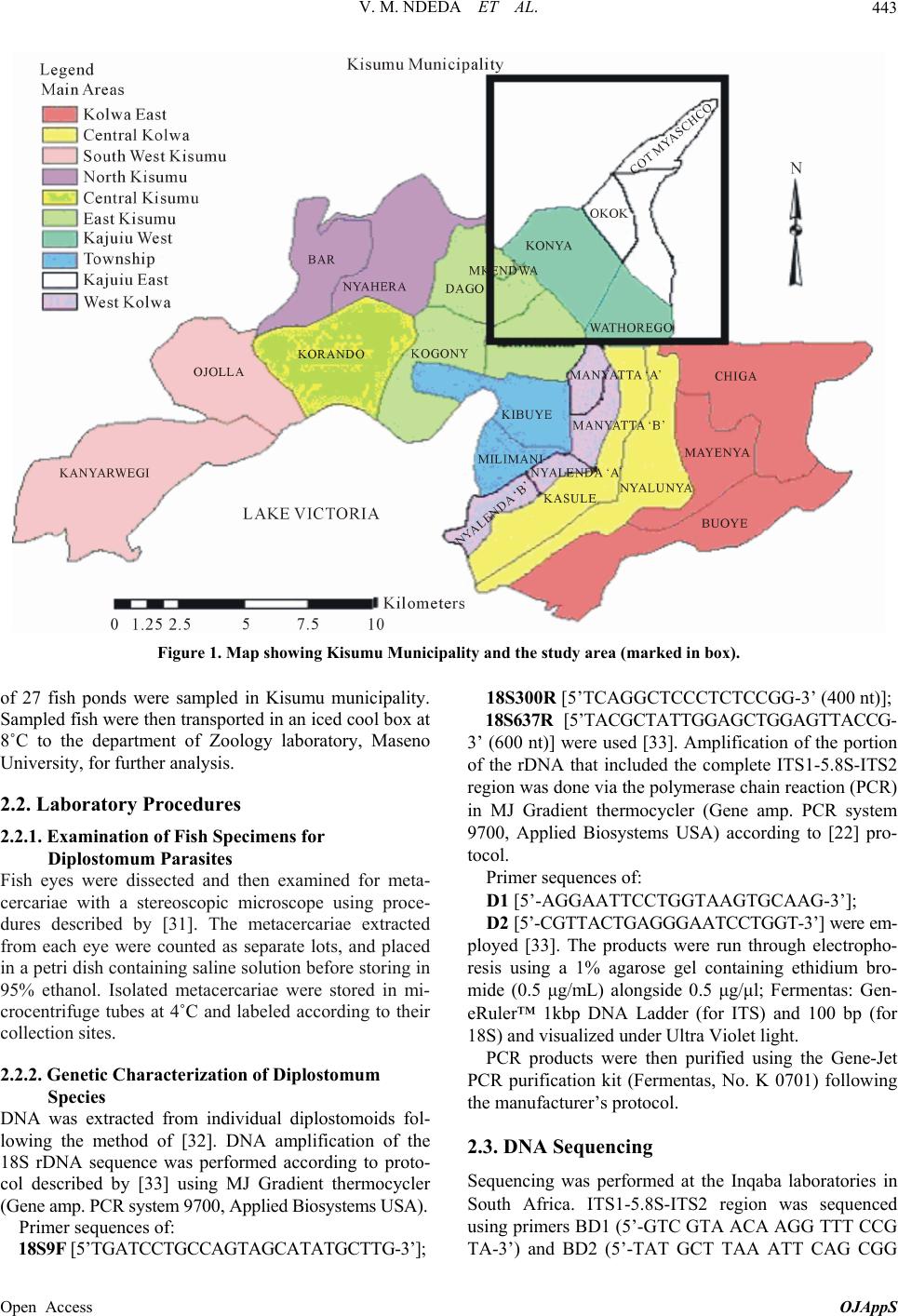
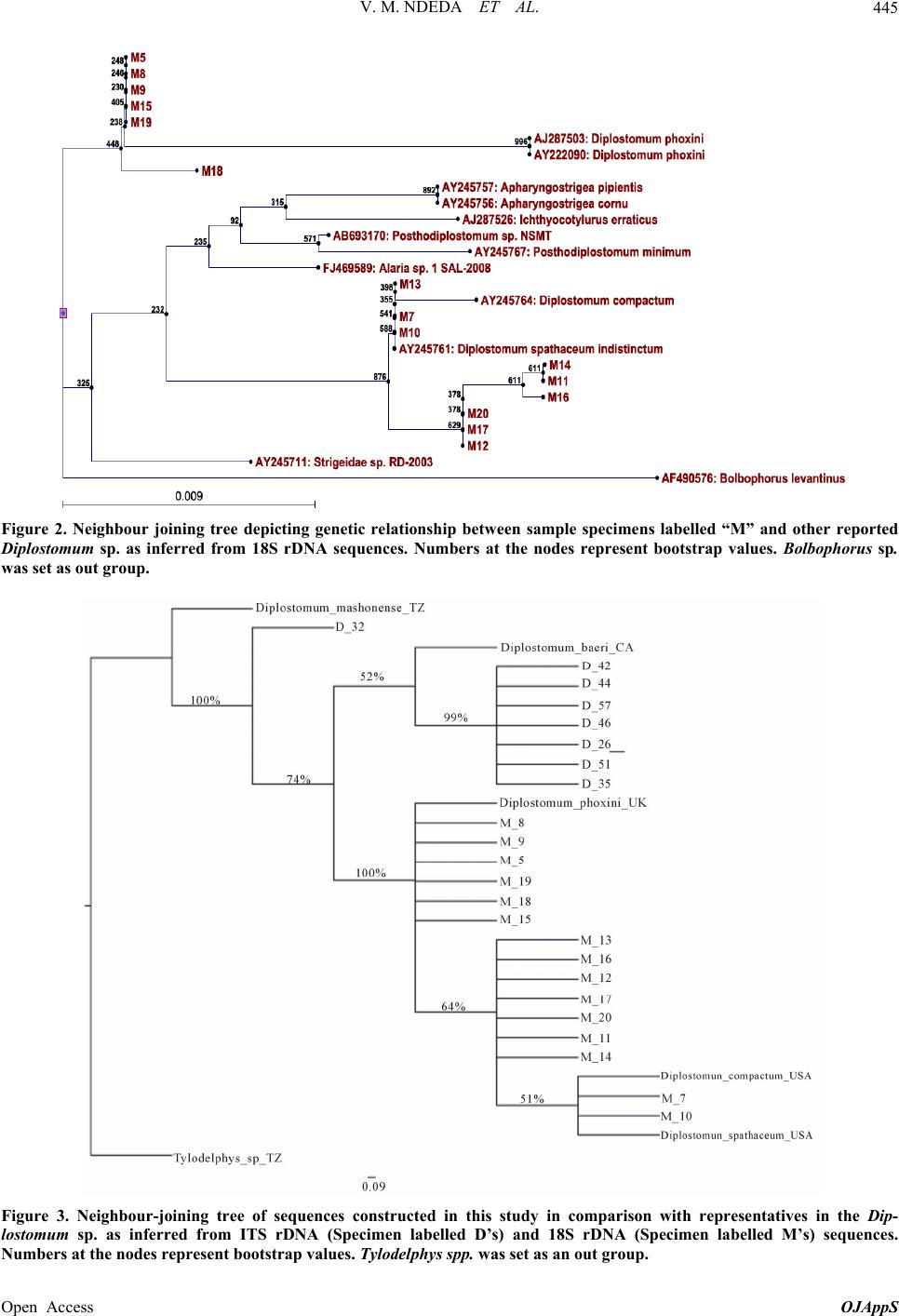
V. M. NDEDA ET AL. 447
Haematocheila (Temminck & Schlegel 1845) (Mugili-
dae),” Parasites and Vectors, Vol. 1, 2008, p. 48.
http://dx.doi.org/10.1186/1756-3305-1-48
[14] C. J. Mwita and G. Nkwengulila, “Determinants of th
Parasite Community of Clariid Fishes from
e
Lake Victoria,
Tanzania,” Journal of Helminthology, Vol. 82, No. 1,
2008, pp. 7-16.
http://dx.doi.org/10.1017/S0022149X07839745
[15] C. J. Mwita and
tionships of the Metazoan Parasites of the Clari
G. Nkwengulila, “Phylogenetic Rela-
id Fishes
s of Diplostomid Metacercariae (Digenea:
of Lake Victoria Inferred from Partial 18S rDNA Se-
quences,” Tanzanian Journal of Sciences, Vol. 37, 2011,
pp. 179-185.
[16] F. D. Chibwana and G. Nkwengulila, “Variation in the
Morphometric
Trematoda) Infecting the Catfish, Clarias gariepinus in
Tanzania,” Journal of Helminthology, Vol. 84, No. 1,
2010, pp. 61-70.
http://dx.doi.org/10.1017/S0022149X09990083
[17] F. Chibwana, I. Bla
G. Nkwengulila, T. Tomaš Scholz and A. Ko
sco-Costa, S. Georgieva, M. K Hosea,
stadinov
,
“First Insight into the Barcodes for African Diplostomids
(Digenea: Diplostomidae): Brain Parasites in Clarias
gariepinus (Siluriformes: Clariidae),” Infection Genetics
and Evolution, Vol. 17, 2013, pp. 62-70.
http://dx.doi.org/10.1016/j.meegid.2013.03.037
[18] G. K. Matolla, “Parasites of Tilapia (Oreo
cus) in Chepkoilel-Fish Farm of Moi University
chromis niloti-
, Eldore
t,
Kenya,” In: S. S. Drexler and H. Waidbacher, Eds., Pro-
ceedings of the 6th Framework Programme on BOMOSA
Projects: Integrating BOMOSA Cage Fish Farming Sys-
tem in Reservoirs, Ponds and Temporary Water Bodies in
Eastern Africa, Machakos, 2-4 September 2009, p. 28.
[19] E. M. Kembenya, G. K. Matolla, V. Ochieng’ and M.
Ngaira, “Diplostomum Parasites Affecting Oreochromis
niloticus in Chepkoilel Fish Farm and Two Dams in El-
doret-Kenya,” International Journal of Scientific & En-
gineering Research, Vol. 7, No. 3, 2012, pp. 1-7.
[20] G. N. Madanire-Moyo, W. J. Luus-Powell and P. A. S.
Olivier, “Ecology of Metazoan Parasites of Clariasgarie-
pinus (Osteichthyes: Clariidae) from the Nwanedi-Lu-
phephe Dams of the Limpopo River System, South Af-
rica,” African Zoology, Vol. 45, No. 2, 2010, pp. 233-243.
http://dx.doi.org/10.3377/004.045.0202
[21] J. April, R. L. Mayden, R. H. Hanner and L. Bernatcheza,
“Genetic Calibration of Species Diversity among North
America’s Freshwater Fishes,” PNAS, Vol. 108, No. 26,
2011, pp. 10602-10607.
http://dx.doi.org/10.1073/pnas.1016437108
[22] D. Gallazzo, E. S. Dayana
D. McLaughlin, “Molecular Systematics of
ndan, D. J. Marcogliese and J.
Some North
y and Specificity in Diplostomum
American Species of Diplostomum (Digenea) Based on
rDNA-Sequence Data and Comparisons with European
Congeners,” Canadian Journal of Zoology, Vol. 30, 2002,
pp. 2207-2217.
[23] S. Locke, J. D. Mclaughlin, S. Dayanandan and D. J.
Marcogliese, “Diversit
spp. Metacercariae in Freshwater Fishes Revealed by
Cytochrome c Oxidase I and Internal Transcribed Spacer
Sequences,” International Journal for Parasitology, Vol.
40, No. 3, 2010, pp. 333-343.
http://dx.doi.org/10.1016/j.ijpara.2009.08.012
[24] S. Locke, J. D. Mclaughlin and D. J. Marcogliese, “DNA
Barcodes Show Cryptic Diversity and a Potent
logical Basis for Host Specificity among Diplo
ial Physio-
stomoidea
(Platyhelminthes: Digenea) Parasitizing Freshwater Fishes
in the St. Lawrence River, Canada,” Molecular Ecology,
Vol. 19, No. 13, 2010, pp. 2813-2827.
http://dx.doi.org/10.1111/j.1365-294X.2010.04713.x
[25] K. Niewiadomska and Z. Laskowski, “Systematic Rela-
tionships among Six Species of Diplostomum nordma
1832 (Digenea) Based on Morphological and Molec
nn,
ular
Data,” Acta Parasitologica, Vol. 47, No. 1, 2002, pp. 20-28.
[26] C. Rellstab, K. Louhi, A. Karvonen and J. Jokela, “Ana-
lysis of Trematode Parasite Communities in Fish Eye
Lenses by Pyrosequencing of Naturally Pooled DNA,”
Infection, Genetics and Evolution, Vol. 11, No. 6, 2011,
pp. 1276-1286.
http://dx.doi.org/10.1016/j.meegid.2011.04.018
[27] Republic of Kenya, “Nyando District Development Plan,
Nairobi; Kenya,” 2002.
[28] National Environmental Management Authority [NEMA],
“Strategy for Flood Management in Lake Victoria Basin,
Kenya,” 2004.
[29] Lake Basin Development Authority, “Feasibility Study on
Kano Plain Irrigation Project,” Vol. 1, No. 71, 1992.
[30] W. W. Daniel, “Biostatistics: A Foundation for Analysis
in the Health Sciences,” John Wiley & Sons, New York,
1999.
[31] S. Yamaguti, “Synopsis of Digenetic Trematodes of Ver-
tebrates,” Keigaku Publishing, Tokyo, 1971.
[32] G. E. Truett, R. L. Heeger, A. A. Mynatt, J. A. Truett and
M. L. Warman, “Preparation of PCR-Quality Mouse Ge-
nomic DNA with Hot Sodium Hydroxide and Tris (Hot-
ne in
atoda: Dige-
SHOT),” BioTechniques, Vol. 29, 2000, pp. 52-54.
[33] A. Moszczynska, S. A. Locke, J. D. McLaughlin, D. J.
Marcogliese and T. J. Crease, “Development of Primers
for the Mitochondrial Cytochrome c Oxidase I Ge
Digenetic Trematodes Illustrates the Challenge of Bar-
coding Parasitic Helminthes,” Molecular Ecology Re-
sources, Vol. 9, No. S1, 2009, pp. 75-82.
[34] K. Luton, D. Walker and D. Blair, “Comparison of Ribo-
somal Internal Transcribed Spacers from Two Congeneric
Species of Flukes (Platyhelminthes: Trem
nea),” Molecular Biochemistry of Parasitology, Vol. 56,
No. 2, 1992, pp. 323-328.
http://dx.doi.org/10.1016/0166-6851(92)90181-I
[35] K. Tamura, J. Dudley, M. Nei and S. Kumar, “MEGA 4:
Molecular Evolutionary Genetics Analysis (MEGA) Soft-
ware Version 4.0,” Molecular Biology and Evolution, Vol.
24, No. 8, 2007, pp. 1596-1599.
http://dx.doi.org/10.1093/molbev/msm092
[36] M. S. Blouin, C. A. Yowell, C. H. Courtney and J. B.
Dame, “Host Movement and the Genetic
Populations of Parasitic Nematodes,” Gene Structure of
tics, Vol. 141,
No. 3, 1995, pp. 1007-1014.
[37] F. Prugnolle, A. Theron, J. P. Pointier, R. Jabbour-Zahab,
P. Jarne, P. Durand and T. de Meeus, “Dispersal in a
Open Access OJAppS