Agricultural Sciences
Vol.06 No.11(2015), Article ID:61350,9 pages
10.4236/as.2015.611127
Can Improve Iron Biofortification Antioxidant Response, Yield and Nutritional Quality in Green Bean?
Juan P. Sida-Arreola1, Esteban Sánchez1*, Graciela D. Ávila-Quezada2, Paul B. Zamudio-Flores1, Carlos H. Acosta-Muñiz1
1Centro de Investigación en Alimentación y Desarrollo, Chihuahua, México
2Facultad de Zootecnia y Ecología, Universidad Autónoma de Chihuahua, Chihuahua, México

Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).


Received 22 October 2015; accepted 20 November 2015; published 23 November 2015

ABSTRACT
The aim of this study was to evaluate the effect of iron biofortification on antioxidant response, yield and nutritional qualityof green bean (Phaseolus vulgaris L.) under greenhouse conditions. Fe was applied using two forms (FeSO4 and Fe-EDDHA) at four doses of application (0, 25, 50 and 100 µm) added under a hydroponic system, and were tested over a period of 40 days. The Fe content was assessed in seeds, as well as the activity of antioxidant enzymes, production of H2O2, yield and nutritional quality. The results being obtained indicated that the accumulation of Fe in bean seeds enhanced with the application of Fe-EDDHA at the dose of 25 µm. This demonstrated that low Fe application dose was enough to increase Fe levels in seeds of common bean. In addition, Fe- EDDHA application form at 50 µmol was the best treatment to improve crop yield. Respect to antioxidant system, chelated form of Fe (Fe-EDDHA) was more effective in the activation of antioxidant enzymes (CAT, SOD and GSH-PX), and a lower content of H2O2 in green bean seeds. Finally, to raise the Fe concentration in bean under biofortification program was a promising strategy in cropping systems in order to increase the ingestion of iron and antioxidant capacity in the general population and provided the benefits that this element offered in human health.
Keywords:
Phaseolus vulgaris L., Antioxidant Activity, Biofortification, Iron, Nutritional Quality

1. Introduction
Beans (Phaseolus vulgaris L.) are grown and consumed in nearly all the world. In many developing countries, 20% of the available proteins are provided by beans and represent also integral part of dietary protein for 50% of the world’s population [1] . This crop is cultivated mainly in the American Continent and East Africa. Edible crops, such as beans, require certain concentrations of macro and micronutrients for optimal development and yield [2] . Biofortifications is a prominent strategy that can increase levels of micronutrients in crops. Agronomic and genetic biofortification represent complementary agricultural approaches. It is considered that application of Fe fertilizers is a short-term solution and complement to plant breeding [3] .
Today, there is a convincing evidence that especially foliar application of micronutrients fertilizers (such as selenium and zinc) are effective in improving their concentration in grain of Phaseolus vulgaris L. cultivars [4] - [6] . Respect to Fe biofortificacion, it appears that improving nitrogen nutritional status of plants promotes accumulation of Fe in grain. Therefore, the plant nitrogen status deserves special attention in biofortification of food crops with Fe. Studies under both field and greenhouse conditions have demonstrated that increasing N application (soil or foliar spray of urea) significantly improves shoot and grains Fe concentrations in gramineous species [3] [4] [7] . However, a role of Fe fertilizers and their application methods in improving Fe concentration in legumes is rarely studied.
Fe is an essential micronutrient [8] and cofactor of many antioxidant enzymes, and it can act as pro-oxidant [9] . It has been shown that many enzymes require Fe in order to function properly. In particular, Fe is present in the active site of enzymes on antioxidant pathway involved in the scavenging of reactive oxygen species (ROS) [10] . Production and activity of ROS have been reported frequently in several plants under different types of stresses [11] . Failure to quench ROS or subsequent propagation of chain reaction may cause oxidative stress and lead to injury. The generation of ROS such as superoxide anion radical (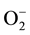 ), singlet oxygen (O2), hydrogen peroxide (H2O2) and hydroxide radical (OH) can damage many cellular components, including protein, membrane lipids and nucleic acids.
), singlet oxygen (O2), hydrogen peroxide (H2O2) and hydroxide radical (OH) can damage many cellular components, including protein, membrane lipids and nucleic acids.
To face with increased levels of H2O2, plants have evolved different enzymatic and nonenzymatic mechanisms [11] . These include free radical scavengers, such as superoxide dismutase (SOD), catalase (CAT), and peroxidases (GPX), and the enzymes involved in the ascorbate-glutathione cycle [12] . The enzymatic processes basically involve dismutation of O2 by SOD that generates another partially reduced oxygen species, like H2O2 [11] . H2O2 is generated in cells by the direct transfer of two electrons to the superoxide anion (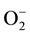 ). This is mediated by enzymes such as glycolate and glucose oxidases, or by the dismutation of O2, which frequently occurs in chloroplasts and peroxisomes under conditions of high light stress. In the dark, or in non-photosynthetic tissues, H2O2 is produced mainly by leakage of the electron transport chain in mitochondria or microsomes [13] and during fatty acid oxidation [14] .
). This is mediated by enzymes such as glycolate and glucose oxidases, or by the dismutation of O2, which frequently occurs in chloroplasts and peroxisomes under conditions of high light stress. In the dark, or in non-photosynthetic tissues, H2O2 is produced mainly by leakage of the electron transport chain in mitochondria or microsomes [13] and during fatty acid oxidation [14] .
Many researches have identified several beneficial effects of biofortification with micronutrients, which include antioxidant properties that can stimulate plant growth and protect plants against different types of abiotic stress [5] [6] [15] . It demonstrates that biofortification with micronutrients (such as selenium, zinc and iron) induces higher increases in enzymatic activities that detoxify H2O2, as well as an increase in the levels of antioxidant compounds (ascorbate, glutathione, etc). Thus, this technology than promoting the application of micronutrients at low rates can be used to control the induction in bean plants of the antioxidant system, thereby improving crop yield, stress resistance and accumulation of antioxidant compounds in bean seeds [6] [15] .
The aim of the present study was to analyze the effect of Fe biofortification, with different application rates and forms of Fe, on enzymatic activities of SOD, CAT and GSH-PX as bioindicators of the antioxidant system efficiency and improve fruit yield of bean plants (Phaseolus vulgaris L.).
2. Materials and Methods
2.1. Crop Handling
Seeds of green bean (Phaseolus vulgaris L., cv. Strike) were germinated and grown in a substrate mix (peat moss, vermiculite and perlite at a ratio 3:1:1) at an experimental greenhouse located in Delicias, Chihuahua, Mexico. Green beans seeds cv. Strike was acquired from the company Agrow Mexicoin Mexico City. This variety of green beans is characterized as a kind of short cycle (60 days), erect bearing, consumed both green pods and dry grains, thrives in warm and cold climates, it is grown in not very saline soils is suitable to grow in greenhouses. The temperature inside of the greenhouse was controlled at 25˚C ± 4˚C, relative humidity of 60% - 80%, and a photoperiod of 16/8 h (light/darkness). The plants were grown in individual pots (25 cm upper diameter, 17 cm lower diameter, 25 cm in height) of 8 L volume, filled with substrate mix. Throughout the growing cycle the bean plants received a growth nutrient solution composed of 6 mm NH4NO3, 1.6 mm K2HPO4, 2.4 mm K2SO4, 4.0 mm CaCl2∙2H2O, 1.4 mm MgSO4, 5 µM Fe-EDDHA, 2 µM MnSO4∙H2O, 1.0 µM ZnSO4∙7H2O, 0.25 µM CuSO4∙5H2O, 0.3 µM (NH4) Mo7O24∙4H2O and 0.5 µM H3BO3 (pH 5.5 - pH 6.0). For Fe biofortification, plants were subjected to different Fe treatments described in next experimental design section. These Fe treatments were applied in combination with the nutrient solution for 40 days (beginning 20 days after of seed germination). The application of the different treatments of Fe was through a hydroponic system for 40 days.
2.2. Experimental Designs and Treatments
In this study was used a completely randomized experimental design, with different forms of Fe at different concentrations. This were four treatments of Fe chelate, at 4 doses and 4 replicates per treatment: Fe-EDDHA (Fe-EDDHA) and FeSO4 (FeSO4) at doses of 0, 25, 50 and 100 μmol, respectively. Doses and application forms Fe were selected based on the study of Hermosillo-Cereceresetal [6] .
2.3. Plant Sampling
Plant materials were sampled at 60 days after germination, when the plants had a phenological phase of complete development and fruit maturity. The different organs of each bean plant were separated (leaf, petiole, stem, root, pod, seed) and washed three times with tap water and once with deionized water. These materials were blotted on filter paper. One part of plant material (plant fresh matter) was frozen using liquid nitrogen and stored a −30˚C, this material was used for antioxidant enzyme assays (SOD, CAT, APX and GSH-PX) and H2O2 quantification. The rest of the plant material (plant dry matter) was dried at 65˚C and used to determine the biomass, crop yield and Fe concentration in bean seed.
2.4. Biomass and Yield
Plant biomass was determined as the average dry weight of the entire plant and expressed as mg DW−1. Yield was expressed as the mean weight of fruits per plant in grams of dry weight.
2.5. Iron Quantification in Bean Seeds
Fe concentration was determined by an Induced Plasma Optical Emission Spectrometer (Agilent Technologies 700 Series ICP-OES) with a complete dissolution of 0.5 g of sample with 6 ml of concentrated HNO3 in an analytical microwave oven, START D (Milestone, Italy) at a temperature of 180˚C during 38 minutes. The resulting solution was diluted to 50 ml with deionized water and the metal concentration determined by ICP-OES. Fe concentration was expressed in mg∙kg−1 of dry weight [16] .
2.6. Activity of Antioxidant Enzymes
2.6.1. SOD Assay
SOD (EC 1.15.1.1) activity was determined by its ability to inhibit the formation of nitro blue formazan from NBT according to two methods [17] , with some modifications. Total soluble proteins were extracted from 0.5 g of frozen bean seeds ground in 5 ml of extraction buffer (50 mm of Hepes-HCl, at pH 7.6) with a mortal pestle. This homogenized was centrifuged at 11,500 rpm, for 20 min, at 4˚C; the supernatant liquid was separated by decantation and used for the enzymatic assay of SOD and total protein quantification. The supernatant was diluted at a proportion of 1:5 with the maceration buffer. For the SOD assay, it was used 0.1 ml de protein extract for each sample and mix with 5 ml enzymatic reaction buffer (50 mm Na2CO3, pH 10.0), 13 mm methionine, 0.025% v/v, triton X-100, 63 µmol NBT, 1.3 µM riboflavin). The reaction was illuminated at PPFD of 380 µmol∙m−2∙s−1, for 15 min. Reactions without illumination were used as a control to correct for background absorbance. One unit of SOD activity was defined as the amount of enzyme required to cause 50% inhibition of the reduction of NBT as monitored at 560 nm. SOD specific activity was expressed as U min−1∙g−1 FW. One unit of SOD was defined as the amount required inhibiting the photo reduction of NBT by 50%.
2.6.2. CAT Assay
CAT (EC 1.11.1.6) activity was determined by spectrophotometrically following H2O2 consumption at 240 nm [18] . Total soluble proteins were extracted from 0.5 g of frozen bean seeds ground in 5 ml of extraction buffer (25 mm of Hepes-HCl, at pH 7.8) with a mortal pestle. This homogenized was centrifuged at 11,500 rpm, for 20 min, at 4˚C. The supernatant liquid was separated by decantation and then this extract was used for the enzymatic assay of CAT and total protein quantification. CAT activity was determinate using the next reaction: 0.75 ml of Na2HPO4-NaH2PO4 buffer (25 mm, at pH 7), 0.75 ml de EDTA-Na2 (0.8 mm), 1 ml de H2O2 (20 mm) and 0.5 ml of protein extract. The absorbance change was measured at a wavelength of 240 nm, for 3 min. This enzymatic assay was performed at 25˚C. The results were expressed as specific activity (nmol H2O2 decomposed min−1∙mg−1 protein).
2.6.3. GSH-PX Assay
GSH-PX (EC 1.11.1.9) activity was measured using H2O2 as substrate [19] . Total soluble proteins were extracted from 0.5 g of frozen bean seeds ground in 5 ml of extraction buffer (25 mm of KNaHPO4, at pH 7) with a mortal pestle. This homogenized was centrifuged at 11,500 rpm, for 20 min, at 4˚C. The supernatant liquid was separated by decantation and used for the enzymatic assay of GSH-PX and total protein quantification. For the enzyme reaction, an aliquot of 0.2 ml of the protein extract was mixed with 0.4 ml GSH (0.1 mm) and 0.2 ml KNaHPO4 (0.067 M). This mixture was preheating on water bath at 25˚C, for 5 min. The above mention reagents without supernatant extract were used for the non-enzyme reaction. After preheating, 0.2 ml H2O2 (1.3 mm) was added to start the reaction. The reaction lasted 10 min and was stopped by adding 1 ml, 1% trichloroacetic acid and cooling at 4˚C, for 30 min. Then the mixture was centrifuged for 10 min at 11904 g. 0.48 ml of the supernatant was placed into a cuvette and 2.2 ml of 0.32 M Na2HPO4 and 0.32 ml of 1.0 mm DNTB were added for color development. The absorbance at wavelength 412 nm was measured after 5 min. The enzyme activity was calculated as a decrease in GSH within the reaction time when compared with that in the non-enzyme reaction.
2.6.4. Protein Assay
Concentration of total soluble proteins extracted of bean tissue, which were used for the 3 enzyme assays (CAT, SOD and GSH-PX), was determinate according to the manufacture’s protocol of Quick Start Bradford Protein Assay Kit (BioRad, USA). The standard protocol was performed with the next relation: 20 µl of sample and 1 ml of 1x Dye Reagent. The protein standard used for these protein assays was bovine serum albumin, provide in the kit (BSA Standard Set with 7 concentrations: 2, 1.5, 1, 0.75, 0.5, 0.25, 0.125 mg∙ml−1).
2.6.5. H2O2 Quantification
The H2O2 content of seed samples was colorimetrically measured [20] ). Seed samples were homogenized with 5 ml of cold acetone to extract the H2O2. An aliquot (1 ml) of this extract was mixed with 0.5 ml of titanium tetrachloride 20% (v/v) and 2.5 ml of NH4OH, centrifuged at 5000 g for 15 min and the supernatant was used to measure the absorbance at 415 nm (intensity of the yellow color). The H2O2 concentration was calculated according with Sánchez et al. [21] , using a standard curve of H2O2.
2.7. Statistical Analysis
Data were subjected to a simple ANOVA at 95% confidence, using SAS (SAS Institute Inc, Cary, NC). Means were compared by Tukey test (P ≤ 0.05). The data shown are mean values ± standard error (SE).
3. Results and Discussion
3.1. Effect of Fe Biofortification on Yield and Fe Accumulation in Bean Seed
Previous studies pointed out that Fe biofortification improves biomass and yield in legumes, such as soybean and beans [22] . It has also been found that when Fe is applied in the form of Fe-EDDHA is more effective in increasing biomass, that when this is applied in the form of FeSO4 in these plants [23] . Our results coincide with those studies, showing an improvement in the amount of biomass according Fe levels were increased (Figure 1, P ≤ 0.05) and yield increases notably when Fe-EDDHA form was applied (Figure 2, P ≤ 0.05). In this study,

Figure 1. Biomass in plants of P. vulgaris L. cv strike subjected to different iron doses and forms. Data are means ± SE (n = 4) (P ≤ 0.05).

Figure 2. Yield in plants of P. vulgaris L. cv strike subjected to different iron doses and forms. Data are means ± SE (n = 4) (P ≤ 0.05).
there were significant differences in the Fe application form. Fe-EDDHA form, at 50 and 100 µmol doses, were significantly different with respect to the control, obtaining an increase in yield of 1.5 fold (Figure 2, P ≤ 0.05).
Stress negatively affects growth and development in plants, genering ROS that damage macromolecules and cell structures. One of the most widely used indicators of stress is plant biomass [15] . Therefore, we studied this indicator of stress as a one way to define the effect of the different Fe treatments. In general, the results indicate that Fe-EDDHA is less toxic than FeSO4 because the plants tolerated and responded positively to Fe-EDDHA that even enhanced growth up to rate of 25, 50, 100 µM. While this effect was not detected for FeSO4, the increases of biomass and crop yield were similar at rates of 25, 50 and 100 µM (Figure 1, P ≤ 0.05).
Respect to Fe accumulation in seed, Fe biofortification induced the accumulation of this mineral in bean seed, with the two Fe forms applied (Figure 3, P ≤ 0.05). However, our results indicate that biofortification with Fe-EDDHA was more effective in improving Fe accumulation bean seed. The dose of Fe applied to the lowest level in concentration (25 µM) produced a highly significant increase in the accumulation of Fe in bean seed, increasing 29% the seed levels of Fe, with respect to the control (Figure 3, P ≤ 0.05). However, when Fe doses were applied at higher concentration (50 and 100 µM), we fail to increase the Fe accumulation in seeds (Figure 3,

Figure 3. Fe concentration in bean seeds subjected to different concentrations and forms of Fe. Data are means ± SE (n = 4) (P ≤ 0.05).
P ≤ 0.05). Therefore, we believe that these levels were toxic to these plants, so the plants did not accumulate more Fe in their seeds.
3.2. Effect of Fe Biofortification on Yield and Fe Accumulation in Bean Seed
Like other ROS, H2O2 accumulation is one of the causes of lipid peroxidation than attacks lipid membranes [24] . H2O2 is relatively “safe” in the absence of transition metals, being unreactive even at levels higher than biological system would ever generate. However, H2O2 is potentially quite reactive with molecules containing Fe2+ or other transition metals [25] . In previous studies of Se biofortification, it has been found that the content of H2O2 in seed can increased as increasing the doses of Se, which can be toxic levels and produce oxidative stress [15] . In this study, ours results shows a similar behavior with an increase in the H2O2 concentration as the Fe application rate increases (25, 50 and 100 µM of Fe). In addition, the H2O2 concentration detected is higher when the form of application used was FeSO4 (Figure 4, P ≤ 0.05). For these reason, we assumed than FeSO4 is more toxic than Fe-EDDHA in this bean crop, overall at a rate of 100 µM. In Figure 4, we can have appreciated than these increases of H2O2 levels were notably high at 50 µM dose of Fe-EDDHA and 100 µM dose of FeSO4, compared to the control. Fe-EDDHA at 50 and 100 µM doses had significant values of H2O2 in bean seed. Respect to FeSO4 form, it was an increase of H2O2 concentration in more than two fold, compared to the control. These results indicate that studied plants were more tolerant to Fe-EDDHA than FeSO4 improvement of stress tolerance to toxic levels of metals is often related to an increase in activity of antioxidant enzymes. Actually, early accumulation of H2O2 during metal exposition can signal an increase in antioxidant enzyme activities, which in turn protected plants from the oxidative stress caused by metals such as Fe. Thus, increased SOD, CAT and GSH-Pxactivities in bean seeds indicate that this specie of plant has the capacity to adapt to Fe toxicity by developing an antioxidant defense system.
CAT is a component of the plant antioxidant system, and controls ROS levels by removing the overproduction of H2O2 in their cells [26] . The effect of Fe application on CAT activity reported in this study, are in agreement with studies carried out in grasses plants. For example, it has been observed that doses of 50 and 100 µM of Fe applied to maize plants increased CAT activity in their seeds [27] . Our results are in line with this behavior, CAT activity shown an increase in bean seeds as the Fe doses were elevated, with both application forms of Fe (FeSO4, Fe-EDDHA). However, FeSO4 form produced a higher increase in the CAT activity of bean seed than Fe-EDDHA form, at the three doses used (Table 1, P ≤ 0.05).
SOD is an important member of the cell protective antioxidant system. This enzyme catalyzes the dismutation of the superoxide anion into H2O2 plus molecular oxygen [28] . Similar to CAT, we found that SOD activity was increasing in bean seeds as higher Fe dose were used (25 and 50 µM), with the two forms of Fe used. These increased resulted highly significant with respect to controls. However, we did not detect an increase of SOD activity at the highest dose of Fe used (100 µM), for both compounds of Fe studied (Table 1, P ≤ 0.05).

Figure 4. Hydrogen peroxide activity in seeds of P. vulgaris L. cv strike subjected to different iron doses and forms. Data are means ± SE (n = 4) (P ≤ 0.05).

Table 1. CAT, SOD and GSH-PX activities in bean seeds to different concentrations and forms of Fe.
CAT, catalase; SOD, superoxide dismutase; GSH-PX, glutathione peroxidase; FW, fresh weight. Data are means ± SE. Means followed by different letters in each column are significant at P < 0.05 (n = 4).
GSH-PX is another enzyme responsible of the detoxification of H2O2 produced by SOD, through transformation of superoxide ion [29] . Compared to the increase in enzymatic activity detected for CAT and SOD, we did not detect such behavior with the activity of GSH-PX in bean seeds, cultivated through biofortification with two forms of Fe. Regarding the form of Fe that was applied as FeSO4, we were unable to detect statically significant differences when we increased the dose of Fe at concentrations of 25, 50 and 100 µM (Table 1, P ≤ 0.05). For the case of Fe-EDDHA form, we registered a small decrease in GPX activity at doses of 50 and 100 µM (Table 1, P ≤ 0.05). In future projects, it would be necessary to confirm these results that focus on the study of another class of plant peroxidases, involved in this antioxidant system.
4. Conclusion
In conclusion, the increased SOD, CAT and GSH-PX activities in the bean seeds indicate that this specie has the capacity to adapt to Fe toxicity by developing an antioxidant defense system. In this study, the antioxidant system has significantly improved with respect to control, which can explain the beneficial effect of Fe found in plants subjected to diverse abiotic stress. In addition, we can indicate that the effect of Fe on this system depends largely on the form in which this trace element is applied. In bean seeds, application of FeSO4 produces a higher increase in the activity of the antioxidant enzymes than Fe-EDDHA form at the doses studied. The biofortification with Fe increases significantly the accumulation of Fe in bean seeds, and we considered that the best treatment is the application of Fe-EDDHA at 25 µM dose, which increases in high percentage of the levels of Fe in bean seeds and crop yield of this cultivar. In general, these results will contribute to define the utility and application of Fe biofortification, which promote the application of micronutrients at low rates. This technology has a great potential to control the induction of the antioxidant system in bean plants, thereby improving crop yield, stress resistance and accumulation of antioxidant compounds in bean seeds. Finally, to raise the Fe concentration in bean under biofortification program is a promising strategy in cropping systems in order to increase the ingestion of iron and antioxidant capacity in the general population and provide the benefits that this element offers in human health.
Acknowledgements
We appreciate the support provided by CONACyT to perform this work. This project was supported by FOMIX- CHIHUAHUA (CHIH-2010-C01-148114). Additionally, we thank the staff of the Laboratory of Plant Physiology and Nutrition of CIAD Delicias for their support to conduct the experiments of this study.
Cite this paper
Juan P.Sida-Arreola,EstebanSánchez,Graciela D.Ávila-Quezada,Paul B.Zamudio-Flores,Carlos H.Acosta-Muñiz, (2015) Can Improve Iron Biofortification Antioxidant Response, Yield and Nutritional Quality in Green Bean?. Agricultural Sciences,06,1324-1332. doi: 10.4236/as.2015.611127
References
- 1. Broughton, W., Hernandez, G., Blair, M., Beebe, S., Gepts, P. and Vanderleyden, J. (2003) Beans (Phaseolus spp.)— Model Food Legumes. Plant and Soil, 252, 55-128.
http://dx.doi.org/10.1023/A:1024146710611 - 2. Miklas, P.N., Kelly, J.D., Beebe, S.E. and Blair, M.W. (2006) Common Bean Breeding for Resistance against Biotic and Abiotic Stresses: From Classical to MAS Breeding. Euphytica, 147, 105-131.
http://dx.doi.org/10.1007/s10681-006-4600-5 - 3. García-Bañuelos, M.L., Sida-Arreola, J.P. and Sánchez, E. (2014) Biofortification Is a Promising Approach to Increase the Content of Iron and Zinc in Staple Food Crops. Journal of Elementology, 19, 865-888.
- 4. Cakmak, I., Pfeiffer, W.H. and McClafferty, B. (2010) Review: Biofortification of Durum Wheat with Zinc and Iron. Cereal Chemistry, 87, 10-20.
http://dx.doi.org/10.1094/CCHEM-87-1-0010 - 5. García-Bañuelos, M.L., Hermosillo-Cereceres, M.A. and Sánchez, E. (2011) The Importance of Selenium Biofortification in Food Crops. Current Nutrition and Food Science, 7, 181-190.
http://dx.doi.org/10.2174/157340111797264796 - 6. Hermosillo-Cereceres, M.A., Sánchez, E., Guevara-Aguilar, A., Muñoz-Márquez, E. and García-Bañuelos, M.L. (2013) Biofortification and Distribution Patterns of Selenium in Bean: Response to Selenate and Selenite. Journal of Food, Agriculture and Environment, 11, 421-426.
- 7. Aciksoz, S.B., Yazici, A., Ozturk, L. and Cakmak, I. (2011) Biofortification of Wheat with Iron through Soil and Foliar Application of Nitrogen and Iron Fertilizers. Plant and Soil, 349, 215-225.
http://dx.doi.org/10.1007/s11104-011-0863-2 - 8. Hänsch, R. and Mendel, R.R. (2009) Physiological Functions of Mineral Micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Current Opinion in Plant Biology, 12, 259-266.
http://dx.doi.org/10.1016/j.pbi.2009.05.006 - 9. Minotti, G. and Aust, S. (1987) The Requirement for Iron (III) in the Initiation of Lipid Peroxidation by Iron (II) and Hydrogen Peroxide. Journal of Biological Chemistry, 262, 1098-1104.
- 10. Elstner, E.F. and Osswald, W. (1994) Mechanisms of Oxygen Activation during Plant Stress. Proceedings-Royal Sciety of Edinburgh B, 102, 131-131.
- 11. Alscher, R.G., Donahue, J.L. and Cramer, C.L. (1997) Reactive Oxygen Species and Antioxidants: Relationships in Green Cells. Physiologia Plantarum, 100, 224-233.
http://dx.doi.org/10.1111/j.1399-3054.1997.tb04778.x - 12. Noctor, G. and Foyer, C.H. (1998) Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annual Review of Plant Biology, 49, 249-279.
http://dx.doi.org/10.1146/annurev.arplant.49.1.249 - 13. Vanlerberghe, G.C. and McIntosh, L. (1997) Alternative Oxidase: From Gene to Function. Annual Review of Plant Biology, 48, 703-734.
http://dx.doi.org/10.1146/annurev.arplant.48.1.703 - 14. Mittler, R. (2002) Oxidative Stress, Antioxidants and Stress Tolerance. Trends in Plant Science, 7, 405-410.
http://dx.doi.org/10.1016/S1360-1385(02)02312-9 - 15. Ríos, J., Blasco, B., Cervilla, L., Rosales, M., Sanchez-Rodriguez, E., Romero, L. and Ruiz, J. (2009) Production and Detoxification of H2O2 in Lettuce Plants Exposed to Selenium. Annals of Applied Biology, 154, 107-116.
http://dx.doi.org/10.1111/j.1744-7348.2008.00276.x - 16. Karacan, M.S. and Aslantas, N. (2008) Simultaneous Preconcentration and Removal of Iron, Chromium, Nickel with N-Etylenebis-(Ethane Sulfonamide) Ligand on Activated Carbon in Aqueous Solution and Determination by ICP-OES. Journal of Hazardous Materials, 155, 551-557.
http://dx.doi.org/10.1016/j.jhazmat.2007.11.107 - 17. Giannopolitis, C.N. and Ries, S.K. (1977) Superoxide Dismutases I. Occurrence in Higher Plants. Plant Physiology, 59, 309-314.
http://dx.doi.org/10.1104/pp.59.2.309 - 18. Rao, M.V., Paliyath, G., Ormrod, D.P., Murr, D.P. and Watkins, C.B. (1997) Influence of Salicylic Acid on H2O2 Production, Oxidative Stress, and H2O2-Metabolizing Enzymes (Salicylic Acid-Mediated Oxidative Damage Requires H2O2). Plant Physiology, 115, 137-149.
http://dx.doi.org/10.1104/pp.115.1.137 - 19. Flohé, L. and Günzler, W.A. (1984) Assays of Glutathione Peroxidase. Methods in Enzymology, 105, 114-120.
http://dx.doi.org/10.1016/S0076-6879(84)05015-1 - 20. Brennan, T. and Frenkel, C. (1977) Involvement of Hydrogen Peroxide in the Regulation of Senescence in Pear. Plant Physiology, 59, 411-416.
http://dx.doi.org/10.1104/pp.59.3.411 - 21. Sánchez, E., Soto, J.M., García, P.C., López-Lefebre, L.R., Rivero, R.M., Ruiz, J.M. and Romero, L. (2000) Phenolic Compounds and Oxidative Metabolism in Green Bean Plants under Nitrogen Toxicity. Functional Plant Biology, 27, 973-978.
http://dx.doi.org/10.1071/PP00008 - 22. Caliskan, S., Ozkaya, I., Caliskan, M. and Arslan, M. (2008) The Effects of Nitrogen and Iron Fertilization on Growth, Yield and Fertilizer Use Efficiency of Soybean in a Mediterranean-Type Soil. Field Crops Research, 108, 126-132.
http://dx.doi.org/10.1016/j.fcr.2008.04.005 - 23. Ortega-Blu, R. and Molina-Roco, M. (2007) Comparison between Sulfates and Chelated Compounds as Sources of Zinc and Iron in Calcareous Soils. Agrociencia, 41, 491-502.
- 24. Djanaguiraman, M., Devi, D.D., Shanker, A.K., Sheeba, J.A. and Bangarusamy, U. (2005) Selenium—An Antioxidative Protectant in Soybean during Senescence. Plant and Soil, 272, 77-86.
http://dx.doi.org/10.1007/s11104-004-4039-1 - 25. Becana, M., Moran, J. and Iturbe-Ormaetxe, I. (1998) Iron-Dependent Oxygen Free Radical Generation in Plants Subjected to Environmental Stress: Toxicity and Antioxidant Protection. Plant and Soil, 201, 137-147.
http://dx.doi.org/10.1023/A:1004375732137 - 26. Gupta, R., Kannan, G.M., Sharma, M. and Flora, S.J. (2005) Therapeutic Effects of Moringaoleifera on Arsenic-Induced Toxicity in Rats. Environmental Toxicology and Pharmacology, 20, 456-464.
http://dx.doi.org/10.1016/j.etap.2005.05.005 - 27. Sun, B., Jing, Y., Chen, K., Song, L., Chen, F. and Zhang, L. (2007) Protective Effect of Nitric Oxide on Iron Deficiency-Induced Oxidative Stress in Maize (Zea mays). Journal of Plant Physiology, 164, 536-543.
http://dx.doi.org/10.1016/j.jplph.2006.02.011 - 28. Slesak, I., Slesak, H., Libik, M. and Miszalski, Z. (2008) Antioxidant Response System in the Short-Term Post-Wounding Effect in Mesembryanthemum crystallinum Leaves. Journal of Plant Physiology, 165, 127-137.
http://dx.doi.org/10.1016/j.jplph.2007.03.015 - 29. Arora, A., Sairam, R. and Srivastava, G. (2002) Oxidative Stress and Antioxidative System in Plants. Current Science, 82, 1227-1238.
NOTES

*Corresponding author.