 Engineering, 2013, 5, 376-380 http://dx.doi.org/10.4236/eng.2013.510B076 Published Online October 2013 (http://www.scirp.org/journal/eng) Copyright © 2013 SciRes. ENG Isolation and Characterization of an L-Amino Acid Oxidase-Producing Marine Bacterium Zhiliang Yu1*, Hua Qiao1, Juanping Qiu1, Peiya Xu1, Peng Li2 1College of Biological and Environmental Engineering, Zhejiang University of Technology, Hangzhou, China 2College of Marine Science and Technology, Zhejiang Ocean University, Zhoushan, China Email: *zlyu@zjut.edu.cn Received 2013 ABSTRACT One marine bacterial strain, R3, has been newly isolated from the intertidal zone of Dinghai sea area. Measurements of a-keto acids and H2O2 existing in fermentation supernatant were carried out to show that R3 can produce L-amino acid oxidase (LAAO) with a broad substrate specificity. Physiological and biochemical analysis showed that it can grow great at the conditions with sodium chloride concentration of 1.5% - 3%, temperature of 15˚C - 35˚C and pH of 6 - 7. In addition, molecular identification of 16S rDNA was performed to show that R3 was proximal to Pseudoalterom onas spp. with the highest identity of 98.5% to Pseudoalteromonas ru bra. Therefore, it was designated as Pseudoalterom o- nas sp. R3. Further studies are required to arrive at a better understanding of this LAAO and secure an application. Keywords: LAAO; Pseudoalteromonas ; Physiological Property; Biochemical Property; Identification 1. Introduction L-amino acid oxidase (LAAO; EC 1.4.3.2) is dimeric flavoprotein, and each subunit contains a non-covalently bound FAD molecule as cofactor. It is able to catalyze the stereospecific oxidative deamination of L-amino ac- ids to a-keto acids, and H2O2. When H2O2 is not degraded by catalase, it can cause a decarboxylation of the a-keto acid to the corresponding carboxylic acid. Diverse studies have indicated that LAAOs have broad bioactivities such as inducing apoptosis [1], cytotoxicity [2,3], edema [4], hemolysis [5], hemorrhage [6], induc- ing or inhibiting platelet aggregation [7,8], parasite-kill- ing activity [9], and antimicrobial activity [10]. LAAO may act as defence or attack weapons via H2O2 [11] or as ideal molecular mechanisms for the acquisition of nitro- gen from diverse amino acid sources [12]. In addition, researches have showed that LAAO affects the relation of tumor cells with the immune system [13] and is in- volved in violacein synthesis [14]. So far, LAAO was found to be applied as catalysts in bio-transformation [15] and as part of biosensors in determination of the different forms (D- or L-) of free amino acids [16]. This enzyme has been widely found in nature includ- ing snake venoms, insect drugs [17], sea hare, algae [18] and terrestrial microorganisms [19]. In contrast, little is known about LAAO from marine microorganisms. The objec tive of th is study is to isolate and chara cterize LAAO- producing marine microorganism. We believe that this study lays the foundation for further investigation on en- zymatic properties, structure, biological function and ap- plicat i on of L A A Os f r om marine microorganisms. 2. Materials and Methods 2.1. Sample Collection and Isolation of LAAO-Producing Marine Microorganism The intertidal zone sludge samples (30.03˚N, 122.11˚E) were collected from different locations at the Dinghai sea area. From each location, sample was collected at 50 to 100 cm depth under the sea surface. These samples were placed in special pre-sterilized plastic bottles and brought to the laboratory in aseptic co ndition. Then, 10 g of each sludge sample was subjected to 90 mL of sterilized dis- tilled water and serially diluted (up to 10-6 dilution). After dilution, about 100 µL of each diluted sample was plated on different agar medium including PDA (potato 200 g/L, sucrose 10 g/L, sea salt 30 g/L), Gause’s Me- dium NO.1 (soluble starch 20 g/L, NaCl 0.5 g/L, K2HPO4 0.5 g/L, FeSO4 0.01 g/L, MgSO4∙7H2O 0.5 g/L, KNO3 1 g/L, sea salt 30 g/L; pH 7.2-7.4), and MM me- dium (yeast extract 3 g/L, peptone 5 g/L, sea salt 30 g/L; pH 7.2 - 7.4), and separately incubated at 28˚C and 25˚C for 2 - 7 days, as necessary. The isolated colonies were purified by streak-plate technique. LAAO activity was determined by measuring its fer- mentation products includ ing H2O2 and a-keto acids. The *Corresponding a uthor.  Z. L. YU ET AL. Copyright © 2013 SciRes. ENG 377 production of H2O2 was measured by using Amplex Red Hydrogen Peroxide/Peroxidase Assay kit (Invitrogen, USA). The production of a-keto acids were spectrophotometri- cally measured based on hydrazine assay, according to Len Sikora’s me t hod [20]. 2.2. Sodium Chloride Tolerance and Cultural Condition Different concentrations of sodium chloride (0%, 1.5%, 3%, 4.5%, 6%, 7.5%, 9%, 10.5% and 12%) were added to the MM medium (without sea salt). The seed of iso- lated LAAO-producer was planted into different MM medium and incubated at 25˚C, 160 rpm for 24 hours, and salt tolerance was tested. The growth of the LAAO- producer on MM medium incubated at different temper- ature (5˚C, 15˚C, 25˚C, 35˚C and 45˚C) and at different pH (3, 4, 5, 6, 7, 8, 9, 10, 11 and 12) was also investi- gated to determine its growth range of temperature and pH. 2.3. Physiological and Biochemical Characteriz ation The ability of the isolate to utilize various carbon and n i- trogen sources, and other physiological and biochemical properties were studied by the method recommended in “The Manual of Systematic Methods of Determinative Bacterial”, with minor modification by adding 3% so- dium chloride to each medium. 2.4. Sequencing Analysis The genomic DNA was isolated by bacterial genomic extraction method (EDTA treatment method). PCR reac- tions were performed in 50 µL containing 37 μL ddH2O, 5 μL 10 × Easy Taq buffer, 4 μL 2.5 mM dNTPs, 100 nM primer 27F (5’-GAGTTTGATC CTGGCTCAG-3’), 100 nM primer 1527R (5’-AGAAAGGAGG T GATCC- AGCC-3’), 1 ng genomic DNA, and 1 U Taq DNA po- lymerase with denaturation at 94˚C for 5 minutes fol- lowed by 30 cycles of 1 minute at 94˚C, 50 seconds at 55˚C, 90 seconds at 72˚C and a final 10-minutes exten- sion at 72˚C. At the end of reaction, PCR product was cooled to 4˚C to await further use. After size confirma- tion on 1.0% agarose gel, the PCR DNA product was sent to Sangon Biotech (Shanghai) Co. Ltd for sequenc- ing of 16 S rDNA. After sequencing, the similarity and homology of partial 16S rDNA were online analyzed us- ing BLAST search via NCBI. 3. Results 3.1. Isolation of LAAO-Producing Strain A total of 157 pure isolates were obtained from the inter- tidal zone of Dinghai sea area located in Zhoushan, Zhe- jiang province, China. Three domains including 32 acti- nomycetes, 51 fungus and 74 bacteria were morphologi- cally characterized. Out of 157 isolates subjected to screen- ing process, one isolate (R3) showed the capability to produce LA AO based on the below re sults. LAAO is able to catalyze the stereospecific oxidative deamination of L-amino acid to a-keto acid. As carbonyl derivative, a-keto acid can be easily measured by sensi- tive method using 2,4-dinitrophenylhydrazine (DNP) which can react with carbonyl group to generate dinitro-ph e - nylhydrazone with a brown-red color and characteristic absorbance maxima at 520 nm. As shown in Figure 1(c), R3 fermentation supernatant without L-amino acid sub- strate showed flaxen and OD520 was very low. Similarly, a mixture of R3 fermentation supernatant treated at 95˚C for 5 minutes and substrate L-Phe (Figure 1(d)) also showed flaxen with low OD520 value. In contrast, me- dium with L-Phenylpyruvic acid (Figure 1(a)) and R3 fermentation supernatants with L-Phe (Figure 1(b)) both had brown-red color with high OD520. All these results indicate that R3 with L-amino acid substrate can generate a-keto acid. Therefore, R3 can produce LAAO. LAAO is able to catalyze L-amino acid to release H2O2 which can be detected using Amplex Red Hydro- gen Peroxide/Peroxidase Assay kit. Our results showed that, for either R3 fermentation supernatant without sub- strate L-Met or R3 fermentation supernatant treated at 95˚C for 10 minutes followed by addition of L-Met, no H2O2 was detected. In contrast, huge amount of H2O2 (1.84 mmol/L) was detected in R3 fermentation superna- tant with L-Met, indicating that R3 can use L-Met to re- lease H2O2. Therefore, R3 can produce LAAO. 3.2. Morphological Characteristics On MM media plate, R3 displayed a red color and round form with smooth. Based on the Gram-staining, R3 was identified as a gram-negative strain with the size of about 3.15 μm × 1.05 μm (15 × 100). (a) (b) (c) (d) Figure 1. Detection of a-keto acids. (a) Positive control: Phe- nylpyruvi c acid with MM medium (OD520 0.202); (b) L-Phe with R3 (OD520 0.145); (c) R3 without L-Phe (OD520 0.038); (d) Treatment of B3 at 95˚C for 10 minutes followed by adding of L-Phe (OD520 0.051). 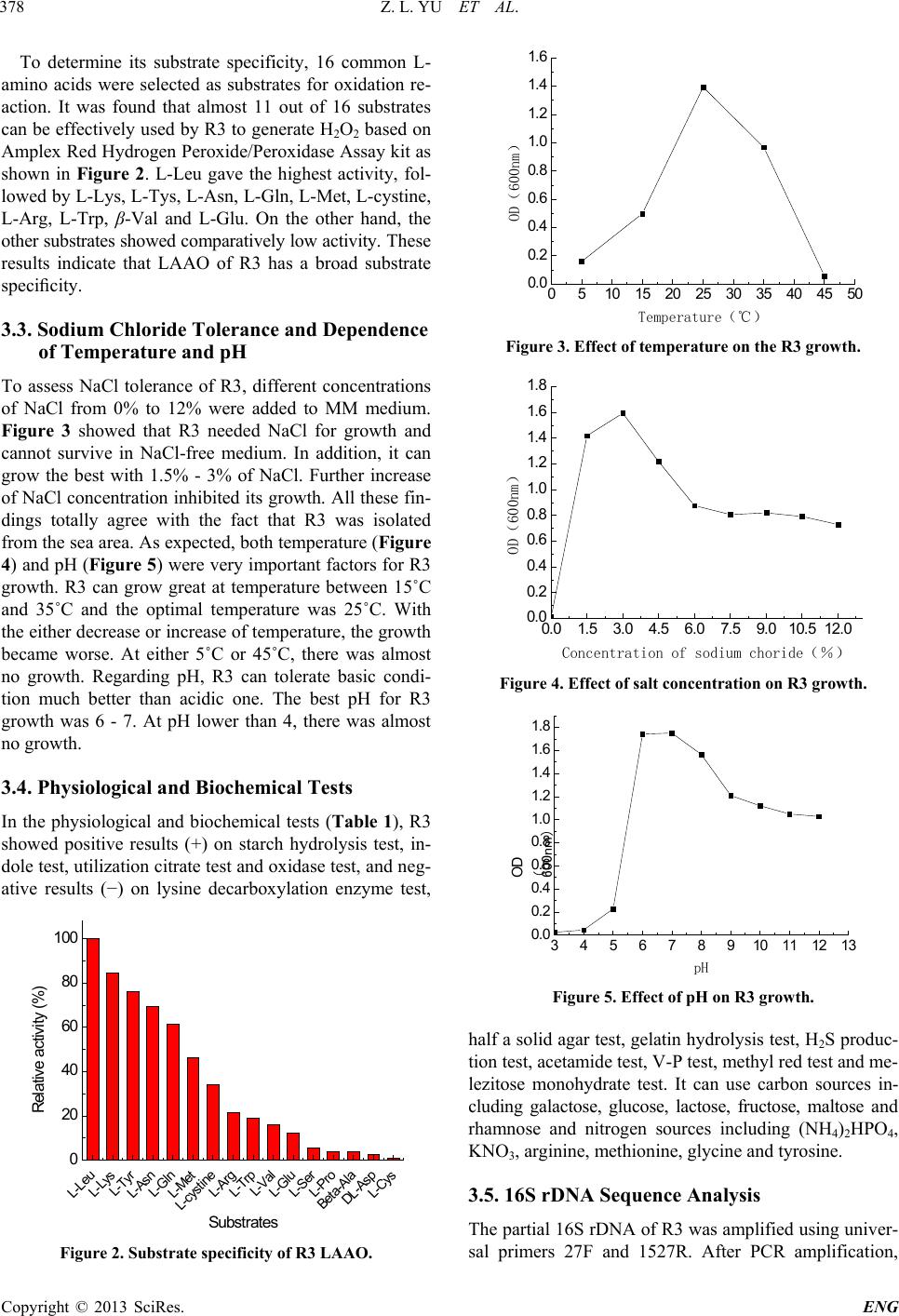 Z. L. YU ET AL. Copyright © 2013 SciRes. ENG To determine its substrate specificity, 16 common L- amino acids were selected as substrates for oxidation re- action. It was found that almost 11 out of 16 substrates can be effectively used by R3 to generate H2O2 based on Amplex Red Hydrogen Peroxide/Peroxidase Assay kit as shown in Figure 2. L-Leu gave the highest activity, fol- lowed by L-Lys, L-Tys, L-Asn, L-Gln, L-Met, L-cystine, L-Arg, L-Trp, β-Val and L-Glu. On the other hand, the other substrates showed comparatively low activity. These results indicate that LAAO of R3 has a broad substrate specificity. 3.3. Sodium Chloride Tolerance and Dependence of Temperature and pH To assess NaCl tolerance of R3, different concentrations of NaCl from 0% to 12% were added to MM medium. Figure 3 showed that R3 needed NaCl for growth and cannot survive in NaCl-free medium. In addition, it can grow the best with 1.5% - 3% of NaCl. Further increase of NaCl concentration inhib ited its growth. All these fin- dings totally agree with the fact that R3 was isolated from the sea area. As expected, both temperature (Figure 4) and pH (Figure 5) were very important factors for R3 growth. R3 can grow great at temperature between 15˚C and 35˚C and the optimal temperature was 25˚C. With the either decrease or increase of temperature, the growth became worse. At either 5˚C or 45˚C, there was almost no growth. Regarding pH, R3 can tolerate basic condi- tion much better than acidic one. The best pH for R3 growth was 6 - 7. At pH lower than 4, there was almost no growth. 3.4. Physiological and Biochemical Tests In the physiological and biochemical tests (Table 1), R3 showed positive results (+) on starch hydrolysis test, in- dole test, utilization citr ate test and oxidase test, and neg- ative results (−) on lysine decarboxylation enzyme test, Figure 2. Substrate specificity of R3 LAAO. Figure 3. Effect of temperature on the R3 growth. Figure 4. Effect of salt concentration on R3 growth. Figure 5. Effect of pH on R3 growth. half a solid agar test, gelatin hydrolysis test, H2S produc- tion test, acetamide test, V-P test, methyl red test and me- lezitose monohydrate test. It can use carbon sources in- cluding galactose, glucose, lactose, fructose, maltose and rhamnose and nitrogen sources including (NH4)2HPO4, KNO3, arginine, methionine, glycine and tyrosine. 3.5. 16S rDNA Sequence Analysis The partial 16S rDNA of R3 was amplified using univ er- sal primers 27F and 1527R. After PCR amplification, L-Leu L-Lys L-T yr L-Asn L-Gln L-Met L-c ystine L-Arg L-Trp L-Val L-Glu L-Ser L-Pro Beta-Ala DL-Asp L-Cys 0 20 40 60 80 100 Relative activity (%) Substrates 05 10 15 20 25 30 35 40 45 50 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 OD(600nm) Temperature(℃) 0.0 1.5 3.0 4.5 6.0 7.5 9.010.512.0 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 OD(600nm) Concentration of sodium choride(%) 345678910 11 12 13 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 OD ( 600nm) pH  Z. L. YU ET AL. Copyright © 2013 SciRes. ENG 379 Table 1. Physiological and biochemical characteristics of the R3 isolate. Ite ms R3 Carbon Sources R3 Nitrogen Sources R3 Lysine decarboxylation enzyme − Inositol − (NH4)2HPO4 + Half a solid agar − Sorbitol − KNO3 + Gelatin hydrolysis − Mannitol − Arginine + Starch hydrolysis + Glucose + Methionine + H2S production − Sucrose − Phenylalanine − Indole test + Rhamnose + Glycine + Methyl red test − Lactose + Tyrosine + V-P test − Fructose + Utilization citrate + Maltose + Acetamide − Galactose + Melezitose monohydrate − Oxidase test + around 1.4 Kb fragment was successfully obtained (data not shown) and sequenced. Then the Blast search via NCBI was carried out to evaluate the similarity and ho- mology of R3 with other organisms. It was found that R3 had very high homology with Pseudoalteromonas spp., with the highest identity of 98.5% to Pseudoalteromonas rubra. Therefore, we designated it as Pseudoalteromonas sp. R3. 4. Discussion LAAOs form a family of proteins with various enzymatic properties, structure and biological function. Extensive studies indicate that LAAOs have promising biotechno- logical and medical applications. This enzyme is widely distributed in nature including snake venoms, insect drugs, sea hare, fungi, bacteria and algae. Unlike snake venom LAAOs which have been widely and deeply investigated to show broad bioactivities such as apoptosis, cytotoxic- ity, edema, hemolysis, hemorrhage, platelet aggregation, parasite-killing activity and antimicrobial activity, non- snake venom LAAOs need to be further studied and their functional role and application remain to be revealed. Especially, very little is known about LAAOs from ma- rine microorganism. In this study, we successfully iso- lated an LAAO-producing marine bacterium from inter- tidal zone of Dinghai sea area. Based on physiological and biochemical tests together with molecular ana lysis, it was designated as Pseudoalteromonas sp. R3. To arrive at a better understanding and address the commercializa- tion of LAAO from this isolate, future works are planned to clone the gene coding for this Pseudoalteromonas sp. R3 LAAO and to study its structure, biological and phy- siological roles, relationship between function and struc- ture, and mechanism of transcription in vivo. 5. Acknowledgements This work was sup ported by Natural Science F oundation of Zhejiang Province, China (Y5100153), Welfare Tech- nology Applied Research Project of Zhejiang Province, China (2011C23007), and Natural Science Foundation of ZJUT (2010021 3) to Z.L. Yu. REFERENCES [1] S. Torii, M. Naito and T. Tsuruo, “Apoxin I, a Novel Apoptosis-Inducing Factor with L-Amino Acid Oxidase Activity Purified from Western Diamondback Rattlesnake Venom,” Journal of Biological Chemistry, Vol. 272, No. 14, 1997, pp. 9539-9542. http://dx.doi.org/10.1074/jbc.272.14.9539 [2] G. B. Naumanna, L. F. Silvaa, L. Silvaa, G. Faria, M. Ri- chardson, K. Evangelista, et al., “Cy totoxicity and Inhibi- tion of Platelet Aggregation Caused by an L-Amino Acid Oxidase from Bothrops leucurus Venom,” Bi ochimica et Biophysica Acta, Vol. 1810, No. 7, 2011, pp. 683-694. http://dx.doi.org/10.1016/j.bbagen.2011.04.003 [3] M. Y. Ahn, B. M. Lee and Y. S. Kim, “Characterization and Cytotoxicity of L-Amino Acid Oxidase from the Ve- nom of King Cobra (Ophiophagus hannah),” The Inter- national Journal of Biochemistry & Cell Biology, Vol. 29, No. 6, 1997, pp. 911-919. http://dx.doi.org/10.1016/S1357-2725(97)00024-1 [4] R. G. Stábeli, S. Marcussi, G. B. Carlos, R. C. Pietro, H. S. Selistre -de-Araújo, J. R. Giglio, et al., “Platelet Aggre- gation and Antibacterial Effects of an L-Amino Acid Oxi- dase Purifie d from Bothrops alternatus Snake Venom,” Bioorganic & Medicinal Chemistry, Vol. 12, No. 11, 2004, pp. 2881-2886. http://dx.doi.org/10.1016/j.bmc.2004.03.049 [5] Y. Abe, Y. Shimoyama, H. Munakata, J. Ito, N. Nagata and K. Ohtsuki, “Characterization of an Apoptosis-Induc- ing Factor in Habu Snake Venom as a Glycyrrhizin (GL)- Binding Protein Potently Inhibited by GL in Vitro,” Bio- logical & Pharmaceutical Bulletin, Vol. 21, No. 9, 1998, pp. 924-927. http://dx.doi.org/10.1248/bpb.21.924 [6] D. H. Souza, L. M. Eugenio, J. E. Fletcher, M. S. Jiang, R. C. Garratt, G. Oliva, et al., “Isolation and Structural Cha- racterization of a Cytotoxic L-Amino Acid Oxidase from Agkistrodon contortrix laticinctus Snake Venom: Prelim- inary Crystallographic Data,” Archives of Biochemistry  Z. L. YU ET AL. Copyright © 2013 SciRes. ENG and Biophysics, Vol. 368, No. 2, 1999, pp. 285-290. http://dx.doi.org/10.1006/abbi.1999.1287 [7] H. Takatsuka, Y. Sakurai, A. Yoshioka, T. Kokubo, Y. Usami, M. Suzuki, et al., “Molecular Characterization of L-Amino Acid Oxidase from Agkistrodon halys blomhof- fii with Special Reference to Platelet Aggregation,” Bio- chimica et Biophysica Acta-Protein Structure and Mole- cular Enzymology, Vol. 1544, No. 1-2, 2001, pp. 267-277. http://dx.doi.org/10.1016/S0167-4838(00)00229-6 [8] S. A. Alib, S. Stoeva b, A. Abbasi, J. M. Alam, R. Kayed, M. Faigle, et al. “Isolation, Structural, and Functio nal Cha- racterization of an Apoptosis-Inducing L-Amino Acid Oxidase from Leaf-Nosed Viper (Eristocophis macmaho- ni) Snake Venom,” Archives of Biochemistry and Biophy- sics, Vol. 384, No. 2, 2000, pp. 216-226. http://dx.doi.org/10.1006/abbi.2000.2130 [9] P. Deolindo, A. S. Teixeira-Ferreira, R. A. DaMatta and E. W. Alves, “L-Amino Acid Oxidase Activity Present in Fractions of Bothrops jararaca Venom Is Responsible for the Induction of Programmed Cell Death in Trypanosoma cruzi,” Toxicon, Vol. 56, No. 6, 2010, pp. 944-955. http://dx.doi.org/10.1016/j.toxicon.2010.06.019 [10] B. G. Stilesa, F. W. Sextona and S. A. Weinstein, “Anti- bacterial Effects of Different Snake Venoms: Purification and Characterization of Antibacterial Proteins from Pseu- dechis australis (Australian King Brown or Mulga Snake) Venom,” Toxicon, Vol. 29, No. 9, 1991, pp. 1129-1141. http://dx.doi.org/10.1016/0041-0101(91)90210-I [11] Y. Hsiuchin, M. J. Paul, K. Ko-Chun, M. Kamio, M. W. Germann, C. D. Derby and P. C. Tai, “Cloning, Characte- rization and Expression of Escapin, a Broadly Antimicro- bial FAD-Containing L-Amino Acid Oxidase from Ink of the Sea Hare Aplysia californica,” The Journal of Experi- mental Biology, Vol. 208, No. pt 18, 2005, pp. 3609-3622. http://dx.doi.org/10.1242/jeb.01795 [12] M. A. Davis, M. C. Askin and M. J. Hynes, “Amino Acid Catabolism by an Area-Regulated Gene Encoding an L- Amino Acid Oxidase with Broad Substrate Specificity in Aspergillus nidulans,” Applied and Environmental Micro- biology, Vol. 71, No. 7, 2005, pp. 3551-3555. http://dx.doi.org/10.1128/AEM.71.7.3551-3555.2005 [13] A. Carbonnelle-Puscian, C. Copie-Bergman, M. Baia, N. Martin-Garcia, Y. Allory, C. Haioun, et al. “The Novel Immunosuppressive Enzyme IL4I1 Is Expressed by Neo- plastic Cells of Several B-Cell Lymphomas and by Tu- mor-Associated Macrophages,” Leukemia, Vol. 23, No. 5, 2009, pp. 952-960. http://dx.doi.org/10.1038/leu.2008.380 [14] R. Genet, P. H. Benetti, A. Hammadi and A. Menez, “L- Tryptophan 2’,3’-Oxidase from Chromobacterium viola- ceum. Substrate Specificity and Mechani stic Implications,” The Journal of Biological Chemistry, Vol. 270, No. 40, 1995, pp. 23540-23545. http://dx.doi.org/10.1074/jbc.270.40.23540 [15] R. L. Hanson, K. S. Bembenek, R. N. Patel and L. J. Szar- ka, “Transformation of N-Epsilon-CBZ-L-Lysine to CBT- L-Oxylysine Using L-Amino Acid Oxidas e from Provi- dencia Alcalifaciens and L-2-Hydroxy-Iso-Caproate De- hydrogenase from Lactobacillus confuses,” Applied Mi- crobiology and Biotechnology, Vol. 37, No. 5, 1992, pp. 599-603. http://dx.doi.org/10.1007/BF00240733 [16] M. Varadi, N. Adanyi, E. E. Szabo and N. Trummer, “De- termination of the Ratio of D-and L-Amino Acids in Brewing by an Immobili ze Amino Acid Oxida se Enzyme Reactor Coupled to Amperometric Detection,” Biosensors & Bioelectronics, Vol. 14, No. 3, 1999, pp. 335-340. http://dx.doi.org/10.1016/S0956-5663(98)00130-4 [17] M. Y. Ahn, K. S. Ryu, Y. W. Lee and Y. S. Ki m, “Cyto- toxicity and L-Amino Acid Oxidase Activity of Crude Insect Drugs,” Archives of Pharmacal Research, Vol. 23, No. 5, 2000, pp. 477-481. http://dx.doi.org/10.1007/BF02976576 [18] K. Ito, K. Hori and K. Miyazawa, “Purification and Some Properties of L-Amino Acid Oxidase from Amphiroa crassissima Yendo,” Hydrobiologia, Vol. 152, 1987, pp. 563-569. http://dx.doi.org/10.1007/BF00046183 [19] G. M. Brearley, C. P. Price, T. Atkinson and P. M. Ham- mond, “Purification and Partial Characterisation of a Broad- Range L-Amino Acid Oxidase from Bacillus carotarum 2Pfa Isolated from Soil,” Applied Microbiology and Bio- technology, Vol. 41, No. 6, 1994, pp. 670-676. http://dx.doi.org/10.1007/BF00167283 [20] L. Sikora and G. A. Marzluf, “Regulation of L-Amino Acid Oxidase and of D-Amino Acid Oxidase in Neuros- pora crassa,” Molecular & General Genetics, Vol. 186, No. 1, 1982, pp. 33-39. http://dx.doi.org/10.1007/BF00422908
|