 Psychology 2013. Vol.4, No.12, 998-1003 Published Online December 2013 in SciRes (http://www.scirp.org/journal/psych) http://dx.doi.org/10.4236/psych.2013.412144 Open Access 998 Dependence of Manual Grasping on the Behavioral Context: A Comparison between Arms and between Age Groups Otmar Bock, Benjamin Baak Institute of Physiology and Anatomy,German Sport University, Cologne, Germany Email: bock@dshs-koeln.de Received September 12th, 2013; revised October 16th, 2013; accepted November 15th, 2013 Copyright © 2013 Otmar Bock, Benjamin Baak. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2013 are reserved for SCIRP and the owner of the intellectual property Otmar Bock, Benjamin Baak. All Copyright © 2013 are guarded by law and by SCIRP as a guardian. We evaluated the kinematics and dynamics of grasping in a typical laboratory situation (L) and in a more everyday-like situation (E), using right-handed subjects. Performance was compared when young subjects used their right versus left arm, and when young versus old subjects used their left arm. As in our previ- ous work, multiple differences emerged between parameter values in the two contexts, L and E. These context differences were, however, more pronounced for the left rather than for the right arm of young subjects, and more pronounced for the left arm of young rather than older subjects. We propose an expla- nation based on the differential involvement of the dorsal and ventral cortical processing stream in L and in E: The differential involvement would be accentuated for the left arm of young, but not for the left arm of older subjects. Keywords: Motor Control; Prehension; Context-Dependence; Sensorimotor Integration Introduction It has been suggested before that findings on motor perfor- mance, yielded in the laboratory, may not necessarily apply in everyday life (Chaytor & Schmitter-Edgecombe, 2003; In- gram & Wolpert, 2011). We have recently scrutinized this view by asking subjects to grasp and move a lever either in a typical laboratory context (L)—grasping was instructed, externally triggered, repetitive, and served no ultimate purpose—or in a more everyday-like context (E)—grasping was not explicitly instructed, self-initiated, embedded in complex behavior and had the ecologically valid purpose to earn money (Bock & Hagemann, 2010). Even though the mechanical constraints were identical in both contexts, movement kinematics and dy- namics widely differed. These differences could not be reduced to a single underlying cause since factor analysis yielded multi- ple orthogonal factors (Bock & Züll, 2013). This led us to con- clude that grasping is controlled by multiple functional modules which are differently sensitive to context. Further research revealed that context-sensitivity can’t be reduced to differences between L and E regarding movement speed, attention focusing or task complexity, since manipula- tions of those differences didn’t consistently change con- text-sensitivity (Steinberg & Bock, 2013c). However, we ob- served a consistent effect of personality traits: context-sensitiv- ity was accentuated in subjects who prefer slow, attentive and prudent processing (Steinberg & Bock, 2013b). We proposed that this processing style is characteristic for the ventral rather than the dorsal occipito-frontal stream in the human cortex (Goodale & Milner, 1992; Milner & Goodale, 1993). while the dorsal stream engages in quick automated reactions, the ventral stream specializes in slow and attention-demanding behavior (Buxbaum, Johnson-Frey, & Bartlett-Williams, 2005; Daprati & Sirigu, 2006; Rossetti & Pisella, 2002). Our interpretation thus links context-sensitivity to different cortical processing streams. The present study investigates whether context-sensitivity, observed previously for the dominant arm, holds equally for the non-dominant arm. It is well established that movement per- formance is not the same for both arms: the non-dominant arm controls intersegmental torques less well (Sainburg & Kalaka- nis, 2000), which shows poorer performance on tasks requiring high precision (Gonzalez, Ganel, & Goodale, 2006; Wing, Tur- ton, & Fraser, 1986; Woodworth, 1899), but better performance than the dominant arm on tasks requiring high speed (Annett, Annett, Hudson, & Turner, 1979; Carson, Chua, Goodman, By- blow, & Elliott, 1995; Elliott et al., 1993). It has been conclud- ed that the non-dominant arm is optimized for controlling limb posture, and the dominant arm for regulating limb trajectory (Sainburg, 2004). Such a specialization might reflect the pre- ferred arm use in bimanual activities: objects are typically held and stabilized by the non-dominant, and manipulated by the dominant hand (Grosskopf & Kuhtz-Buschbeck, 2006; Trevar- then, 2010). Arm specialization has also been linked to differ- ences in the underlying control principles, feedback control for the non-dominant versus preplanning for the dominant arm (Sain- burg & Kalakanis, 2000). Given these profound differences between arms, we reasoned that context-sensitivity might also be different. Specifically, we formulated two alternative hypo- theses: according to the first, the non-dominant arm depends more heavily on sensory feedback and therefore should be less susceptible to extraneous influences such as behavioral context.  O. BOCK, B. BAAK According to the second, the non-dominant arm is not well- practiced in manipulation tasks such as grasping and therefore should be more susceptible to extraneous influences such as behavioral context. We have shown before that context-sensitivity of grasping changes in old age; some parameters become more and other less context-sensitive, with no substantiable net change across all parameters (Bock & Steinberg, 2012). Again, these data have been yielded in the dominant arm. Since handedness is less pronounced in old age (Kalisch et al., 2006), we expected that any increase or decrease of context-sensitivity observed in the left arm of young subjects should be smaller in the left arm of seniors. To find out, the present study includes data from the non-dominant arm of elderly participants. Methods Participants Forty-eight young (24.3 ± 3.9 years) and thirty older subjects (71.8 ± 7.4 years) participated. All were right-handed, free of musculoskeletal impairments, diseases of the nervous system and visual deficits except for corrected vision by self-report, and lived independently in the community. None of them had par- ticipated in research on grasping or cognition within the last 12 months. An ethical approval for this study was given by the ins- titutional review board of the German Sport University Colog- ne, and all subjects signed an informed consent statement be- fore participating. Half of the young subjects were tested using their dominant (right) arm. The other half of the young and all older subjects were tested using their non-dominant (left) arm. Task and Procedure Experimental hardware and procedures were as in our previ- ous studies (Bock & Beurskens, 2010). Subjects sat at a table facing a 17 computer screen 67 cm ahead. A cylindrical lever of 4 cm length and 1.5 cm diameter was positioned 35 cm away from the front edge and 16 cm above the surface of the table 10 cm to the right of the screen or, for left arm testing, 10 cm to the left. The lever was covered by a hood from three sides, to ensure that subjects could only grasp it with the precision grip (thumb and index finger). The lever could slide 3.5 cm towards the subjects’ body midline along a rail (see Figure 1), where it met a mechanical stop. A displacement sensor (Burster® 8740) registered the lever’s position and a 6 df force transducer (ATI® Nano 17) registered the forces applied to the lever, both with a sampling rate of 250 Hz. A joystick was mounted 41 cm in front of the screen with its tip 12 cm above the table’s surface, such that its distance from the lever was 32 cm horizontally and 4 cm vertically. Six reflecting markers of 6 mm diameter were placed on thumb and index finger of the subjects’ grasping hand with double-sided adhesive tape, and two Vicon® MX- F20 3D high resolutions infrared cameras (sampling rate: 250 Hz, 1680 × 1280 pixels) registered their positions. In a laboratory task (L), the joystick was locked in its central position and subjects touched its tip with thumb and index fin- ger. At randomly varying intervals of 2 - 6 s, a green dot was displayed on the screen accompanied by a beep, prompting subjects to release the joystick and grasp the lever, to slide it towards them and back again, and then to return the hand to the joystick. In an everyday-like task (E), the joystick was un- locked and subjects were asked to play a computer game of C1 C2 S J L Figure 1. Schematic representation of the experimental set-up with joystick (J), lever (L), screen (S) and cameras (C1 & C2). chasing spiders on the screen with a joystick-driven cursor. A reward of .02 € was displayed near the right edge of the screen for each spider hit. Each game level terminated after 10 s, and subjects then had to collect their reward by moving the joystick to the center, grasping the lever, moving it towards them and back, and then returning their hand to the joystick. To keep the game motivating, speed and complexity of spider movement increased after every fifth level. No instructions were given on how to grasp the lever in task E, and subjects were not told that the purpose of their participation was to collect data on grasp- ing. In accordance with our earlier study (Bock & Steinberg, 2012), spider speed was 30% lower in older subjects than in young ones. To exclude carryover effects, each subject was engaged in only one of the tasks. Thus, 12 young subjects were tested in L and 12 in E using their left arm, 12 young subjects were tested in L and 12 in E using their right arm, and 15 older subjects were tested in L and 15 in E using their left arm. Each subject had 3 - 5 practice trials with the pertinent task and hand, to ensure that procedures were understood, and data were then collected for 20 grasping responses per subject. Note that both tasks used the same objects (joystick and lever) in the same location, and required the same hand and lever movements; they only differed with respect to their context: grasping was instructed, repetitive and served no ultimate purpose in L, but was uninstructed, part of complex behavior and had financial gain as purpose in E. Data Analysis Registered data were reduced by an interactive computer al- gorithm to 20 parameters representing the means of kinematic and dynamic landmarks across trials, and 20 parameters repre- senting the pertinent coefficients of variation. The additional parameter “Peaks” can’t be parsed into a mean and a CV. A definition of all parameters is provided in Table 1. Each pa- rameter was submitted to a two-way analysis of variance (ANOVA). Left and right arm performance of young subjects was compared with the between-factors Arm (left, right) and Open Access 999  O. BOCK, B. BAAK Open Access 1000 Table 1. Parameter definitions and the pertinent ANOVA outcomes*. ANOVA left/right ANOVA young/old Acronym Definition Task Arm Task * Arm Task Age Task * Age TT (s) Time from movement onset to lever contact (transport time) Mean CV 70.7*** 1.2 2.8 2.5 2.1 4.6* 91.7*** 4.7* 10.6*** 4.5* 0.8 0.0 Vmax (cm/s) Peak tangential hand velocity Mean CV 28.1*** 0.0 8.9** 0.8 1.0 3.7 15.2*** 4.1* 3.1 3.8 11.8** 3.5 Skew-T Ratio of deceleration time (Vmax to lever contact) and TT Mean CV 29.9*** 8.3** 0.0 0.0 3.3 0.1 7.8** 16.4*** 1.6 3.0 1.0 1.6 Detour-V (cm) Peak vertical distance of hand from a straight path Mean CV 27.1*** 7.1* 7.8** 0.2 1.4 0.0 22.4*** 0.9 1.8 3.0* 0.4 1.6 Transport component Detour-H (cm) Peak horizontal distance of hand from a straight path Mean CV 13.8*** 7.1* 11.1** 1.1 6.3* 3.4 22.0*** 0.0 5.4* 0.6 10.6** 0.3 GT (s) Time during which finger aperture changes (grasp time) Mean CV 79.5*** 4.5* 3.2 2.5 2.9 4.5* 90.0*** 13.6*** 7.6** 5.9* 0.1 1.8 PGA (cm) Peak 3D distance from thumb to index finger (peak grip aperture)Mean CV 6.1* 1.8 3.0 0.2 2.0 1.5 14.1*** 0.0 1.1 3.5 0.4 0.0 Peaks Proportion of multi-peaked aperture profiles 17.2*** 9.4** 2.0 12.3*** 4.9* 4.8* t(PGA) (s) Interval movement onset to PGA Mean CV 102.3*** 2.9 2.1 6.2* 2.1 0.5 98.1*** 11.3** 4.9* 4.9* 0.1 3.9 t(FGA) (s) Interval PGA to lever contact (final grip aperture) Mean CV 3.4 0.2 3.8 1.6 2.8 4.1* 8.3** 0.6 6.3* 5.9* 0.0 1.0 Skew-G Ratio of t(FGA) and GT Mean CV 89.3*** 1.0 1.3 1.1 0.0 1.7 40.9*** 5.5* 0.0 4.0 2.1 0.0 incli-start (˚) Hand inclination with respect to horizontal at movement onsetMean CV 0.0 1.7 0.5 1.4 1.7 0.0 2.9 0.1 2.2 1.5 0.3 0.7 incli-100 (˚) Hand inclination after 100 ms Mean CV 0.9 0.4 0.0 0.0 2.4 0.2 7.7** 0.4 0.1 1.9 0.5 0.3 incli-PGA (˚) Hand inclination at time of PGA Mean CV 4.3* 5.3* 1.5 1.2 4.7* 2.9 16.3*** 10.9** 6.9* 0.3 0.8 1.3 Grasp component incli-end (˚) Hand inclination at lever contact Mean CV 7.8** 7.2* 10.7** 2.9 3.9 6.7* 16.4*** 13.0*** 9.5** 0.1 1.8 1.1 Sync-start Interval onset of finger opening and of hand transport Mean CV 27.7*** 0.0 1.7 7.0* 1.6 11.2** 27.2*** 3.8 1.1 0.1 0.3 0.5 Coupling Sync-peak Interval t(PGA) and t(Vmax) Mean CV 79.8*** 0.4 1.7 8.9** 3.9 0.5 61.5*** 2.7 3.9 1.9 0.1 5.7* RT-lever (s) Interval lever contact and onset of lever motion (reaction time)Mean CV 11.1** 1.3 110.8*** 3.1 0.7 1.8 8.9** 0.2 7.9** 0.8 2.2 0.4 F-100 (N) Force compressing the lever 100 ms after lever contact Mean CV 25.6*** 7.2** 3.2 0.0 0.0 8.4** 3.5 0.4 1.3 4.3* 8.7** 17.4*** TQ-100 (N/mm) 3D lever torque 100 ms after lever contact Mean CV 25.2*** 9.6** 3.4 2.6 0.0 16.7*** 35.6*** 7.2** 1.0 19.9*** 5.7* 15.0*** Lever manipulation LT (s) Interval onset and end of lever motion (lever time) Mean CV 12.7*** 0.1 2.2 5.8* 1.5 2.2 12.2*** 0.1 0.6 2.8 3.3 2.1 Note: *Numbers are F-values with 1.44 degrees of freedom. *,** and ***Represent p < 0.05, p < 0.01 and p < 0.001, respectively. For each parameter, ANOVA for the means is presented above ANOVA for the CVs. Task (L, E). Left arm performance of young and older subjects was compared with the between-factors Age (young, older) and Task. Since the main effects of Task have already been ana- lysed in several earlier publications, the present work focuses on the other effects. Each parameter with a significant effect of Arm * Task was transformed by rh RHLHLH rh rhLHLH rh RH 1hLH rhRHLH1h 1EL E or LeLE or LEIE or LELe (1) where lrh, erh, llh and elh are parameter values of individual sub- jects participating with their right hand in task L or E, or with their left hand in task L or E, respectively, while LRH, ERH, LLH and ELH are the corresponding group means. Thus, large scores represent a stronger task-dependence of the right com- pared to the left hand. Significant effects of Age * Task were transformed accord- ingly. We then submitted the scores to factor analyses with varimax rotation, using the inclusion criterion F = 1. Results The right part of Table 1 summarizes the ANOVA outcome for young subjects using the left or right arm in L or E. As in our previous work (Bock & Steinberg, 2012; Bock & Züll, 2013; Steinberg & Bock, 2013a; Steinberg & Bock, 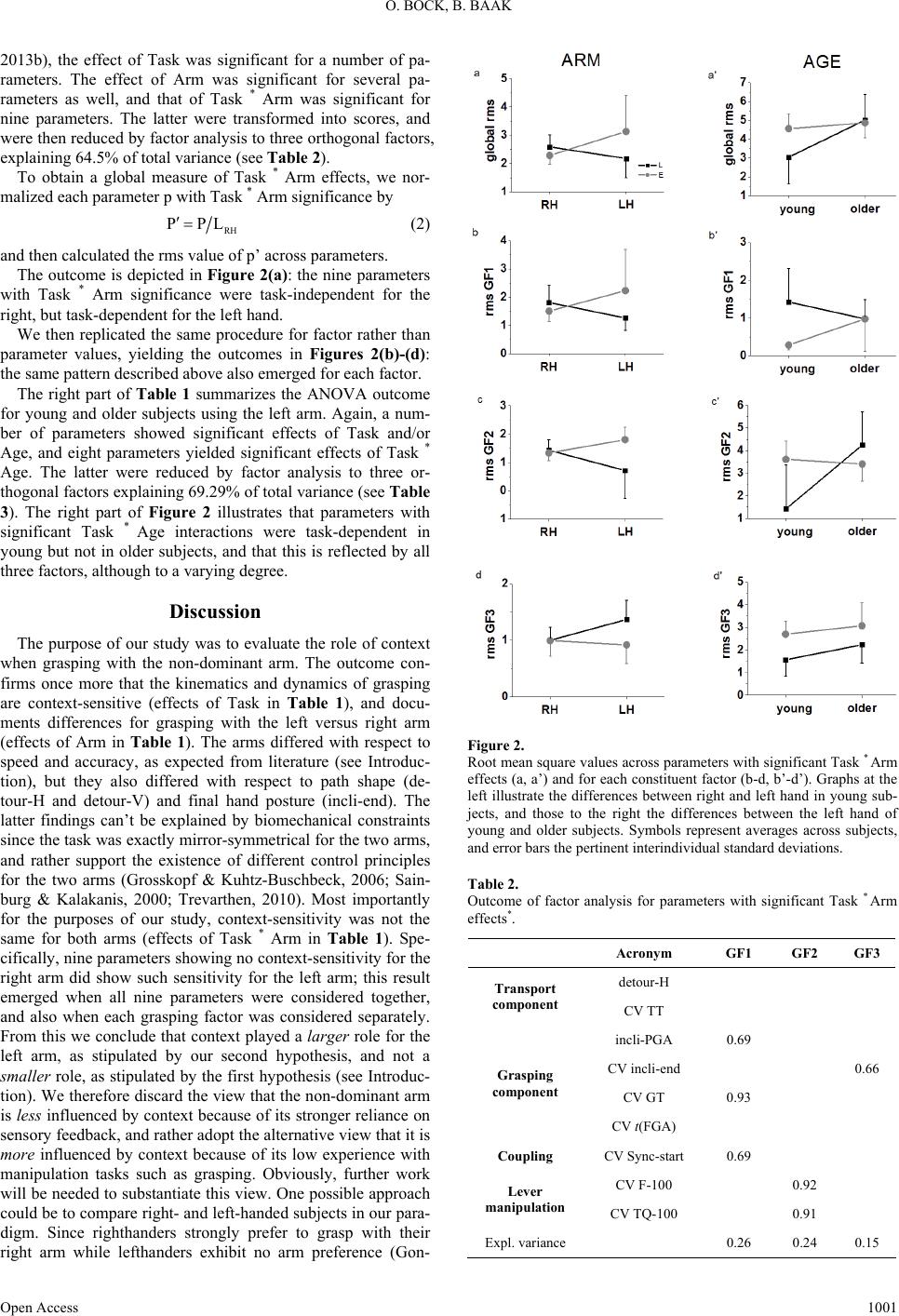 O. BOCK, B. BAAK 2013b), the effect of Task was significant for a number of pa- rameters. The effect of Arm was significant for several pa- rameters as well, and that of Task * Arm was significant for nine parameters. The latter were transformed into scores, and were then reduced by factor analysis to three orthogonal factors, explaining 64.5% of total variance (see Table 2). To obtain a global measure of Task * Arm effects, we nor- malized each parameter p with Task * Arm significance by RH PPL (2) and then calculated the rms value of p’ across parameters. The outcome is depicted in Figure 2(a): the nine parameters with Task * Arm significance were task-independent for the right, but task-dependent for the left hand. We then replicated the same procedure for factor rather than parameter values, yielding the outcomes in Figures 2(b)-(d): the same pattern described above also emerged for each factor. The right part of Table 1 summarizes the ANOVA outcome for young and older subjects using the left arm. Again, a num- ber of parameters showed significant effects of Task and/or Age, and eight parameters yielded significant effects of Task * Age. The latter were reduced by factor analysis to three or- thogonal factors explaining 69.29% of total variance (see Table 3). The right part of Figure 2 illustrates that parameters with significant Task * Age interactions were task-dependent in young but not in older subjects, and that this is reflected by all three factors, although to a varying degree. Discussion The purpose of our study was to evaluate the role of context when grasping with the non-dominant arm. The outcome con- firms once more that the kinematics and dynamics of grasping are context-sensitive (effects of Task in Table 1), and docu- ments differences for grasping with the left versus right arm (effects of Arm in Table 1). The arms differed with respect to speed and accuracy, as expected from literature (see Introduc- tion), but they also differed with respect to path shape (de- tour-H and detour-V) and final hand posture (incli-end). The latter findings can’t be explained by biomechanical constraints since the task was exactly mirror-symmetrical for the two arms, and rather support the existence of different control principles for the two arms (Grosskopf & Kuhtz-Buschbeck, 2006; Sain- burg & Kalakanis, 2000; Trevarthen, 2010). Most importantly for the purposes of our study, context-sensitivity was not the same for both arms (effects of Task * Arm in Table 1). Spe- cifically, nine parameters showing no context-sensitivity for the right arm did show such sensitivity for the left arm; this result emerged when all nine parameters were considered together, and also when each grasping factor was considered separately. From this we conclude that context played a larger role for the left arm, as stipulated by our second hypothesis, and not a smaller role, as stipulated by the first hypothesis (see Introduc- tion). We therefore discard the view that the non-dominant arm is less influenced by context because of its stronger reliance on sensory feedback, and rather adopt the alternative view that it is more influenced by context because of its low experience with manipulation tasks such as grasping. Obviously, further work will be needed to substantiate this view. One possible approach could be to compare right- and left-handed subjects in our para- digm. Since righthanders strongly prefer to grasp with their right arm while lefthanders exhibit no arm preference (Gon- Figure 2. Root mean square values across parameters with significant Task * Arm effects (a, a’) and for each constituent factor (b-d, b’-d’). Graphs at the left illustrate the differences between right and left hand in young sub- jects, and those to the right the differences between the left hand of young and older subjects. Symbols represent averages across subjects, and error bars the pertinent interindividual standard deviations. Table 2. Outcome of factor analysis for parameters with significant Task * Arm effects*. Acronym GF1 GF2 GF3 detour-H Transport component CV TT incli-PGA 0.69 CV incli-end 0.66 CV GT 0.93 Grasping component CV t(FGA) Coupling CV Sync-start 0.69 CV F-100 0.92 Lever manipulation CV TQ-100 0.91 Expl. variance 0.26 0.24 0.15 Open Access 1001  O. BOCK, B. BAAK Table 3. Outcome of factor analysis for parameters with significant Task * Age effects*. Acronym GF1 GF2 GF3 detour-H 0.77 Transport component Vmax Grasping component Peaks 0.78 Coupling CV Sync-peak F-100 0.86 TQ-100 0.90 CV F-100 0.89 Lever manipulation CV TQ-100 0.84 Expl. variance 0.25 0.24 0.20 Note: *Tables 2 & 3, Numbers are factor loadings, only values ≥ 0.6 are shown. The bottom row indicates the fraction of total variance explained by the respective factor. GF stands for grasping factor. zalez, Whitwell, Morrissey, Ganel, & Goodale, 2007a), the nondominant arm of lefthanders is experienced with manipula- tion tasks and their Task * Arm effects should therefore be less pronounced than in righthanders, if our second hypothesis is indeed correct. In our previous work, we have related context-sensitivity to the existence of two occipito-frontal processing streams in the human cortex: A dorsal stream is mainly concerned with fast automated reactions, and a ventral stream dealing with slow, attention-demanding behavior (Buxbaum et al., 2005; Goodale & Milner, 1992; Rossetti & Pisella, 2002). Since both streams are interconnected (Goodale & Westwood, 2004), processing of a given sensorimotor action may not be exclusively confined to one of the two streams, but may involve both of them in vary- ing degrees. Given these facts, we posit that young subjects using their right arm in L will preferentially engage the dorsal stream, since L requires externally triggered, stereotyped be- havior. In E, however, they will more strongly involve the ven- tral stream since E requires complex, volitional behavior. This view is illustrated in a simplified fashion by the top half of Figure 3. The bottom half of that figure illustrates how the neu- ral activation might change when young subjects use their left arm. Since arm is specialized for postural rather than voli- tional responses (see Introduction), its control circuitry might be well suited for the automated responses in L, but might re- quire particularly strong ventral activation for the volitional responses in E. Accordingly, Figure 3 shows no difference between left and right arm in L, but a shift towards the ventral stream for the left arm in E. Note that as a consequence, context sensitivity (i.e., the difference between L and E) is more pro- nounced for the left than for the right arm, as observed experi- mentally. Our data further show that grasping performance is affected by old age (effects of Age in Table 1). Elderly persons expect- edly differ from younger ones regarding movement duration and variability, but also regarding path shape and hand posture. Most importantly for the purposes of our study, context-sen- sitivity of the left arm differed between age groups (effects of Task * Age in Table 1): eight parameters showing context dif- ferences in young subjects showed smaller, null or even in- versed context differences in the elderly, with the net effect across all parameters being an absence of an appreciable con- text-sensitivity. In other words, the increase of context-speci- Figure 3. Schematic representation of assumed sensorimotor processing through the dorsal and the ventral stream in task L and E, when young subjects use their right versus left arm. ficity from the dominant to the non-dominant arm, as observed in young subjects, was attenuated if not absent in the elderly. This conforms to our expectation (see Introduction), according to which less pronounced handedness in old age is paralleled by less pronounced differences between the two arms regarding context-sensitivity. Referring back to Figure 3, one could argue that seniors grasping in E can’t increase the ventral contribution when using their left rather than their right arm, and their per- formance with the left arm therefore resembles that of young subjects using their right arm. Acknowledgements We thank Thomas Kesnerus for software development, the team of Hans-Martin Küsel-Feldker for hardware modifications and Malte Kraul & Annica Brosel for Figure preparation. Be- sides we wish to thank Fabian Steinberg for his support in de- signing the experiment and procedures. REFERENCES Annett, J., Annett, M., Hudson, P. T., & Turner, A. (1979). The control of movement in the preferred and non-preferred hands. The Quar- terly journal of experimental psych o logy, 31, 641-652. http://dx.doi.org/10.1080/14640747908400755 Bock, O., & Beurskens, R. (2010). Changes of locomotion in old age depend on task setting. Gait & Posture, 32, 645-649. http://dx.doi.org/10.1016/j.gaitpost.2010.09.009 Bock, O., & Hagemann, A. (2010). An experimental paradigm to com- pare motor performance under laboratory and under everyday-like conditions. Journal of Neuroscience Methods, 193, 24-28. http://dx.doi.org/10.1016/j.jneumeth.2010.08.005 Bock, O., & Steinberg, F. (2012). Age-related deficits of manual grasping in a laboratory versus in an everyday-like setting. Ageing Research, 3. http://dx.doi.org/10.4081/ar.2012.e7 Bock, O., & Züll, A. (2013). Characteristics of grasping movements in a laboratory and in an everyday-like context. Human Movement Sci- ence, 32, 249-256. http://dx.doi.org/10.1016/j.humov.2012.12.009 Buxbaum, L. J., Johnson-Frey, S. H., & Bartlett-Williams, M. (2005). Open Access 1002  O. BOCK, B. BAAK Open Access 1003 Deficient internal models for planning hand-object interactions in apraxia. Neuropsychology Review, 43, 917-929. http://dx.doi.org/10.1016/j.neuropsychologia.2004.09.006 Carson, R. G., Chua, R., Goodman, D. A., Byblow, W., & Elliott, D., (1995). The preparation of aiming movements. Brain and Cognition, 28, 133-154. http://dx.doi.org/10.1006/brcg.1995.1161 Chaytor, N., & Schmitter-Edgecombe, M. (2003). The ecological valid- ity of neuropsychological tests: A review of the literature on every- day cognitive skills. Neuropsychology Review, 13, 181-197. http://dx.doi.org/10.1023/B:NERV.0000009483.91468.fb Daprati, E., & Sirigu, A. (2006). How we interact with objects: Learn- ing from brain lesions/How we interact with objects: Learning from brain lesions. Trends in C o g n i t i ve Science, 10, 265-270. http://dx.doi.org/10.1016/j.tics.2006.04.005 Elliott, D., Roy, E. A., Goodman, D. A., Chua, R., Carson, R., Maraj, B., et al. (1993). Asymmetries in the preparation and control of ma- nual aiming movements. Canadian Journal of Experimental Psycho- logy, 47, 570-589. http://dx.doi.org/10.1037/h0078856 Fagard, J., & Lockman, J. J. (2005). The effect of task constraints on infants’ (bi)manual strategy for grasping and exploring objects. In- fant Behavior and Development, 28, 305-315. http://dx.doi.org/10.1016/j.infbeh.2005.05.005 Gonzalez, C. L. R., Ganel, T., & Goodale, M. A. (2006). Hemispheric specialization for the visual control of action is independent of handedness. Journal of Neu rophy siology, 95, 3496-3501. http://dx.doi.org/10.1152/jn.01187.2005 Gonzalez, C. L. R., Whitwell, R. L., Morrissey, B., Ganel, T., & Goodale, M. A. (2007). Left handedness does not extend to visually guided precision grasping. Experimental Brain Research, 182, 275- 279. http://dx.doi.org/10.1007/s00221-007-1090-1 Gonzalez, C. L. R., & Goodale, M. A. (2009). Hand preference for precision grasping predicts language lateralization. Neuropsycholgia, 47, 3182-3189. http://dx.doi.org/10.1016/j.neuropsychologia.2009.07.019 Goodale, M., & Milner, A. (1992). Separate visual pathways for per- ception and action. Trends in N e ur o sc i en c es , 15, 20-25. http://dx.doi.org/10.1016/0166-2236(92)90344-8 Goodale, M. A., & Westwood, D. A. (2004). An evolving view of duplex vision: Separate but interacting cortical pathways for percep- tion and action. Current Opinion in Ne urobiology, 14, 203-211. http://dx.doi.org/10.1016/j.conb.2004.03.002 Grosskopf, A., & Kuhtz-Buschbeck, J. P. (2006). Grasping with the left and right hand: A kinematic study. Experimental Brain Research, 168, 230-240. http://dx.doi.org/10.1007/s00221-005-0083-1 Ingram, J. N., & Wolpert, D. M. (2011). Naturalistic approaches to sensorimotor control. Progress in Brain Research, 191, 3-29. http://dx.doi.org/10.1016/B978-0-444-53752-2.00016-3 Kalisch, T., Wilimzig, C., Kleibel, N., Tegenthoff, M., Dinse, H. R., & Miall, C. (2006). Age-related attenuation of dominant hand superior- ity. PLoS ONE, 1, e90. http://dx.doi.org/10.1371/journal.pone.0000090 Milner, A., & Goodale, M. (1993). Visual pathways to perception and action. Progress in Brain Research, 95, 317-337. http://dx.doi.org/10.1016/S0079-6123(08)60379-9 Rossetti, Y., & Pisella, L. (2002). Several “Vision for Action” systems: A guide to dissociating and integrating dorsal and ventral functions (tutorial). In W. Prinz, & B. Hommel (Eds.), Attention and perform- ance; common mechanisms in perception and action (pp. 62-119). Oxford: Oxford University Press. Sainburg, R. L., & Schaefer, S. Y. (2004). Interlimb differences in con- trol of movement extent. Journal of Neurophysiology, 92, 1374-1383. http://dx.doi.org/10.1152/jn.00181.2004 Sainburg, R. L., & Kalakanis, D. (2000). Differences in control of limb dynamics during dominant and nondominant arm reaching. Journal of Neurophysiology, 83, 2661-2675. Steinberg, F., & Bock, O. (2013). Context dependence of manual grasping movements in near weightlessness. Aviation, Space, and Environmental Medicine, 84 , 467-472. http://dx.doi.org/10.3357/ASEM.3348.2013 Steinberg, F., & Bock, O. (2013). Influence of cognitive functions and behavioral context on grasping kinematics. Experimental Brain Re- search, 225, 387-397. http://dx.doi.org/10.1007/s00221-012-3379-y Steinberg, F., & Bock, O. (2013). The context dependence of grasping movements: An evaluation of possible reasons. Experimental Brain Research, 229, 587-594. http://dx.doi.org/10.1007/s00221-013-3618-x Trevarthen, C. (2010). Hemispheric specialization. In R. Terjung (Ed.), Comprehensive Physiology (pp. 1129-1190). Hoboken, NJ: John Wiley & Sons, Inc. Wing, A. M., Turton, A., & Fraser, C. (1986). Grasp size and accuracy of approach in reaching. Journal of Motor Behavior, 18, 245-260. http://dx.doi.org/10.1080/00222895.1986.10735380 Woodworth, R. (1899). The accuracy of voluntary movement. Psycho- logical Review, 3, 1-119.
|