Agricultural Sciences
Vol.5 No.9(2014), Article ID:48426,8 pages
DOI:10.4236/as.2014.59083
Effects of Ce4+ on Membrane Integrity of Rice in Seedling Hydroponic Cultures
Qiuman Xu*, Qianqian Zhao, Chaoyu Zhao, Hong Chen
College of Life Science, Tianjin Key Laboratory of Animal and Plant Resistance, Tianjin Normal University, Tianjin, China
Email: *qmxu0929@126.com
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 30 April 2014; revised 28 June 2014; accepted 18 July 2014
ABSTRACT
Rare earth elements have been extensively used as micro-fertilizers for crops in China. To understand the potential benefits or damages of Ce4+ on rice, the effects of Ce4+ on viability and membrane integrity of rice were investigated under hydroponic cultures. It has been found that the cell viability, electrical conductivity, pH changes, and concentration of malonyl dialdehyde did not change obviously when the content of Ce4+ was 0.5 mg/L, but varied markedly when the contents of Ce4+ were raised to 10 mg/L or more, indicating that the Ce4+ at higher concentrations severely affected the cell membrane permeability.
Keywords: Ce4+, Membrane Integrity, Hydroponic Culture, Rice

1. Introduction
Rare earth elements (REEs) enriched fertilizers have been commonly used in China since the 1980s, thus inducing a growing concern about their environmental impact in agriculture. It has already been proved that REEs can improve the production and quality of crops. Recently, the research of crops has been studied at extensive aspects, including biological geotextiles [1] . The effects of REEs on plants have been studied at the physiological and biochemical level [2] -[5] , including the promotion of the growth of Crocus sativus cells, the cellular location of REEs in a cell, and reactive oxygen metabolism. Most of the evidences are focused on the REEs (III) ions. Less attention was paid to the mechanism of Ce4+ acting as a stimulating signal of cell membrane defense responses.
The defense responses of plant cells to pathogens include the activation of defense-related genes [6] , accumulation of phytoalexins, hypersensitive response and disposition of lignin and other phenolic compounds in cell wall [7] [8] . The activation of plant defense requires the recognition of specific endogenous or exogenous signal molecules and the transduction of these signals to the cell nucleus. The cell membrane acts as “signal sensors” for external stimuli. Most of environmental stresses activate membrane-bound G-protein and enzymes such as Ca-ATPase, H+-ATPase and NADPH oxidase [9] , resulting in early cell responses including ROS accumulation [10] [11] , extra-cellular medium acidification/alkalinization and subsequent defense gene activation. Therefore, the maintenance of cell-membrane integrity under stress conditions is essential not only for cells to transform external signals but also for them to survive.
Whether and how the Ce4+ affects the cell membrane permeability in seedlings hydroponic cultures is an issue worth investigating. As the cell membrane is an interface for cells to exchange with the outer environment, the external stimuli may change some cellular functions [12] .
In this work, the effects of Ce4+ on membrane integrity of seedlings hydroponic cultures of rice were investigated in detail so as to have a better understanding of the molecular mechanism of membrane responses. These will help us to understand the mechanism of Ce4+ on crops production changes and concern about its environmental impact in agricultures.
2. Materials and Methods
2.1. Cultural Procedure
The seeds of rice (Oryza sative. Jinxing1) were sterilized by H2O2 (10%) for 30 min and washed with deionized water, then soaked in water for 6 h and germinated in an incubator at 28˚C. When the length of hypocotyl was about 1 cm, seedlings were transplanted to plastic pots (diameter 10 cm) filled with deionized water in an incubator with the illumination of 6 klx (12 h light per day) at 28˚C. When the second leaves appeared, the seedlings were cultured in a 1/2 Hoagland nutrient solution. The nutrient solution was renewed every three days to stabilize pH. When the third leaves appeared, the seedlings were separately treated with different concentration of Ce4+ (0.5, 5, 10 mg/L) supplied as Ce(NH4)2(NO3)6. The control was treated with same amount of deionized water. Samples were collected for analyses at a predetermined time.
2.2. Seedlings Viability
Seedlings viability was assayed using 2,3,5-triphenyltetrazolium chloride (TTC) following the method of Iborra et al. [13] . Following Ce4+ treatment, the TTC solution was infiltrated into the rice cells. This assay directly determines the mitochondrial electron transport activity. The relative level of TTC reduction to formazan quantifies the cell viability by the spectrophotometric assay of the red formazan [13] . Fresh seedlings (200 mg) were collected in a tube and 8 ml TTC solution (0.8% TTC in 0.05 M phosphate buffer, pH 7.4, and 0.5 mL/L Tween 20) were added and the mixture was infiltrated for 5 min at reduced pressure. The seedlings samples were incubated in the TTC solution for 24 h at 25˚C in the dark, then seedlings were removed and rinsed with distilled water, placed in separate quartz tubes containing 3 ml of 95% ethanol in each, and submerged for 24 h at 25˚C in the dark. The absorbance was measured at 485 nm. Cell viability was expressed as the absorbance of the sample at 485 nm per gram of biomass (fresh wt.).
2.3. pH Measurement
pH measurement was performed by the pH meter (Radiometer Copenhagen, PHM210, Tacussel, France).
2.4. Measurement of Electrical Conductivity
Rice seedlings were subjected to the same concentrations of Ce4+ as in the electrolyte leakage assay. The electrolyte leakage was quantified by electrical conductivity measurement. Membrane stability was expressed in terms of the electrolytic conductivity [14] . This method measures the increased electrolyte diffusion induced by Ce4+ as a result of the changed cell membrane permeability. A total of 200 mg seedlings were transferred from the Ce4+-treated samples to a 10 ml centrifuge tube containing 4 ml ultrapure water at predetermined time of cultivation. Absolute electrical conductivity (uS cm−1) was measured at 25˚C using a Model DDS-307 conductivity meter.
2.5. Assay of Membrane Lipid Peroxidation
Malonyl dialdehyde (MDA), a final product of lipid peroxidation, was measured to evaluate the extent of lipid peroxidation in seedling hydroponic culture of rice. The MDA was measured by thiobarbituric acid (TBA) reaction [15] . Briefly, seedlings (200 mg) were homogenized in 4 ml 0.1% trichloroacetic acid (TCA). The homogenate was centrifuged at 13,000 g for 10 min. To 2 ml aliquot of the supernatant was added 4 ml 20% TCA containing 0.5% TBA. The mixture was heated at 95˚C for 30 min and quickly cooled down in an ice bath. After centrifugation at 10,000 g for 10 min, the absorbance of the supernatant was measured at 532 nm and the unspecific turbidity was exempted by subtracting the absorbance of the same sample at 600 nm. MDA concentration was calculated using the molar extinction coefficient of 155 mM−1·cm−1.
2.6. Evans Blue Staining
Cell membrane permeability was analyzed using Evans blue dye following the method of Suzuki et al. [16] with a slight modification. Seedlings (200 mg) washed three times with 50 mM phosphate buffer (pH 5.8); seedlings were then placed on the sterilized dry filter paper. After doing so, the seedlings were collected in a tube and stained for 5 min with a 0.15% (w/v) solution of Evans Blue. Afterwards, the seedlings were re-filtered and washed five times with PBS to remove the excess dyes. The stained seedlings were collected and re-suspended in 5 ml solution consisting of 1.0% (w/v) SDS and 50% (v/v) methanol for 30 min at 50˚C. The solution was then cooled down to room temperature and the supernatant was collected and its absorbance at 600 nm was measured.
3. Results
3.1. Effects of Ce4+ on Cell Viability and Electrical Conductivity in Seedlings
Figure 1 showed that the seedlings cell viability was hardly affected during the whole period when the concentration of Ce4+ was below 0.5 mg/L. However, the cell viability began to decreased rapidly after 2 days in the case of 5 mg/L Ce4+ and decreased 49.5% in the case of rice of that of control on Day 6. Further increase of Ce4+ (above 10 mg/L) reduced the cell viability, while the cell viability decreased 65.2% in the case of rice on day 8 after the addition of the Ce4+.
Figure 2 displays the time course of the conductivity with the addition of Ce4+. The conductivity was hardly affected compared to that of the control culture when the Ce4+ content was 0.5 mg/L. When the Ce4+ content was raised to 10 mg/L, however, the conductivity began to increase rapidly after 2 days and increased 693.3% in the case of rice of that of the control on Day 4, indicating that a net outflow of ions from the treated cells.
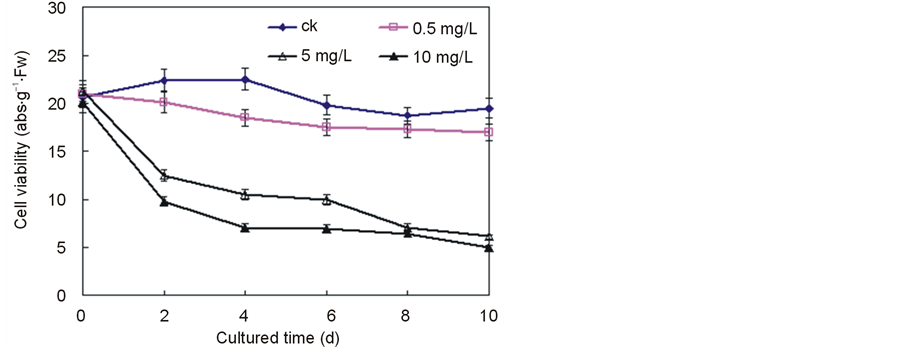
Figure 1. Effect of Ce4+ on cell viability in the seedling hydroponic cultures of rice. Cell viability was determined by the TTC method and expressed as the absorbance at 485 nm per gram of biomass (fresh weight). The data were the average of triplicate experiments. The errors bars indicate SD. CK, 0 mg/L Ce4+. (Xu et al., Effects of Ce4+ on membrane integrity…)
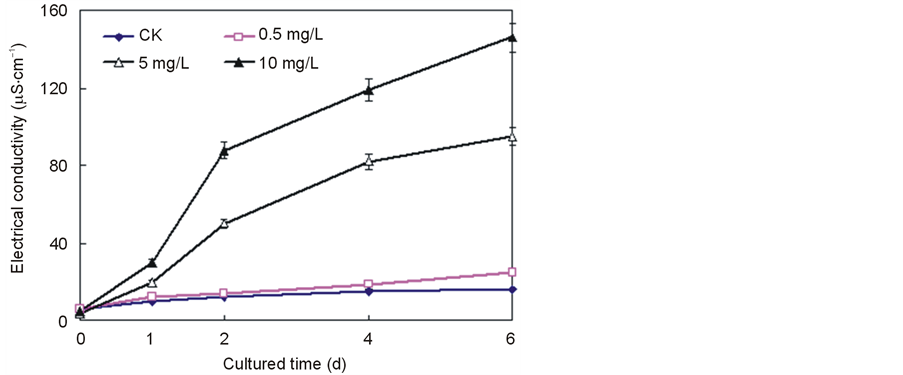
Figure 2. Effect of Ce4+ on electrical conductivity (EC) in the seedlings hydroponic cultures of rice. The data were the average of triplicate experiments. The errors bars indicate SD. CK, 0 mg/L Ce4+. (Xu et al., Effects of Ce4+ on membrane integrity…)
3.2. Membrane Lipid Peroxidation Induced by Ce4+Addition
To further confirm the changes in cell membrane permeability caused by the Ce4+ addition, the peroxide product of the main membrane lipids as a marker of membrane lipid breakage, the production of MDA, was tested. Figure 3 showed that MDA was induced by Ce4+. When the content of Ce4+ was 0.5 mg/L, the accumulation of MDA was slight. However, the MDA content increased 123.1% of that of the control on Day 5 after the addition of 10 mg/L Ce4+ and reached 158.3% in the case of rice on Day 7.
3.3. Changes of Extra-Cellular Medium pH
Figure 4 shows the dynamic changes of medium pH induced by Ce4+. As shown in Figure 4, upon the addition of Ce4+ the extra-cellular medium pH sharply decreased to a maximum value then began to increase and gradually restored to its initial level in the case of rice. The increase in Ce4+ content led to a severer acidification of the extra-cellular medium. For example, when Ce4+ concentration was 0.5 mg/L, no obvious change in extracellular pH was observed. However, when Ce4+ concentration was raised to 10 mg/L, the extra-cellular pH decreased 1.0 unit compared to that of the control sample. At Day 2, all culture media returned to its initial level. These results indicated that Ce4+ might take a function of inhibiting proton influx into cells across plasma membrane.
3.4. Changes of Membrane Permeability by Evens Blue Staining
Integrated cell membrane is impermeable for macromolecules such as Evans blue. When cell membrane undergoes an injury or loses its selective permeability, macromolecules can penetrate it. Thus, the amount of Evans blue entering and binding on cells reflects the degree of loss of cell membrane integrity. The change in cell membrane permeability was also reflected by the Evans blue assay. When the content of Ce4+ was 0.5 mg/L, it was evident from Figure 5 that the cell membrane integrity was well maintained as normal. However, when the content of Ce4+ was higher than 5 mg/L, an obvious loss in cell membrane integrity was observed. These results clearly indicated that Ce4+ broke the membrane integrity of rice cells when their content reached 5 mg/L or more.
4. Discussion
The treatment of Ce4+ (5 mg/L and 10 mg/L) led to a significant increase in ion leakage and decrease in rice seedlings cell viability compared to those of the control. Higher lipid peroxidation has been correlated with membrane permeability and electrolyte leakage [17] (Figure 2, Figure 3). To confirm it, membrane lipid
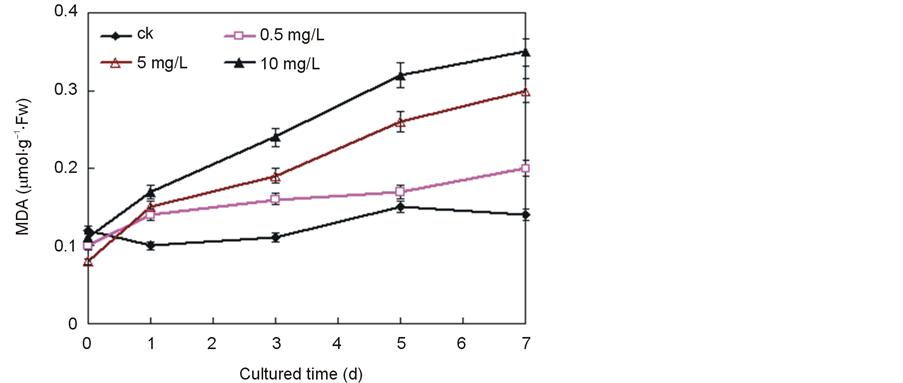
Figure 3. Effect of Ce4+ on membrane lipid peroxidation as reflected by the accumulation of malonyl dialdehyde (MDA) in the seedling hydroponic cultures of rice. The data were the average of triplicate experiments. The errors bars indicate SD. CK, 0 mg/L Ce4+. (Xu et al., Effects of Ce4+ on membrane integrity…)
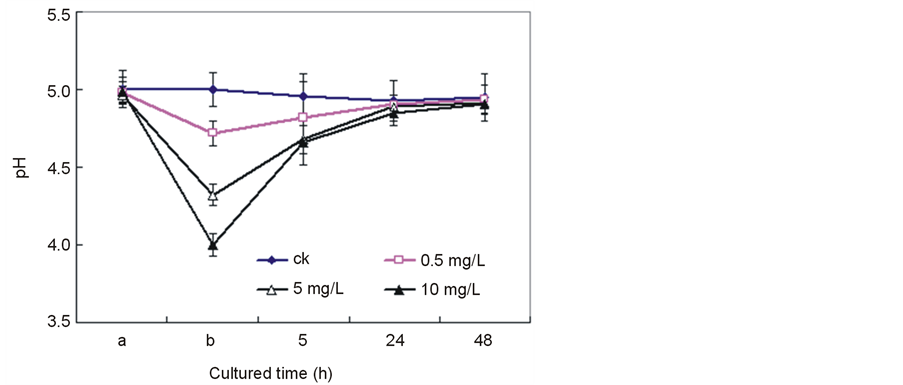
Figure 4. Effect of Ce4+ on pH in the seedling hydroponic cultures of rice. The data were the average of triplicate experiments. The errors bars indicate SD. CK, 0 mg/L Ce4+. (a) The pH value of culture media before adding Ce4+; (b) The pH value of culture media immediately detected after adding Ce4+. (Xu et al., Effects of Ce4+ on membrane integrity…)
peroxidation caused by Ce4+ was studied (Figure 3). The development of any adverse processes is associated with disorders in cell metabolisms, structures and disintegration of the barrier and structural functions of the cell membrane [18] . The state of cell membranes is one of the most important factors in regulating biochemical and physical processes and maintaining homeostasis in cells.
Lipid peroxidation is considered to be a process induced by free radicals [19] . Increased lipid peroxidation has been known to occur during senescence [20] [21] . When the content of Ce4+ was over 5 mg/L, the lipid peroxidation markedly increased (Figure 3). This may reflect a decline of the anti-oxidative enzymes [22] . Lipid membrane peroxidation might alter the cell membrane permeability resulting in solute leakage, membrane damage and consequently a loss of cellular physiological functions. The results here showed that the lipid peroxidation increased with increasing culture time and Ce4+ content.
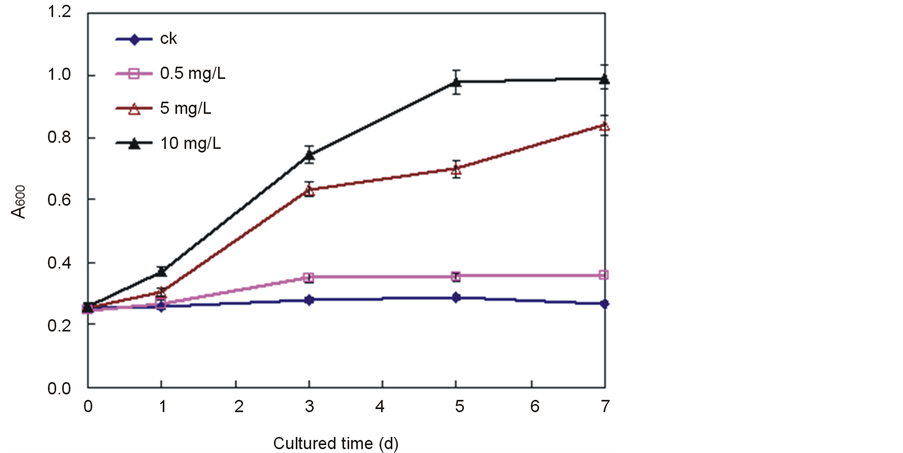
Figure 5. Effect of Ce4+ on the membrane permeability as demonstrated by Evans blue staining of rice in the seedling hydroponic cultures. The data were the average of triplicate experiments. The errors bars indicate SD. CK, 0 mg/L Ce4+. (Xu et al., Effects of Ce4+ on membrane integrity…)
Generally, the extraor intracellular pH maintains constant at normal physiological conditions. The pH change is regarded as a mechanism of cell biochemical responses under environment stresses [23] . Figure 4 displays the dynamic changes of medium pH induced by Ce4+. The H+-ATPase on the plasma membrane is a key enzyme responsible for H+ extrusion. The activation and inhibition of H+-ATPase is one of the possible mechanisms of the acidification or alkalinization of extra-cellular medium caused by various factors [24] . Therefore, the acidification of extra-cellular medium might be due to the activation of Ce4+ on H+-ATPase as it went on (Ca2+-Mg2+)-ATPase [25] . As the stabilization of cellular medium pH is essential for cells to perform their biological functions, cells have other mechanisms to protect themselves from the unfavorable conditions. The restoration of the medium pH to its initial level might show that the alkaline plasma environment in turn inhibited H+-ATPase.
Cell membrane is considered to be the primary site of toxicity for cyclic hydrocarbons with hydrocarbons preferentially partitioning into the lipid-containing plasmalemma from the aqueous media due to their hydrophobic nature and likely accumulating in the central, aliphatic part of the membrane bilayer [26] [27] . Recently, Xia et al. reported that the low level of Ce(III) had little effect on the mitochondrial permeability transition (MPT) of rice MPT, however, the higher level of Ce(III) could induce rice MPT [28] . As the initial site of interaction, the cell membrane disintegrated with increasing the content of Ce4+ to the limit (5 mg/L) that the cells could suffer.
5. Conclusion
The treatment with Ce4+ severely resulted in the decreases of cell viability, especially with 5 mg/L and 10 mg/L. Meanwhile, the pH of culture media dramatically decreased after adding the Ce4+, indicating the Ce4+ addition causes the acidification of the extracellular medium. Additionally, the Ce4+ addition (5 mg/L or more) caused the malonyl dialdehyde accumulation. The Evans blue assay further proofed that Ce4+ broke the membrane integrity of rice cells when their content reached 5 mg/L or more. Taken together, a higher content (5 mg/L or higher) remarkably impacted the membrane integrity of rice cells in seedling hydroponic cultures.
Acknowledgements
The authors are grateful for the financial support from the Natural Science Foundation of Tianjin (program: 13JCYBJC25500).
References
- Bhattacharyya, R., Zheng, Y., Li, Y.M., Tang, L., Panomtaranichagul, M., Peukrai, S., Thu, D.C., Cuong, T.H., Toan, T.T., Jankauskas, B., Jankauskiene, G., Fullen, M.A., Subedi, M. and Booth, C.A. (2012) Effects of Biological Geotextiles on Aboveground Biomass Production in Selected Agro-Ecosystems. Field Crops Research, 126, 23-36.http://dx.doi.org/10.1016/j.fcr.2011.09.006
- Tyler, G. (2004) Rare Earth Elements in Soil and Plant Systems—A Review. Plant and Soil, 267, 191-206.http://dx.doi.org/10.1007/s11104-005-4888-2
- d’Aquino, L., de Pinto, M.C., Nardi, L., Morgana, M. and Tommasi, F. (2009) Effect of Some Light Rare Earth Elements on Seed Germination, Seedling Growth and Antioxidant Metabolism in Triticum durum. Chemosphere, 75, 900-905. http://dx.doi.org/10.1016/j.chemosphere.2009.01.026
- Huang, S.F., Li, Z.Y., Wang, X.Q., Wang, Q.X. and Hu, F.F. (2010) Cerium Caused Life Span Shortening and Oxidative Stress Resistance in Drosophila melanogaster. Ecotoxicology and Environmental Safety, 73, 89-93.http://dx.doi.org/10.1016/j.ecoenv.2009.09.017
- Xu, Q.M. and Chen, H. (2011) Antioxidant Responses of Rice Seedling to Ce4+ under Hydroponic Cultures. Ecotoxicology and Environmental Safety, 74, 1693-1699. http://dx.doi.org/10.1016/j.ecoenv.2011.04.005
- van der Hoorn, R.A. and Kamoun, S. (2008) From Guard to Decoy: A New Model for Perception of Plant Pathogen Effectors. Plant Cell, 20, 2009-2017. http://dx.doi.org/10.1105/tpc.108.060194
- Pennell, R.I. and Lamb, C. (1997) Programmed Cell Death in Plants. Plant Cell, 9, 1157-1168.http://dx.doi.org/10.1105/tpc.9.7.1157
- Koch, W., Wagner, C. and Seitz, H.U. (1998) Elicitor-Induced Cell Death and Phytoalexin Synthesis in Daucus carrota L. Planta, 206, 523-532. http://dx.doi.org/10.1007/s004250050429
- Chanvitayapongs, S., Draczynska, L.B. and Sun, A.Y. (1997) Amelioration of Oxidative Stress by Antioxidants and Resveratrol in PC12 Cell. Neuroreport, 8, 1499-1502. http://dx.doi.org/10.1097/00001756-199704140-00035
- Ippolito, M.P., Fasciano, C., d’Aquino, L., Morgana, M. and Tommasi, F. (2010) Responses of Antioxidant Systems after Exposition to Rare Earths and Their Role in Chilling Stress in Common Duckweed (Lemna minor L.): A Defensive Weapon or a Boomerang? Archives of Environmental Contamination and Toxicology, 58, 42-52.http://dx.doi.org/10.1007/s00244-009-9340-9
- Ruiz-Sánchez, S.M., Aroca, R., Munoz, Y., Polon, R. and Ruiz-Lozano, J.M. (2010) The Arbuscular Mycorrhizal Symbiosis Enhances the Photosynthetic Efficiency and the Antioxidative Response of Rice Plants Subjected to Drought Stress. Journal of Plant Physiology, 167, 862-869. http://dx.doi.org/10.1016/j.jplph.2010.01.018
- Goodman, E.M., Greenebaum, B. and Marron, M.T. (1995) Effects of Electromagnetic Fields on Molecules and Cells. International Review of Cytology, 158, 279-338. http://dx.doi.org/10.1016/S0074-7696(08)62489-4
- Iborra, J.L., Guardiola, J., Montaner, S., Canovas, M. and Manjon, A. (1992) 2,3,5-triphenyltetrazolium Chloride as a Viability Assay for Immobilized Plant Cells. Biotechnology Techniques, 6, 319-322. http://dx.doi.org/10.1007/BF02439319
- Saadalla, M.M., Shanahan, J.F. and Quick, J.S. (1990) Heat Tolerance in Winter Wheat: I. Hardening and Genetic Effects on Membrane Thermostability. Crop Science, 30, 1243-1247. http://dx.doi.org/10.2135/cropsci1990.0011183X003000060017x
- Heath, R.L. and Packer, L. (1968) Photoperoxidation in Isolate Chloroplasts: I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Archives of Biochemistry and Biophysics, 125, 189-198. http://dx.doi.org/10.1016/0003-9861(68)90654-1
- Suzuki, K., Yano, A. and Shinshi, H. (1999) Slow and Prolonged Activation of the p47 Protein Kinase during Hypersensitive Cell Death in a Culture of Tobacco Cells. Plant Physiology, 119, 1465-1472. http://dx.doi.org/10.1104/pp.119.4.1465
- Dhindsa, R.S., Plumb-Dhindsa, P. and Thorpe, T.A. (1981) Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. Journal of Experimental Botany, 32, 93-101. http://dx.doi.org/10.1093/jxb/32.1.93
- Gavrilov, V.B., Kravchenko, O.N. and Konev, S.V. (2000) Dye Sorption as an Indicator of Erythrocyte Membrane Damage and Prehemolytic State of Erythrocytes. Bulletin of Experimental Biology and Medicine, 129, 358-360.http://dx.doi.org/10.1007/BF02433915
- Slater, T.F. (1984) Free-Radical Mechanisms in Tissue Injury. Biochemical Journal, 222, 1-15.
- Paliyath, G. and Droillard, M.J. (1992) The Mechanisms of Membrane Deterioration and Disassembly during Senescence. Plant Physiology and Biochemistry, 30, 789-812.
- Xu, Q.M., Cheng, J.S., Ge, Z.Q. and Yuan, Y.J. (2004) Effects of Organic Solvents on Membrane of Taxus cuspidata Cells in Two-Liquid-Phase Cultures. Plant Cell, Tissue and Organ Culture, 79, 63-69. http://dx.doi.org/10.1007/s11240-004-4709-y
- Xu, Q.M., Cheng, J.S., Ge, Z.Q. and Yuan, Y.J. (2005) Antioxidant Responses to Oleic Acid in Two-Liquid-Phase Suspension Cultures of Taxus cuspidata. Applied Biochemistry and Biotechnology, 125, 11-26. http://dx.doi.org/10.1385/ABAB:125:1:011
- Serrano, R. (1989) Structure and Fuction of Plasma Membrane ATPase. Annual Review of Plant Physiology and Plant Molecular Biology, 40, 61-94. http://dx.doi.org/10.1146/annurev.pp.40.060189.000425
- Schaller, A. and Oecking, C. (1999) Modulation of Plasma Membrane H+-ATPase Activity Differentially Activates Wound and Pathogen Defense Responses in Tomato Plants. Plant Cell, 11, 263-272.
- McConnell, E.J., Wagoner, M.J., Keenan, C.E. and Raess, B.U. (1999) Inhibition of Calmodulin-Stimulated (Ca2+ + Mg2+)-ATPase Activity by Dimethyl Sulfoxide. Biochemical Pharmacology, 57, 39-44. http://dx.doi.org/10.1016/S0006-2952(98)00259-7
- Sikkema, J., de Bont, J.A.M. and Poolman, B. (1994) Interactions of Cyclic Hydrocarbons with Biological Membranes. Journal of Biological Chemistry, 269, 8022-8028.
- Sikkema, J., de Bont, J.A. and Poolman, B. (1995) Mechanisms of Membrane Toxicity of Hydrocarbons. Microbiological Reviews, 59, 201-222.
- Xia, C.F., Zhao, J., Jin, J.C., Yuan, L., Chen, X.Y., Peng, W., Jiang, F.L., Qin, C.Q., Dai, J. and Liu, Y. (2013) Ce(III)-Induced Rice Mitochondrial Permeability Transition Investigated by Spectroscopic and Microscopic Studies. Biological Trace Element Research, 152, 284-291. http://dx.doi.org/10.1007/s12011-013-9621-z
NOTES

*Corresponding author.