Paper Menu >>
Journal Menu >>
 Applied Mathematics, 2013, 4, 1682-1693 Published Online December 2013 (http://www.scirp.org/journal/am) http://dx.doi.org/10.4236/am.2013.412229 Open Access AM Deterministic and Stochastic Schistosomiasis Models with General Incidence Stanislas Ouaro, Ali Traoré Laboratoire d’Analyse Mathématique des Equations (LAME), Unité de Formation et de Recherche en Sciences Exactes et Appliquées, Département de Mathématiques, Ouagadougou, Burkina Faso Email: souaro@univ-ouaga.bf, ouaro@yahoo.fr, traoreali.univ@yahoo.fr Received September 11, 2013; revised October 11, 2013; accepted October 18, 2013 Copyright © 2013 Stanislas Ouaro, Ali Traoré. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT In this paper, deterministic and stochastic models for schistosomiasis involving four sub-populations are developed. Conditions are g iven under which system exhibits thresho lds behavior. The disease-free equ ilibrium is globally asymp- totically stable if and the unique endemic equilibrium is globally asymptotically stable when . The populations are computationally simulated under various conditions. Comparisons are made between the deterministic and the stochastic model. 01R01R Keywords: Computational Simulation; General Incidence; Reproduction Number; Schistosomiasis Model; Stochastic Differential Equation 1. Introduction Schistosomiasis (Bilharzia or snail fever) remains a se- rious public health problem in th e tropics and subtropics. There are about 240 million people worldwide infected and more than 700 million people live in endemic areas [1]. It probably originated around the Great Lakes of Central Africa and possibly spread to other parts of Africa, the west Indies and South America [2]. Human schistosomiasis is a parasitic disease caused by five species of the genus Schistosoma or flatworms: Schisto- soma mansoni, Schistosoma intercalatum, Schistosoma japonicum, Schistosoma mekongi and Schistosoma hae- matobium. The three most widespread are Schistosoma japonicum, Schistosoma haematobium and Schistosoma mansoni. The large number of parasite eggs retained in the host tissues rather than shed in stools can cause the inflammatory reactions. Indeed, a study conducted in Sudan indicated that schistosomiasis depleted the blood hemoglobin levels to the extent that oxygen flowed to the muscles and the brain and impaired physical activity [3]. Several mathematical models for schistosomiasis dis- ease have been done ([4-15] and the references therein). Stochastic differential equation (SDE) model is a natural generalization of ordinary differential equation (ODE) model. SDE models became increasingly more popular in mathematical biology ([4,16,17] and the references therein). In [4], a SDE model for transmission of schi- stosomiasis was analyzed. That model assumes that hu- man births and deaths are neglected. So, the compu- tational work is invo lved in a computation of B that one requires other schemes in which we solve an initial value problem. In this paper, we derive an equivalent stochastic model for schistosomiasis infection which involves four sub- populations. This model allows the recruitments and the natural deaths of human. Additionally, the incidence forms are taken as general as possible. The paper is structured as follows: In Section 2, we present the schi- stosomiasis model while Section 3 provides the basic properties of the model. Section 4 provides an analysis of the full model. We construct a stochastic differential schistosomiasis model in Section 5. In Section 6, we derive an equivalent stochastic model. We describe a numerical method to solve the equivalent stochastic mo- del in Section 7. Computational simulations are per- formed in Section 8 and finally, in the last section, we end with a conclusion. 2. Mathematical Model Here we consider a relatively isolated community where there are no immigration or emigration. Additionaly, we  S. OUARO, A. TRAORÉ 1683 suppose that all newborns are susceptible and the infec- tion does not imply death or isolation of the different hosts. Then, according to Figure 1, we obtain the following system of four differential equations d,, d d,, d d,, d d,, d shhs is i iis hii ssss is iis si HbdH fSHH t HfSH dHH t SbdSgHS t SgHSdS t (1) where s H t is the size of the susceptible human population; i H t St is the size of the infected human population; s St is the size of the susceptible snails; i b is the size of the infected snails; h is the recruitment rate of human hosts; s b is the recruitment rate of snails; h osts d is the per capita natural death rate of human h; s d is the per capita natural death rate of snails; is the treatment rate of human hosts; f is the infection function of susceptible human due to infected snails; g is the infection function of susceptible snails due to in nctions fected human. We assume that the fu f and g satisfies the fopllowing assumtions. H1 f and g are nonnegative functions on the nort. H For all , 1 C nnegative o, i S than 2 f 4 ,, sis HHS ,0 0, 0 si H fS and 0,,0 0 si gS gH. Remark 2.1 f and g are two incidence functions ich explain the contact between two species. Therefore, is no infected in the human and snail populations, then the incidence functions are equal to zero. The incidence functions are also equal to zero when there is no sus- ceptible in the human and snail populations. 3. Model Basic Properties are nnegve. Note also that when there no ati In this section, we study the basic properties of the solu- tions of system (1). Theorem 3.1 [18] Let : be a dif- ferentiable function and let L n X x be a vector field. Let a and let xa: n Gx L be such that Lx 0 for all 1: n x LaxLxa . If 0Xx Lx for all 1 x L aG then, the set is positively in- variant. Lemma 3.2 The system Equation (1) preserve the positivity of solutions. proof. We start by proving that 4 ,,,; 0 sisi i HHSS H is positively invariant. For that, let ,,, s isi x HHSS and i Lx H . Lx is differentiable and for all 0Lx 0, 1,0,0 4 :0x 1 0xLxL . The vector field on the set 4 ,,,; 0 sisi i HHSS H is , ,. hishs is sss si bfSH dH fSH Xx bdS dS (2) We obtain , is Xx LxfSH0 . Then, using Theorem 3.1, we conclude that 4 ,,,; 0 sisi i HHSS H is positively invariant. Similarly we prove that 4 ,,,; 0 sisi s HHSS H, 4 ,,,; 0 sisi s HHSS S and 4 ,,,; 0 sisi i HHSS S are positively invariant Lemma 3.3 Each nonnegative solution of model system (1) is bounded. wh f and g Figure 1. Transfer diagram. Open Access AM  S. OUARO, A. TRAORÉ 1684 Proof. Let si H tHtHt si S t. Then, adding t second two Equations and he first two equa- pectively, we get StSt tions and the of (1) res and . hh ss H bdH SbdS (3) According to [19], it follows that 0e and0e , h s dt hh dt ss ss bb St S dd (4) where 0 and hh bb HtH dd 00 si HH H 000 si SS S. , Thus lim t and lim hs t hs bb Ht St dd (5) Corollary 3.4 The set 4 ,, sis KHtHtS , ; 0,0 i hs si si hs tSt bb Ht HtStSt dd is invariant and attracting for system (1). Theorem 3.5 For every nonzero, nonnegative initial ist for all time dard uments since the right-hand side of system (1) is lo nd analyze the eventual equilibria. For that, let us denote value, solutions of model system (1) ex 0. Proof. Local existence of solutions follows from stan- arg t cally Lipschitz. Global existence follows from the a priori bounds. 4. Reproduction Number and Equilibria Analysis In this section, we define the reproduction number a 12 ,, ,. is is is gg 12 ,, ,, is is is ff fSHf SH SH g HS gHS HS System (1) has a disease-free equilibrium (DFE) given by 00 ,0,,0,0,,0 . hs bb EHS DFE s shs dd Proposition 4.1 The reproduction number fr system (1) is o 00 0, 0, 11 0. ss sh f HgS dd R Proof. We use the method in [20] to compute the reproduction number Let 0 R. 12 , X XX, where 1, s s X HS =, ii is the healthand y population 2 X HS , the infected population. 1 ,hi i is idH H fSH H XS 12 , . si is idS gHS X 12 ,,XX X Then, 1 11 2 1 2 00, ,0 0, 0 0 and ,0. 0 s s h s fH FX gS X d VX d X Hence, 1 11 1 0, 100 and . 10, 00 s hs s sh fH dd VFV gS dd The basic reproduction number is defined as the dominant eigenvalue of matrix 1 F V [20]. Therefore, 00 11 0 0, 0, ss sh dd The basic reproduction numvaluate fHgS R ber ethe average nunfections generated by a single infected individual in a completely susceptible population. Using Theorem 2 in [20], the following result is estab- lished. mber of new i Lemma 4.2 If 01 R, then D FE E is locally asymptotically stable. Proof. Using the assumption H2, it follows that 0, 0fH 2s and 0, 0gS for all 2s s H and s S. Then, the linearization of system (1) at D FE E gives the following equation 11 0 sh dd f00 ,0, 0. ss H gS hs dd (6) We can see that all solution of Equation (6) have negative real part. Indeed, the Equation (6) has negative roots h d , s d and oots are given by ther ro 0 11 0, 0, shs s dd fHgS 0 . (7) Now, we suppose that there exts at least one root 1 is of the Equation (7) which have positive real part. According to Proposition 4.1, it follows that Open Access AM  S. OUARO, A. TRAORÉ 1685 0 11 0 0 s fHg 02 0 , ,. ssh S dd R Since , we obtain 0<1R 00 11 11 0, 0, sssh sh HgS dddd.f This show that the Equation (7) cannot have roots with positive real part. Hence, D FE Eing to T is locally asymptotically stable accordheorem 2 in [20] et us abal behavior of the DFE. The global stability of the DFE will be studied using the basic reproduction number . We first make the fo Now, lnalyze the glo 0 R llowing additional assumption. H3 For all 4 ,,, sis HHSS , 0 1 ,0, is si i f SHfH S and 0 1 ,0, iss i g HSgS H. Theorem 4.3 The disease-free equilibrium is globally asymptotically stable in K whenever 0<1R. Proof. The proof is basedon using a comparison theorem [21]. Note that the equations of the infected co system (1) can be essed as mponents inxpre , d i i i FV S S (8) d di HH t dt where F and V, are defined in the proof of Pro- position 1. Using the fact that all the eigenvalues of the matrix F V whenev eigenvalues have negative real parts, it follows that the linearized differential inequality syste er . Indeed, we can see that all m (8) is stable 0<1R of the matrix F V satisfy Equation th(7). Bye sameasoning as in the proof of Lemma 4.2 we deduce that, the eigenvalues of the matrix re F V have negative real parts since 0<1R. T, hus ,0,0 ii HtSt as t for the system (8). Consequently, by a standard comparison theorem [21 ,0,0 ii tSt as t and substituting 0 ii HS into system (1) gives 0 ], H s s H H and 0 s s SS as t. Thus, 00 ,,, ,0,, sisi s HtHtStStH as t for 0<1R. The0 s S refore, D FE E is globally lly stable if 0<1R. as icymptot Lemma 4.4 If 01R then, a D FE E is unstable and there exists a unim qendemic equilibriue u ,,, s i EHHS Proof Let si S . . ,,, s is ES . It follows that i SHHE is a positive equilibrium if and only if ,0, hhs is bfSH H H ,0 , 0, ihi s fSH H (9) , ,0 . i s i ss is is si dH d bdSgHS HS dS wo equations of sys- tem (9) respectively, we get g Adding the first two and the last t 0 and0. hhshissssi bdHdHb dSdS Then, and . hhi ssi ss hs bdH bdS HS dd Let ,, ,, . hhi iii h ssi hiisi s bdH hS HfSd bdS dHgH dS d on is continuous and so sy (9) hold whenever We can see that the functi stem h ,0hS . Therefore any zero of h in the set ii H 0, 0, sh sh bb dd where i H and i S are positives corresponds to an endemic e. According to H2, quilibrium 0,00h and ,0 sh bb h sh dd olution of equ Then, in order to have a sation 0h . in 0, 0, sh sh dd , h 0. This lea to bb must increase at ds 2 d0,0 0, h (10) dX where 2 X is defined in the proof of Proposition 4.1. We can see that 00 0, , 0,. sh ss 11 2 d d h f Hd gSd X It follows from inequality (10) that 0 1 1 0 1 0 1 0, 0 or 0,0 and 0,0. sh ss sh ss fHd gSd fHd gSd 0 0, 0and 0 0,fHg Which implies that 0 11 0, 1 ss sh S dd , that is To study the global behaviour of this endemic equilibrium, we second make this additional assumption. H4 For all 01R. 4 ,,, sisi HHSS , Open Access AM  S. OUARO, A. TRAORÉ Open Access AM 1686 , 1, , and 1. , is si i is s is s i i is s fSH HS S fSH H gHS SH H gHS S and ,, ,, ,, ,. s hsi sss i hii sii s ssis ss i siis ii H t VtgHSHh H Ht VtgHSHhH St VtfSHSh S St Vtf S HShS (11) * (14) Theorem 4.5 When 0 and 01R, the endemic equilibrium E is globally asymptotically stable in K. when 1R, We can see that * :h has the strict global minimun 10h . Thus, hs with equality if and only if 0,VV0, hi ss V0, 0 si V ,, s siiss H HH HSS and ii . We will study the behaviour of the Lyapunov function Proof. If wem ere exists a e consider the syst (1) thunique endemic equilibrium 0 E. We now ic establish the global asymptotic stability of this endem equilibrium. Evaluating both sides of (1) at E with 0 SS gives . hs hi ss si Vt VVVV (15) ,, ,, ,, ,. hhs is hiis sss is sii s bdHfSH dHfS H bdSgHS dSg HS We can see that 0Vt with equality if and only if 1,1,1 and1. sisi sisi Ht Ht StSt HHS S (12) (1 The derivatives of and ,, hs hi ss VVV s i will be calculated separately and then combined to get the V Let 1lnhx xx 3) desired quantity d d V t. ,1,. s ishhsi s s H Sb dHfSH tH UsiEquation of (12) to replace gives dd ,1 dd hsss is s VHH gHS gH tH ng the first h b 2 2 d,1, , d , ,,,11 , ,, ,,,1 . ,, hs s ishssi sis s ss is s hisis is s s is ss is is ss hisisss is is VH gHSd HHfSHfSH tH HH fSH H dgHSgH Sf S H HH fSH HH fSH fSH HH dgHSgH Sf S H HHH fSH fSH By adding and substracting the quantity is s , 1ln , is s s is fSH H H fSH , we obtain 2 2 d,1ln ,,, , 1ln 1ln ,,, , , ,,, ss ss is ss isis isis ss ss isis isis ss i s hisis is ss HH VHH gS HHH fSHfSH fSHfSH HH HH fSHfSH fSHfSH HH fSH H dgHSgH Sf S Hhh HH ,, dhs hi s is s dgH SH Sf tH ,. ,, sis s s is is fSH H hH fSH fSH (16)  S. OUARO, A. TRAORÉ 1687 d dhi V t. Next, we calculate dd ,1(,)1, d , ,1 ,. , hiiii isisishi ii is ii isi shi ii is VHHH g HSgHSfSHdH tHdtH fSH HH gHS fSHdH HH fSH hi dH Using the second Equation of (12) to replace gives ,, d,,1,,1 d,, isis is hiiii i is isis is ii ii isis is fSHfSH fSH VHH HH fSHgHSfSHgHS tHHHH fSHfSH fSH , . , By adding and substracting the quantity , 1ln , is i i is fSH H H fSH , we obtain ,, d,, 1ln1ln d,, , , n, ,, , is is hii iii is isii ii is is isis is ii is isii is isisis fSHfSH VHHHH fSHgHS tHHHH fSHfSH SHfSHfSH HH fSHgHShhh HH SHfSHfSH . ,, 1l ,, is fSH f fSH f (17) Now, we calculate d d s s V t. dd ,1 ,1, dd sss ss isissssis ss VSSS fSHfSHb dSgHS tStS . Using the third Equation of (12) to replace s bes giv 2 2 d,1, , d , ,,,11 ,, ,,,1 . ,, ss s isss sisis s ss is s is is ss is is ss sisis is sss is is VS fSHdSSgHSgHS tS SS gHS S df SHf SHgH SS S SS gHSgHS SS df SHf SHgH S SSS gHSgHS By adding and substracting the quantity , si s s sis S gH , 1ln , is s s is gHS S S gHS , we obtain 2 d,1ln ,,,, 1ln 1ln ,,,, , ,,, ss ss sisis is sss isis isis ss ss isis isis i s sisisis ss SS VSS Hf S HgH S S SS gHSgHSgHSgHS SS SS gHSgHSgHSgHS gHS S df SHfSHgH Shh SS ,, dss df S t 2 ss SS ,. ,, sis s s is is gHS S hS gHS gHS (18) Open Access AM  S. OUARO, A. TRAORÉ Open Access AM 1688 After that, we evaluate d d s i V t. dd ,1 dd ,1 , , ,1 ,. sii i is i i isis si i is ii isis si , i VSS fSH tSt S fSHgHS dS S gHS SS fSH gHSdS i S is S gHS Using the last Equation of (12) to replace , d,,1 d, ,, ,, 1. ,, is s ii isi sii is is is is is ii ii is is gHS VS gHS fSH tS gHS fSHgHS gHSgHS SS SS gHSgHS i S S By adding and substracting the quantity , 1ln , i gH gH s i dS gives si i is SS S S, we obtain d,, 1ln d ,,, 1ln 1ln ,,, ,, ,, ,, sii i is isii isis isis ii ii isis isis is is ii is isii is is VSS fSHgHS tSS gHSgHSgHSgHS SS SS gHSgHSgHSgHS gHSgHS SS fSHgHShhh SS gHSgHS . , , 6)-(1, we obtain (19) Combining equations (19) 2 2, d,,,, d, ,, , ,, , ss ss is is hiss isisis s si is isi sis is issi is issi isi sis SS HH fSH SH VdgHSdfSHgHS fSHhh tH SS fSH gHSfSHgHS HSHSHS hh hhhh HSHSHS gHSfSHgHS s H . (20) Since the function is monotone on each side of 1 and is minimized aes t 1, H4 implih ,, and . ,, is is si s si s is is fSH gHS HS SH hhh HS SH fSH gHS i i h Since , then 2 t . 0h d0, d V t (21) for all ,,, sisi H HSS K with equality only for ,, s siis s H HH HS S and ii SS. rium Hence, the endemic equilibE is the only pontained in sitively invariant set of the system (0.1) co 4 ,,,;,, , s isiss iissii H HSSHHHHS SS S . Then, it follows that E is globally asymptotically stable on [22]. 5. Stochastic Differential Equation Model To derive a stochastic model, we apply a similar procedure to that described in Allen [23]. Here, we neglect the po ssibility of multiple even ts of order K The possible changes in the popu lations over a sho rt time t , concern individual births, deaths and transform se changes are produced in Table 1, togeth corresponding probability. Let’s denote th ation. er with is change The their by T ,,, sisi H HS S . Neglecting terms of the order , the m system (0.1) is given by t 2 tean of 10 1 , ,. , , hhs is i is hii ii isss is is si bdH fSHH fSH dHH EP t bdSgHS gHSdS (22)  S. OUARO, A. TRAORÉ 1689 Table 1. Possible changes in the population. Change Probability T 11,0,0,0 1h Pbt T 21,0,0,0 2hs PdHt T 31,1,0,0 3, is PfSHt T 0, 1,0,0 44hi T 1, 1,0,0 PHt PdHt 55i T 60,0,1,0 6s Pbt T 70,0,1,0 7ss PdSt T 80,0, 1,1 8, is PgHS t 0,0, T T 90, 1 9si PdSt 10 0,0,0,0 9 10 1 1i i PP , the covariance ma by 23) Furthertrix of system (1) is given 11 12 133 34 00 0 iii i BB Bt BB 10 12 22 TT 34 44 00 , 0 00 BB EP t BB ( where 11 22 12 33 ,, ,, ,, ,, , hhs isi is hii is i sss is BbdHfSHH BfSHdHH BfSHH BbdSgSH BgSH 44 and , is s Bg SHdS Note that it has been 34 . is i proved in [23] that the changes are normally distributed. Then, ,tt ttt ttBt YY YY where for Furt converges strongly to the solution of thm 0,1 iN hermore, as 1, 2,,4i. 0, tYt e stochastic syste ddtt YW , dd tB t YY (24) where and isthe four- dimens i3]. The computationalwork of Equation (24) involves the calculation of the qutity tt T ,,, sisi tHHSSY onal Wiener process [2 tW an BtY ften cumbat each time step [24,25]. This procedure is oersome. In the next section, we derive an equivalent ochastic model which seem toe easier to implement. 6. Equivalent Stochastic Differential Equation Model In this section, we develop a stochastic model to e the changes occured on each vector individually. Unl we say otherwise, we adopt the vectors defined in the previous section but here the Poisson p rocesses st b xamine ess P ean ti are used to establish the different probabilities as mmes before occurence. Then, we have 1234 345 678 89 , , , , s i s i H uuuu Huuu Suuu Suu (25) where 12 34 567 89 ,, ,,, , ,, . hhs is i hi sss is si uPbtuPdHt uPfSHtu PHt uPdHtuPbtuPdSt uPgHStu PdSt ,, We now normalize the Poisson process to get 12 34 3 54 67 8 8 ,, , ,, , ,,, ,, sh hhshs isisii iisis hi hiii ss sssss is is iisis si tbtdHtdHtHb f SH tfSHtHtHt H fSH tfSH t dHtdHtHtHt Sbt btdStdSt StgHSt SgHS tgHS t dS g H 9, si tdSt (26) where 0,1 iN for 1, 2,,9i . Then, as 0t , chastic the systemergellowing Itô sto differential equation [24] (26) convs to the fo 1 2 d,dd ,d shhs is ih hsi s HbdHfSH HtbW dHdWfS H 3 4 3 6 78 89 d, d, d,d , d,d, d,d,dd. i iishiiis h ssi s iissiis si WH W HfSHdHHtfSHW HW dSWg HSW SgHSdStgHSWdSW llows. 45 dd d, dd ii ssss iss HW Sb dSgHStbW d (27) System (27) can be rewritten as fo Open Access AM  S. OUARO, A. TRAORÉ Open Access AM 1690 dd , dd tt t tt YW Y (28) w G here tY and are the same as in system (24), W is the nine-dimensional Wiener process and G is defined by 1234 3 00 00 00 GGGG GG 4 5 0 00 0 0 , G G 34 56 78 9 ,, ,, ,, and . is i hi s , s si si GfSHG H GdHGb GdSGgHS GdS 678 89 00 0000 00 00000 GGG GG (29) where 12 , h GbG, hs dH s 7. Computational Results In this section, computational results are given for the stochastic system (28). We use the Euler-Maruy method with 1000 sample to solve the SDE model Le yama numerical method for system (28) is given by: ama (28). t h be a specified time step. The Euler-Maru 1 1 ,, , kk sshhs h kk iiishiii HHhbdHf S HHhdH HHhf SHdHHhf S 1,2,3,4, 3, 4,5, , , , hkskisk ik ski khi k bfS HH HHdH isi 16, 7,8, 18, 9, ,,, ,,, kk ss sssissksskisk kk iiissiisksi k SShbdSgHS hbdSgHS SShgHSdSh gHSdS for until the maximum time is reached. Here 0, 1,2,k , ,ik for rmally distribute1,2,,9i d numbers and are no with zeunit varia 8. Computational Simulations In this section, computational simulations are given for the models described in the previous sections. Here, two different cases of computational simulations were studied: in the first case while in the second case . In the co, the functions 0, 1,2,k ro mean and nce. 01R mputations 01R f and g are chosen as follows , is si f SH SH and is i ,s g HS HS (mass action), where is the rate rcariainfection of hosts by ce released by an infected snail and is the infection rate of snails by imracidia of the parasite eggs of an infected human. The parameters values used are given as: 0.014 h d (human beings average life span years 701h d ), n is 14b h 1000), sails apan is 2 - 8, 400 s b (the carrying capacities of snails population is 2000), 0.0004 (the carrying capacities of humn pop (sn veragea life sulatio years) 0.2d [26], 0.000027 [26e ]. Th treatment rate to in the whic second case thee have first treatm 01.29 case is 0.1 h give nt rate is 00.97R chosen as . In the 0.05 R are 30 . The initial values , was taken as 300 years. These figures are produced by Matlab. Figure 2 illustrates the deterministic model (1) and t equivalent stochastic model (28) when We t, in the Figure 2 the trajectory of inistic and stochastic graphs are approximy the same behaviour. Indeed, the parasite extinction i effective if of the i H step h popula 0400, S was c tion sizes s hosen as take H 0, 0 i S 0.3 n as 07 year and the fi 060 . The time nal time 0 s 001 h he can 01R. de- term atels see tha 01 R. Figure 3 depict the deterministic model (1) and the equivalent stochastic model (28) when. In Fig- ure 3 we can observe that, the features ministic graph are similar to those of the stochastic namely the parasite extinction is unlikely for thiodels when results we made about the 01R of eter graph, s two m contains the d 01R. The Table 2 and Table 3 below that supports the comparison Table 2. Mean and standard deviation for the ODE epi- demic model (1) and the SDE epidemic model (28) at t=300 years where 0 R= 0.97. Models Variables i X i XE i Xσ s H 992.25 0 i H 7.74 0 ODE (1) s S 1968.63 0 31.36 0 i S s H 993.21 3 2.49 i H 6.44 9.44 SDE (28) s S 1985.71 719.28 26.12 38.78 i S 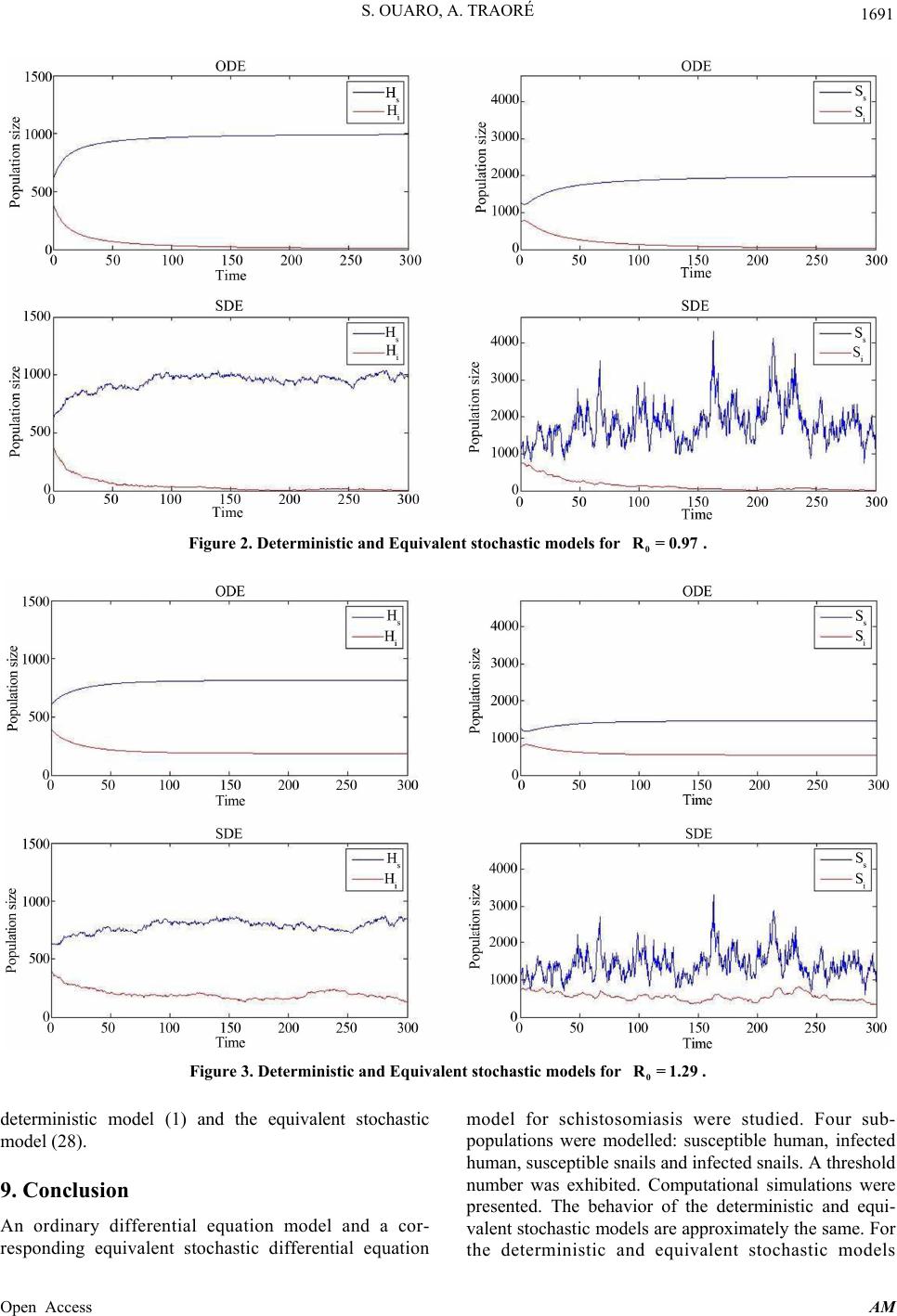 S. OUARO, A. TRAORÉ 1691 stic models for . Figure 2. Deterministic an 0 R= 0.97 d Equivalent stocha Figure 3. Deterministic and Equivalent stochastic models for deterministic model (1) and the equivalent stochastic model (28). 9. Conclusion An ordinary differential equation model and a cor- responding equivalent stochastic differential equation model for schistosomiasis were studied. Four sub- populations were modelled: susceptible human, infected human, susceptible snails and infected snails. A threshold number was exhibited. Computational simulations were presented. The behavior of the deterministic and equi- valent stochastic models are approximately the same. For the deterministic and equivalent stochastic models 0 R= 1.29 . Open Access AM  S. OUARO, A. TRAORÉ 1692 Table 3. Mean and standard deviation for the ODE epidemic model (1) and the SDE epidemic model (28) at years where Models Variables t=300 0 R= 1.29 . i X i XE i Xσ s H 813.52 0 i H 186.47 0 ODE (0.1) s S 1456.71 0 543.28 0 i S s H 818.21 42.05 i H 181.56 36.85 SDE (0.28) s S 1474.13 451.02 532.71 148.20 i S the disease dies out for and the disease limits to an endemic equilibrium REFERENCES [1] World Health Organization, 2012. http://www.who.int/schistosomiasis/en/ P. Jordan and G. Webbe, “Hum les Thomas, Springfield, 1969. [3] P. L. Rosenfield, “The Management of Schistosomiasis,” Resources for the Future, Washington DC, 1979. [4] E. J. Allen and H. D. Victory, “Modelling and Simulation of a Schistosomiasis Infection with Biological Control,” Acta Tropica, Vol. 87, No. 2, 2003, pp. 251-267. http://dx.doi.org/10.1016/S0001-706X(03)00065-2 01R when R01. [2] an Schistosomiasis,” Cha- [5] R. M. Anderson and R. M. May, “Prevalence of Schisto- some Infections within Molluscan Populations: Observed Patterns and Theoretical Predictions,” Parasitology, Vol. 79, No. 1, 1979, pp. 63-94. http://dx.doi.org/10.1017/S0031182000051982 [6] R. M. Anderson and R. M. May, “Helminth Infections of Humans: Mathematical Models, Population Dynamics, and Control,” Advances in Parasitology, Vol. 24, 1985, pp. 1-101. http://dx.doi.org/10.1016/S0065-308X(08)60561-8 [7] J. E. Cohen, “Mathematical Models of Schistosomiasis,” Annual Review of Ecology and Systematics, Vol. 8, 1977, pp. 209-233. http://dx.doi.org/10.1146/annurev.es.08.110177.001233 [8] Z. Feng, C. Li and F. A. Milner, “Schistosomiasis Models with Density Dependence and Age of Infection in Snail Dynamics,” Mathemati 2002, pp. 271-286. ] Z. Feng, C. Li and F. A. Milner, “Schistosomiasis Models r Modelling, Vol. 41, 2005, pp. 1213-1230. [10] Z. Feng and F. A. Milner, “A New Mathematical Model asis,” In: Innovation Applied Mathematics, iversity Press, Nashville, 1998, pp. 117- 128. [11] G. Macdonald, “The Dynamics of Helminth Infections, with Special Reference to Schistosomiasis,” Transactions of the Royal Society of Tropical Medicine and Hygiene, Vol. 59, No. 5, 1965, pp. 489-506. http://dx.doi.org/10.1016/0035-9203(65)90152-5 [12] I. Nasell, “A Hybrid Model of Schistosomiasis with Snail Latency,” Theoretical Population Biology, Vol. 10, No. 1, 1976, pp. 47-69. http://dx.doi.org/10.1016/0040-5809(76)90005-8 [13] M. E. J. Woolhouse, “On the Application of Mathemati- cal Models of Schistosome Transmission Dynamics. I. Natural Tra nsmission,” Acta Trop. 49, 1241-1270, (1991). http://dx.doi.org/10.1016/0001-706X(91)90077-W [14] Woolhouse, M.E.J., On the application of mathematical models of schistosome transmission dynamics. II. Control. Acta Tropica, Vol. 50, 1992, pp. 189-204. http://dx.doi.org/10.1016/0001-706X(92)90076-A [15] Y. Yang and D. Xiao, “A Mathematical Model with De- lays for Schistosomiasis,” Chinese Annals of Mathematics, Vol. 31B, No. 4, 2010, pp. 433-446. http://dx.doi.org/10.1007/s11401-010-0596-1 [16] L. J. S. Allen, “An Introduction to Stochastic Processes with Applications to Biology,” Prentice-Hall, Englewood Cliffs, 2003. duction to Stochastic Differential Equa- tions,” Marcel Dekker, New York, 1987. [18] J. M. Bony, “Principe du Maximum, Inégalité de Harnack et Unicité du Problème de Cauchy Pour les Opérateurs Elliptiques Dégénérés,” Annales de L’institut Fourier (Grenoble), Vol. 19, 1969, pp. 277-304. [19] G. Birkhoff and G. C. Rota, “Ordinary Differential Equa- tions,” Ginn, Boston, 1982. [20] P. Van den Driesche and J. Watmough, “Reproduction Numbers and Subthreshold Endemic Equilibria for the Compartmental Models of Disease Transmission,” Ma- thematical Biosciences, Vol. 180, No. 1-2, 2002, pp. 29- 48. http://dx.doi.org/10.1016/S0025-5564(02)00108-6 cal Biosciences, Vol. 177-178, [9 with Two Migrating Human Groups,” Mathematical and Compute of Schistosomi Vanderbilt Un [17] T. C. Gard, “Intro [21] V. Lakshmikantham, S. Leela, A. A. Martynyuk, “Stabil- ity Analysis of Nonlinear Systems,” Marcel Dekker, New York, 1989. [22] J. P. LaSalle, “The Stability of Dynamical Systems,” SIAM, Philadelphia, 1976. http://dx.doi.org/10.1137/1.9781611970432 [23] E. J. Allen, “Stochastic Differential Equations and Persis- tence Time of Two Interacting Populations,” Dynamics of Continuous, Discrete and Impulsive Systems, Vol. 5, 1999, pp. 271-281. [24] E. J. Allen, L. J. S. Allen, A. Arciniega and P. E. Green wood, “Construction of an Equivalent Stochastic Diffe odels,” Ststic Analysis and Appli- , 2008. 274-297. - r- ential Equation Mocha cations, Vol. 26, No. 2, pp http://dx.doi.org/10.1080/07362990701857129 [25] E. J. Allen, J. Baglama and S. K. Boyd, “Numerical Ap- proximation of the Product of the Square Root of a Ma- trix with a Vector,” Linear Algebra and Its Applications, Vol. 310, No. 1-3, 2000, pp. 167-181. http://dx.doi.org/10.1016/S0024-3795(00)00068-9 Open Access AM  S. OUARO, A. TRAORÉ Open Access AM 1693 [26] Z. Feng, A. Eppert, F. A. Milner and D. J. Minchella, “Estimation of Paramaters Governing the Transmission Dynamics of Schistosomes,” Applied Mathematics Let- ters, Vol. 17, No. 10, 2004, pp. 1105-1112. http://dx.doi.org/10.1016/j.aml.2004.02.002 |