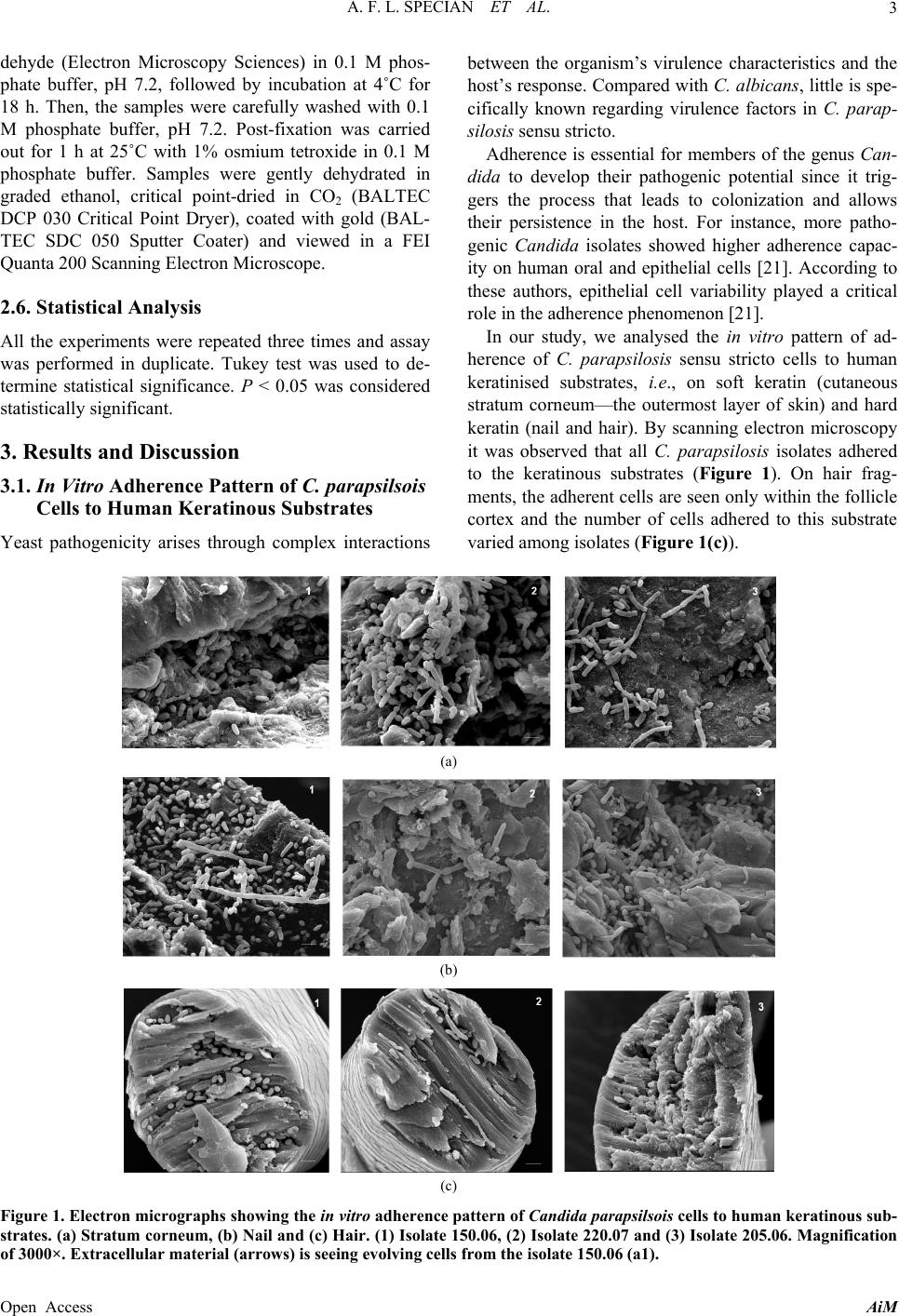
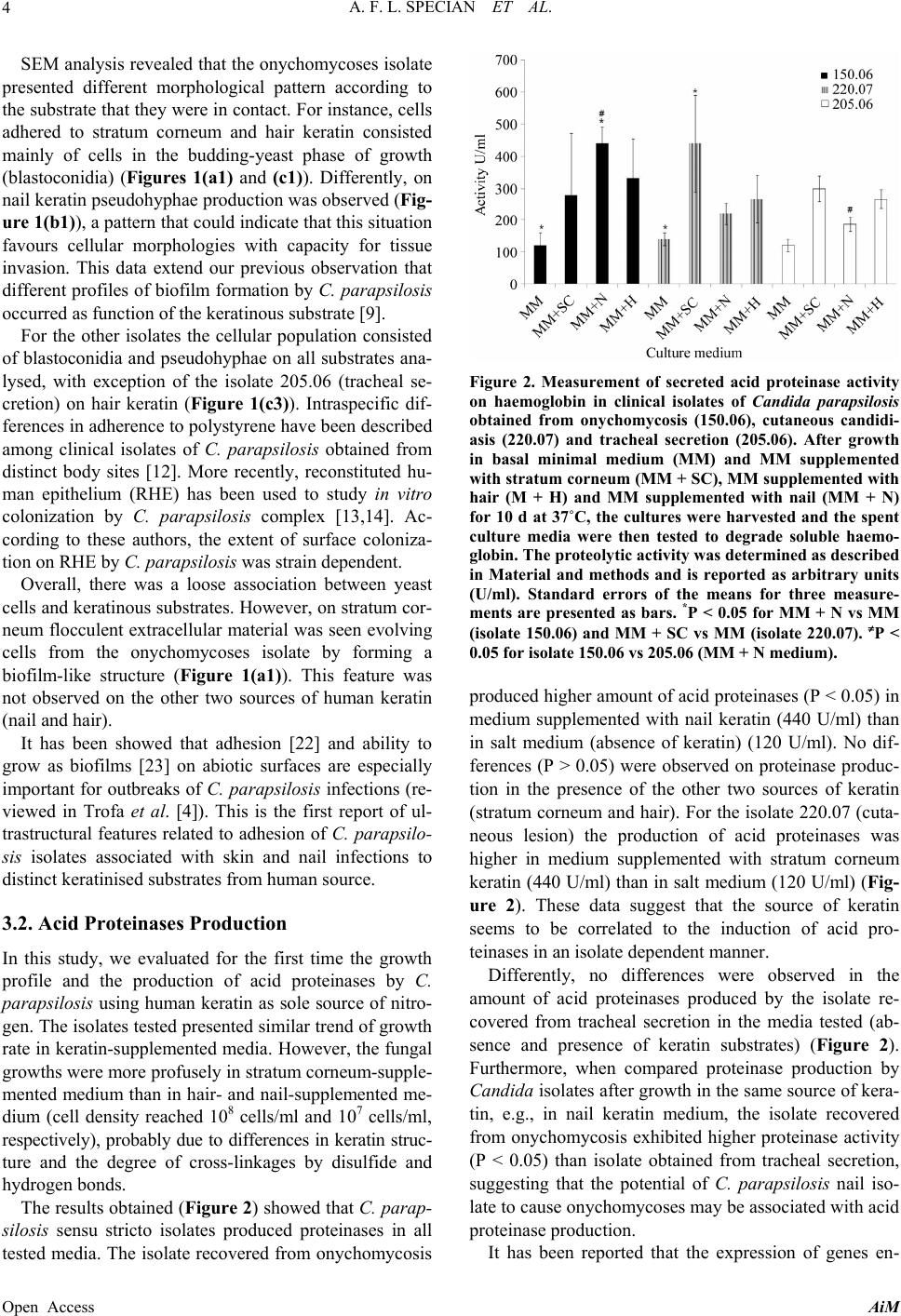
A. F. L. SPECIAN ET AL.
Open Access AiM
6
http://dx.doi.org/10.1111/j.1439-0507.2009.01825.x
[11] D. Favero, L. Furlaneto-Maia, E. J. G. França, H. P. Góes
and M. C. Furlaneto, “Hemolytic Factor Production by
Clinical Isolates of Candida Species,” Current Microbi-
ology, 2013.
http://dx.doi.org/10.1007/s00284-013-0459-6
[12] A. Sanchez-Sousa, D. Tarrago, J. Velasco, M. E. Alvarez
and F. Baquero, “Adherence to Polystyrene of Clinically
Relevant Isolates of Candida Species,” Clinical Microbi-
ology and Infection, Vol. 7, No. 7, 2001, pp. 379-382.
http://dx.doi.org/10.1046/j.1198-743x.2001.00266.x
[13] A. Gácser, W. Schafer, J. S. Nosanchuk, S. Salomon and
J. D. Nosanchuk, “Virulence of Candida parapsilosis,
Candida orthopsilosis, and Candida metapsilosis in Re-
constituted Human Tissue Models,” Fungal Genetics and
Biology, Vol. 44, No. 12, 2007, pp. 1336-1341.
http://dx.doi.org/10.1016/j.fgb.2007.02.002
[14] S. Silva, M. Henriques, R. Oliveira, J. Azeredo, S. Malic,
S. L. Hooper and D. W. Williams, “Characterization of
Candida parapsilosis Infection of an in Vitro Reconsti-
tuted Human Oral Epithelium,” European Journal of
Oral Sciences, Vol. 117, No. 6, 2009, pp. 669-675.
http://dx.doi.org/10.1111/j.1600-0722.2009.00677.x
[15] M. C. Furlaneto, J. F. Rota, R. M. B. Quesada, L.
Furlaneto-Maia, R. Rodrigues, S. Oda, M. T. Oliveira, R.
Serpa and E. J. G. França, “Species Distribution and in
Vitro Fluconazole Susceptibility of Clinical Candida Iso-
lates in a Brazilian Tertiary-Care Hospital over a 3-Year
Period,” Revista da Sociedade Brasileira de Medicina
Tropical, Vol. 44, No. 5, 2011, pp. 595-599.
http://dx.doi.org/10.1590/S0037-86822011000500013
[16] A. Tavanti, A. D. Davidson, N. A. R. Gow, M. C. J. Mai-
den and F. C. Odds, “Candida orthopsilosis and Candida
metapsilosis spp. nov. to Replace Candida parapsilosis
Groups II and III,” Journal of Clinical Microbiology, Vol.
43, No. 1, 2005, pp. 284-292.
http://dx.doi.org/10.1128/JCM.43.1.284-292.2005
[17] H. Gradisar, S. Kern and J. Friedrich, “Keratinase of Do-
ratomyces microsporus,” Applied Microbiology and Bio-
technology, Vol. 53, No. 2, 2000, pp. 196-200.
http://dx.doi.org/10.1007/s002530050008
[18] J. Friedrich, H. Gradisar, D. Mandin and J. P. Chaumont,
“Screening Fungi for Synthesis of Keratinolytic En-
zymes,” Letters in Applied Microbiology, Vol. 28, No. 2,
1999, No. 2, pp. 127-130.
[19] M. A. El-Naghy, M. S. El-Ktatny, E. M. Fadl-Allah and
W. W. Nazeer, “Degradation of Chicken Feathers by
Chrysosporium georgiae,” Mycopathologia, Vol. 143, No.
2, 1998, pp. 77-84.
http://dx.doi.org/10.1023/A:1006953910743
[20] A. L. S. Santos, I. M. Carvalho, B. A. Silva, M. B. Portela,
C. S. Alvino and R. M. A. Soares, “Secretion of Serine
Peptidases by a Clinical Strain of Candida albicans: In-
fluence of Growth Conditions and Cleavage of Human
Serum Proteins and Extracellular Matrix Components,”
FEMS Immunology and Medical Microbiology, Vol. 46,
No. 2, 2006, pp. 209-220.
http://dx.doi.org/10.1111/j.1574-695X.2005.00023.x
[21] R. D. King, J. C. Lee and A. L. Morris, “Adherence of
Candida albicans and Other Species to Mucosal Epithe-
lial Cells,” Infection and Immunity, Vol. 27, No. 2, 1980,
pp. 667-674.
[22] D. M. Kuhn, J. Chandra, P. K. Mukherjee and M. A.
Ghannoum, “Comparison of Biofilms Formed by Can-
dida albicans and Candida parapsilosis on Bioprosthetic
Surfaces,” Infection and Immunity, Vol. 70, No. 2, 2002,
pp. 878-888.
http://dx.doi.org/10.1128/IAI.70.2.878-888.2002
[23] D. M. Kuhn, P. K. Mukherjee and T. A. Clark, “Candida
parapsilosis Characterization in an Outbreak Setting,”
Emerging Infectious Diseases, Vol. 10, No. 6, 2004, pp.
1074-1081. http://dx.doi.org/10.3201/eid1006.030873
[24] M. Schaller, C. Borelli, H. C. Korting and B. Hube, “Hy-
drolytic Enzymes as Virulence Factors of Candida albi-
cans,” Mycoses, Vol. 48, No. 6, 2005, pp. 365-377.
http://dx.doi.org/10.1111/j.1439-0507.2005.01165.x
[25] M. Monod and Z. M. Borg-von, “Secreted Aspartic Pro-
teases as Virulence Factors of Candida Species,” Bio-
logical Chemistry, Vol. 383, No. 7-8, 2002, pp. 1087-
1093. http://dx.doi.org/10.1515/BC.2002.117
[26] F. De Bernardis, F. Mondello, R. San Millàn, J. Ponón
and A. Cassone, “Biotyping and Virulence Properties of
Skin Isolates of Candida parapsilosis,” Journal of Clini-
cal Microbiology, Vol. 37, No. 11, 1999, pp. 3481-3486.
[27] M. Dagdeviren, N. Cerikcioglu and M. Karavus, “Acid
Proteinase, Phospholipase and Adherence Properties of
Candida parapsilosis Strains Isolated from Clinical Spe-
cimens of Hospitalized Patients,” Mycoses, Vol. 48, No. 5,
2005, pp. 321-326.
http://dx.doi.org/10.1111/j.1439-0507.2005.01145.x
[28] A. Tavanti, L. A. M. Hensgens, S. Mogavero, L. Majoros,
S. Senesi and M. Campa, “Genotypic and Phenotypic
Properties of Candida parapsilosis Sensu Strictu Strains
Isolated from Different Geographic Regions and Body
Sites,” BMC Microbiology, Vol. 10, 2010, pp. 2-11.
http://dx.doi.org/10.1186/1471-2180-10-203