M. GUANG
Copyright © 2013 SciRes. ENG
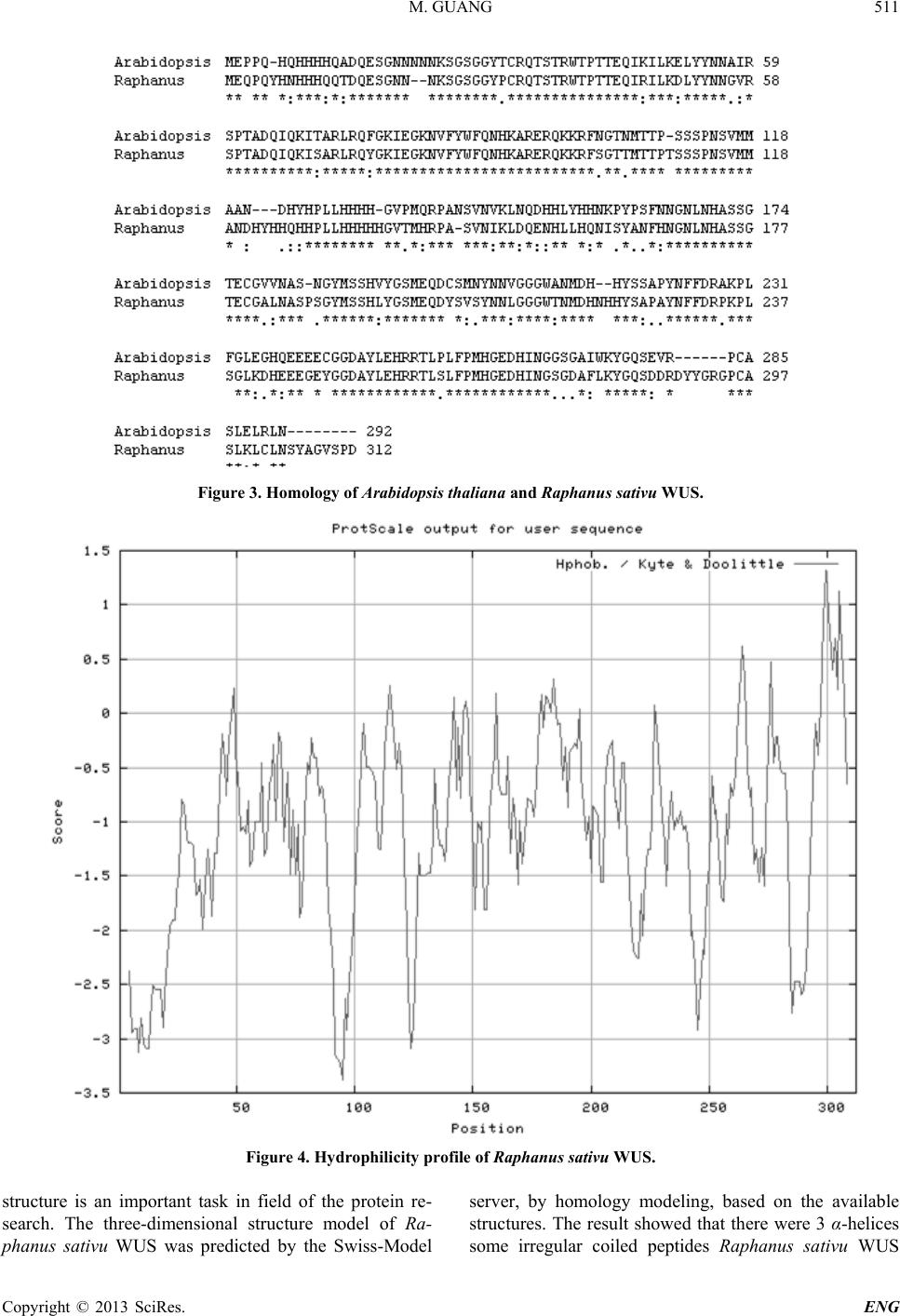
protein (Figure 5).
4. Disscusion
In silico cloning is a method developed in recent years
for functional gene identification by using genome and
EST database. Compared to the traditional methods, such
as molecular hybridization, sceening of genomic or
cDNA library, it is advanced for low cost, high effici e ncy,
easy operation, etc [7]. With more and more EST and
genome sequencing data were r eported, it would beco me
possible and feasible to isolate and identify the functional
genes by in silico cloning. Many successful examples
strongly support the fact that in silico cloning is abso-
lutely a feasible tool for gene cloning and presents some
advantages, compared to the traditional methods [8]. In
this study, the full cDNA of Raphanus sativu WUS was
obtained primarily by searching and splicing the EST
sequences. The structure and function were analyzed and
predicted using bioinformatics methods successfully. The
results revealed that it is a convenient technique for
cloning novel gene by searching EST database with ho-
mologous gene of model living things. To our knowledge,
it was the first report about cloning of Raphanus sativu
WUS cDNA with in silico cloning. This research
Figure 5. The predicted three-dimensional structure of Ra-
phanus sati vu WUS.
achievement will provide theory and reference for plant
stem research in Raphanus sativu.
5. Acknowledgements
The research is supported by fund of Hebei province
colleges and universities (No. Z2010159) and fund of
Hengshui city science and technology research (No.
09055Z).
REFERENCES
[1] I. Baurle and T. Laux, “Regulation of WUSCHEL Tran-
scription in the Stem Cell Niche of the Arabidopsis Shoot
Meristem,” Plant Cell, Vol. 17, 2005, pp. 2271-2280.
http://dx.doi.org/10.1105/tpc.105.032623
[2] Y. Y. Xu and K. Chong, “Progress in Research on Plant
Stem Cell Organizer Gene WUSCHEL,” (in Chinese) J.
Plant Phyoly. Mol. Bio., Vol. 31, 2005, pp. 461-468.
[3] G. Sena, X. Wang, H. Y. Liu, H. Hofhuis and K. D. Bir n-
baum, “Organ Regeneration Does Not Require a Func-
tional Stem Cell Niche in Plants,” Nature, Vol. 457, 2009,
pp. 1150-1153.
http://dx.doi.org/10.1038/nature07597
[4] T. Kondo, S. Sawa A. Kinoshita, S. Mizuno, T. Kakimoto,
H. Fukuda and Y. Sakagami, “A Plant Peptide Encoded
by CLV3 Identified by in Situ MALDI -TOF MS Analy-
sis,” Science, Vol. 313, 2006, pp. 845-848.
http://dx.doi.org/10.1126/science.1128439
[5] U. Brand, M. Grünewald, M. Hobe and R. Simon, “Reg-
ulation of CLV3 Expression by Two Homeobox Genes in
Arabidopsis,” Plant Physiology, Vol. 129, 2002, pp. 565-
575. http://dx.doi.org/10.1104/pp.001867
[6] T. Laux, F. X. Klaus, Mayer, B. Berger and G. Juren,
“The WUSCHEL Gene Is Required for Shoot and Floral
Meristem Integrity in Arabidopsis,” Development, Vol.
122, 1996, pp. 87-96.
[7] H. M. Zhang, M. G. Jiang and Y. J. Feng, “In Silico
Cloning of MgEno-1 cDNA from Magnaporthe grisea,”
China J. Bioinform at ics , 2005, pp. 57-61.
[8] H. Li, G. Y. Zhou, H. Y. Zhang, L. Lin and J. Liu, “In
Silico Cloning and Bioinformatic Analysis of PEPCK
Gene in Fusarium oxysporum,” African Journal of Bio-
technology, Vol. 9, 2010, pp. 1864-1870.