable measure of the antioxidant efficacy of a
molecule, because of the different parameters that each
assay measures and the manifold types of reactions in-
volved in the ROS damage. In fact, biogenic amines
show very different chemical structures and can be as-
sembled into three groups.
Besides primary amine group, they are characterized
respectively: 1) by aliphatic chain spaced or ended by
other secondary or primary amine groups respectively
(spermine, spermidine, putrescine, cadaverine); 2) by a
terminal imidazole (histamine) and indole group (tryp-
tamine, melatonin and serotonin); 3) by phenolic (tyra-
mine and serotonin again) or polyphenolic (L-norepine-
phrine, (-)-epinephrine, dopamine) function.
For this reasons in this work, multip le tests, represent-
ing various redox mechanisms, have been used as the
best strategy for stating an empirical and approximate
scale of effectiveness. In fact, the well-known Folin-
Ciocalteu assay is a general measure of antioxidant’s
reducing capacity and it is largely a specific because
correlated only to the redox potential of the chemical
species involved; DPPH based method measures the
ability of a molecule to scavenge the stable free radical
DPPH, but this last compound may be inert to many an-
tioxidant and the reaction could not go to completion, as
found for some phenolic derivatives [19]; enzymatic
method, we proposed, determines phenolic structures
owing to peroxidise specificity [16]. In this paper, in ad-
dition to these analytical methods, the measures of the
inhibitory capacity of ABIP-induced lipid peroxidation
and of the electrochemical ability to give electrons to
oxidative species are also carried out.
The experimental data indicate that all examined bio-
genic amines show antioxidant properties; in particular,
tyramine, serotonin, L-norepinephrine, (-)-epin ephrine and
dopamine for their (poly)phenolic structure too, while
aliphatic polyamines, histamine, tryptamine and mela-
tonin appear to act specifically on the oxygen con-
suming species involved in the lipid peroxidation of
polyunsaturated fatty acids in biological systems.
Considering that spermine is coupled by electrostatic
bonds to phosphate groups of nucleic acids and mem-
brane phospholip ids, stabilizing helical and bilayer struc-
tures, these results suggest that spermine and probably
other polyamines also could carry out a protective ac-
tion of nucleotide bases and acyl chains of phospholipids
against ROS injuries. Moreover, it is necessary to take
into consideration that polyamines oxidation, consequent
their protective action, decreases their stabilizing role
towards the above reported structures.
Furthermore, catecholamines, owing to effective cate-
chol group which can oxidatively convert to the corre-
sponding o-quinones, can perform a protective function
towards synaptic vesicles and, in particular, their phos-
pholipid moiety from ROS induced peroxidation, but
their loss contribute to neurodegenerative disorder, nota-
bly Parkinson’s disease [2]. Moreover, quinone structure
can easily react with primary amine groups of biogenic
amines or proteins to form Schiff base.
Further investigations to explain a possible chemical
mechanism justifying the antioxidant properties of bio-
genic amines are in progress.
REFERENCES
[1] S. H. Snyder, “Drug and Neurotransmitter Receptors in
the Brain,” Science, Vol. 224, No. 4644, 1984, pp. 22-31.
http://dx.doi.org/10.1126/science.6322304
[2] S. Surendran and S. Rajasankar, “Parkinson’s Disease:
Oxidative Stress and Therapeutic Approaches,” Neuro-
logical Sciences, Vol. 31, No. 5, 2010, pp. 531-540.
http://dx.doi.org/10.1007/s10072-010-0245-1
[3] C. Tabor and H. Tabor, “Polyamines,” Annual Review of
Biochemistry, Vol. 53, 1984, pp. 749-790.
http://dx.doi.org/10.1146/annurev.bi.53.070184.003533
[4] R. Kaur-Sawhney, A. Altman and W. Galston, “Dual
Mechanisms in Polyamine-Mediated Control of Ribonu-
clease Acti vity in Oat Lea f Protopla sts, ” Plant Physiology,
Vol. 2, 1978, pp. 158-160.
http://dx.doi.org/10.1104/pp.62.1.158
[5] R. Stevanato, A. Wisniewska and F. Momo, “Interaction
of Spermine with Dimyristoyl-L-alpha-phosphatidyl-DL-
glycerol Multilamellar Liposomes,” Archives of Biochem-
istry and Biophysics, Vol. 346, No. 2, 1997, pp. 203-207.
http://dx.doi.org/10.1006/abbi.1997.0253
[6] F. Momo, S. Fabris and R. Stevanato, “Interaction of
Open Access PP