J. MONASELIDZE ET AL. 747
pable to wind the double helix and cause narrowing of
minor groove of the duplex, and this narrowing leads to
transition of DNA from B form to C-like and T-like
conformations [10].
We suggest that the observed B-C-like transition in
case of the triple complex occurs due to the joint influ-
ence of porphyrin and a metal ion on DNA in which
TOEPyP4 is an initiator of this transition. In particular,
the surface near groove and groove bindings at AT-rich
sites [13] initiate formation of a C-like conformation in
those local regions. Addition of nanomole concentrations
of Zn ions that effectively bind to these regions due to a
steric compliances of a hydrated ion with size of minor
groove of the C-like DNA causes further effective wind-
ing of C-like conformation helix that leads to narrowing
a minor groove and decrease of the distance between
flats of DNA base pairs [17]. As a result, due to the steric
conditions of TOEPyP4, it cannot stay intercalated and
moves into the solution, it does not bind to Zn ions for a
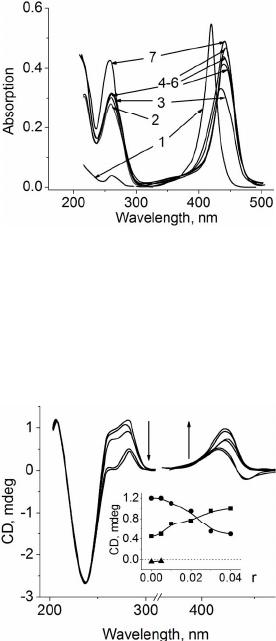
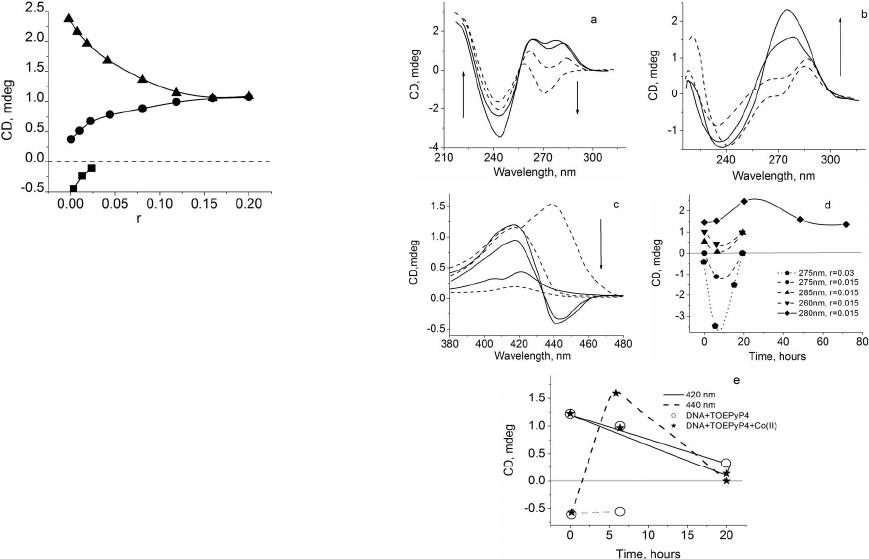
short period of time (see above and also Figure 1) and
forms a surface groove binding that causes some changes
to CD spectra, namely, disappearance of the negative
band at about 440 nm, increase of the positive band at
420 nm by 30% in Soret region, decrease of the positive
band at 285 nm, and invariability of the negative band at
245 nm. All of these demonstrate the B → C transition of
DNA (see Figure 2).
A different picture was observed when incubation of
DNA-TOEP4 complex at presence of Zn ions. In this
case, because of the fact that the binding constant of
TOEPyP4-DNA is by two orders of magnitude than the
binding constant of Me-DNA [18,19], after some minutes
at 37˚C, TOEPyP4 occupies the same sites that have
been occupied by the metal ions, and then this porphyrin
binds to those metals forming Zn-TOEPyP4 complex
form two modes of binding. These binding modes are re-
flected on CD spectra as two positive bands with maxima
at about 420 nm and 440 nm (Figure 4(c)). The intensi-
ties of these bands are significantly higher in comparison
with the some bands at 20˚C at identical TOEPyP4/DNA
ratios [12]. Consequently, the influence of Zn-TOEPyP4
complex on DNA at 37˚C is more effective than at 20˚C.
We suppose that this is the main factor that provokes
DNA transition from B to Z-like conformation, which
happens slowly—within 6 hours of incubation.
5. Summery
Analysis of the obtained CD and spectrophotometer re-
sults revealed the following: 1) combined influence of
nanomole concentrations of Zn ions and cation TOEPyP4
porphyrin initiates a DNA transition from its canonic B
form to C-like conformation; and 2) B → Z transition of
DNA in (DNA-TOEPyP4) + Zn mixture appears due to
an external binding of ZnTOEPyP4 to DNA, at 37˚C.
We think that these data will be interesting for phar-
macologists and medical specialists who use the above
mentioned porphyrins in cancer chemotherapy, because
divalent ion concentration is increased in many human
cancers compared to the norm.
6. Acknowledgements
This work is supported by the IPP/ISTC A-1951 Project.
REFERENCES
[1] C. L. Grand, H. Han, R. M. Munoz, S. Weitman, D. D.
Von Hoff, L. H. Hurley and D. J. Bearss, “The Cationic
Porphyrin TMPyP4 Down-Regulates c-MYC and Human
Telomerase Reverse Ttranscriptase Expression and Inhib-
its Tumor Growth in Vivo,” Molecular Cancer Therapeu-
tics, Vol. 1, No. 8, 2002, pp. 565-573.
[2] N. Nagesh and A. Ganesh Kumar, “Interaction of
TMPyP4, TMPyP3, and TMPyP2 with Intramolecular
G-Quadruplex Formed by Promoter Region of Bcl2 and
KRAS NHPPE,” ISRN Biophysics, Vol. 2012, 2012, Ar-
ticle ID: 786596.
[3] D. J. Cashman, R. Buscaglia, M. W Freyer, J. Dettler, L.
H. Hurley and E. A. Lewis, “Molecular Modeling and
Biophysical Analysis of the c-MYC NHE-III1 Silencer
Element,” Journal of Molecular Modeling, Vol. 14, No. 2,
2008, pp. 93-101.
http://dx.doi.org/10.1007/s00894-007-0254-z
[4] E. Izbicka, R. T. Wheelhouse, E. Raymond, K. K. Davi-
son, R. A. Lawrence, D. Sun, B. E. Windle, L. H. Hurley
and D. D. Von Hoff, “Effects of Cationic Porphyrins as
G-Quadruplex Interactive Agents in Human Tumor Cells,”
Cancer Research, Vol. 59, No. 3, 1999, pp. 639-644.
[5] Z. Zhang, J. Dai, E. Veliath, R. A. Jones and D. Yang,
“Structure of a Two-G-Tetrad Intramolecular G-Quadru-
plex Formed by a Variant Human Telomeric Sequence in
K+ Solution: Insights into the Interconversion of Human
Telomeric G-Quadruplex Structures,” Nucleic Acids Re-
search, Vol. 38, No. 3, 2008, pp. 1009-1021.
http://dx.doi.org/10.1093/nar/gkp1029
[6] J. Monaselidze, G. Majagaladze, Sh. Barbakadze, D.
Khachidze, M. Gorgoshidze, Y. Kalandadze, S. Harouti-
unian, Y. Dalyan and V. Vardanyan, “Microcalorimetric
Investigation of DNA, Poly(Da)Poly(Dt) and Poly[D(A-
C)]Poly[D(G-T)] Melting in the Presence of Water Solu-
ble (Meso Tetra (4 N Oxyethylpyridyl) Porphyrin) and Its
Zn Complex,” Journal of Biomolecular Structure and
Dynamics, Vol. 25, No. 4, 2008, pp. 419-424.
http://dx.doi.org/10.1080/07391102.2008.10507190
[7] Y. Dalyan, S. Haroutiunian, G. Ananyan, V. Vardanyan,
V. Madakyan, R. Kazaryan, L. Saakyan, L. Messory, P.
Orioli and A. Benight, “Interaction of Meso-Tetra-(4-N-
oxyethylpyridyl) Porphyrin, Its 3-N Analog and Their
Metallo-Complexes with Duplex DNA,” Journal of Bio-
molecular Structure and Dynamics, Vol. 18, No. 5, 2001,
pp. 677-687.
http://dx.doi.org/10.1080/07391102.2001.10506698
[8] M. J. Carlin, E. Mark and R. Fiel, “Intercalative and Non-
Open Access AJAC