American Journal of Plant Sciences
Vol.06 No.10(2015), Article ID:57604,12 pages
10.4236/ajps.2015.610166
Induced Systemic Resistance in Two Genotypes of Brassica napus (AACC) and Raphanus oleracea (RRCC) by Trichoderma Isolates against Sclerotinia sclerotiorum
Jawadayn Talib Alkooranee1,2,3, Tamarah Raad Aledan1, Jun Xiang4, Guangyuan Lu5*, Maoteng Li1,3*
1College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan, China
2Plant Protection Department, College of Agriculture, University of Basrah, Basrah, Iraq
3Hubei Key Laboratory of Economic Forest Germplasm Improvement and Resources, Huanggang, China
4Hubei Collaborative Innovation Center for the Characteristic Resources Exploitation of Dabie Mountains, Huanggang, China
5Oil Crops Research Institute, Chinese Academy of Agricultural Sciences, Wuhan, China
Email: *luwiz@163.com, *limaoteng426@mail.hust.edu.cn
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 16 May 2015; accepted 27 June 2015; published 30 June 2015
ABSTRACT
Two different species, Trichoderma viride TV10 and Trichoderma harzianum TH12 from 30 Tricho- derma isolates were selected out based on their high growth inhibition of the phytopathogen Sclerotinia sclerotiorum (Lib) de Bary, which reached 84.44% and 100%, respectively. Their untreated culture filtrates (CF) and culture filtrates treated with heat (CFH) also were tested for growth inhibition of the pathogen in potato dextrose agar (PDA). Morphological and molecular characterisation by internal transcribed spacer (ITS) PCR provided consistent identification of these isolates. The degree of infection and disease index (DI) of S. sclerotiorum were examined in Brassica napus (AACC) and Raphanus alboglabra (RR) and Brassica alboglabra (CC). The results revealed that Raphanus alboglabra showed higher disease resistance than that of B. napus. Biotic elecitors T. harzianum TH12 and T. viride TV10 and their CF and CFH demonstrated the ability to cause induced systemic resistance (ISR) in B. napus and Raphanus alboglabra against sclerotinia stem rot (SSR) disease. Furthermore, a high ability to reduce the degree of infection and DI in B. napus with the biotic elicitors T. harzianum TH12 and T. viride TV10 was observed, with numbers reaching 7.22% to 6.67% and 17.78% to 11.67%, respectively. When CF were used, reached 20.00% to 16.67% and 33.33% to 23.33%, respectively; with CFH, values reached 35.00% to 21.67% and 37.78% to 28.33%, respectively. While in Raphanus alboglabra the degree of infection and DI reached 0.00% and 0.00% with all biotic elicitors treatments. These results show that biotic elicitor treatments significantly (P < 0.05) can increase the fresh and dry weights of both roots and shoots as well as plant height compared with controls. The TH12 treatment in B. napus and Raphanus alboglabra ranked as most effective. This study showed for the first time the ability of genotype Raphanus alboglabra (RRCC) to demonstrate resistance against S. sclerotiorum with or without treatment by biotic elicitors and the ability of genotype B. napus (AACC) to demonstrate resistance to the pathogen after treatment with biotic elicitors.
Keywords:
Brassica napus, Raphanus alboglabra, Induced Systemic Resistance, Sclerotinia sclerotiorum, Trichoderma spp.

1. Introduction
The Brassicaceae family is composed of many of the economically important food crops. Brassica napus is one of the most important oilseed crops in the world after soybean and palm, as it provides oils for human consumption and is used as a forage for animal feed, biofuels, and soil conditioners [1] [2] . B. napus (AACC, 2n = 38) is an allopolyploid, resulting from the natural hybridisation between B. rapa (AA, 2n = 20) and B. oleracea (CC, 2n = 18) [3] . Raphanus alboglabra (2n = 34) was a new materials that obtained by hybridization between Raphanus sativus (RR, 2n = 16) and B. alboglabra (CC, 2n = 18) [4] [5] .
Sclerotinia stem rot (SSR) disease is the most common diseases of canola plants caused by Sclerotinia sclerotiorum (Lib) de Bary, and its economical effects are associated with substantial losses in quality and quantity of seed yield [6] [7] . S. sclerotiorum is a necrotrophic pathogen that could cause disease in more than 400 host plants, and it can cause white mold disease if the conditions are suitable. Although some fungicides have been available to manage this disease, but, several negative effects on the environment have been observed. Excessive use and misuse of agrochemicals may lead to serious environmental consequences, which have reinforced the necessity to consider alternative strategies for the management of this plant disease [8] [9] .
There are a variety of induced resistances that vary according to different signalling. Systemic acquired resistance (SAR) and induced systemic resistance (ISR) are two forms of systemic resistance. In both SAR and ISR, plant defenses are released by previous infection thus the application of biotic and abiotic factors, with most agents urged to reduce disease in infected plants between 20% and 85%. SAR and ISR can be used as alternatives to fungicides [10] . ISR is mediated by the NPR1 gene, which is a key gene involved in disease resistance, and is phenotypically similar to SAR. However, the molecular pathways of each are different; SAR depends on the salicylic acid (SA) responsiveness [11] , while ISR is dependent upon two pathways that respond to ethylene and jasmonic acid [12] . ISR can be defined by induction of defences in plants against many pathogens by application of plant growth-promoting microorganisms in the soil as well as by direct spreading on plants [13] [14] . Increased resistance in B. rapa and Arabidopsis thaliana leaves to the fungal pathogen Colletotrichum higginsianum and bacterial pathogen Pseudomonas syringae pv. Maculicola when treated with the plant activator Housaku Monogatari (HM) prepared from cell wall extracts of yeast appears to induce early activation of jasmonate/ethylene and late activation of SA pathways [15] .
Trichoderma spp. are soil-borne and produce green spores, and they are among ascomycetes that are widespread throughout the world [16] . Fungi of the genus Trichoderma, which comprise a group plant growth-pro- moting fungi (PGPF), are able to colonise the intercellular parts of plant roots and stimulate systemic resistance in all parts of the plant, as demonstrated in cucumber [17] . The ability of Trichoderma spp. to induce systemic resistance in dicotyledonous and monocotyledonous plants has been demonstrated against different pathogens, including the fungus S. sclerotiorum in tomato and cauliflower [18] , the bacteria Ralstonia solanacearum in tomato [19] , the cucumber mosaic virus in A. thaliana [20] , and the nematode Meloidogyne incognita in Vigna radiate [21] . The fungus T. viride can induce systemic resistance against Fusarium oxysporum and Alternaria spp. In addition, in black gram under greenhouse conditions, plants pre-treated with T. viride showed higher levels of antioxidant defensive enzymes that led to enhanced seed germination, increased growth, and decreased disease incidence compared with plants without treatment [22] . T. harzianum T12 spore suspension reduced the severity of disease of the bean rust fungus Uromyces appendiculatus even when autoclaved, and the T12 application on CF had protective but no curative effects [23] . At present, the effects of Trichoderma isolates on S. sclerotiorum in B. napus have not been reported.
The objectives of this study were to determine the ability and effect of the biotic elicitors T. harzianum and T. viride as well as CF and CFH on the pathogen S. sclerotiorum on the following:
Inhibited pathogen growth in a Petridish, onather is ISR in genotypes B. napus and Raphanus alboglabra against S. sclerotiorum-caused SSR disease.
2. Materials and Methods
2.1. Experimental Layout and Design
One spring-type B. napus genotype and newly obtained Raphanus alboglabra were used as plant materials in this experiment and isolates of the fungus S. sclerotiorum were obtained from the College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan, China. We chose these genotypes for comparison where B. napus is sensitive to sclerotinia stem rot (SSR) while we test Raphanus alboglabra for the first time against pathogens to know its ability to resist. Both genotypes of plants materials and S. sclerotiorum were used for greenhouse experiments during the entire period of investigation. The S. sclerotiorum fungus was maintained and cultured on PDA medium (200 g peeled potato, 20 g dextrose, 15 g agar, and 1 liter distilled water) in the dark at 22˚C ± 2˚C, and 5-mm-diameter mycelia agar plugs were punched fromthe growing margin after4 days. For rhizosphere inoculation, ascospores were harvested in sterile distilled water using a sterile brush and filtered through four layers of cheesecloth to remove the mycelia of the culture. The resulting spore suspension was adjusted to 1 × 106 spores/ml.
2.2. Biotic Elicitor Isolation
The study was conducted with 30 isolates of T. harzianum (TH1 to TH13) and T. viride (TV1 to T17) collected in 2014 from soil samples originating from rapeseed fields in Wuhan, Hubei province of China (114˚25'39.7"E; 30˚30'39.8"N). Samples were collected into polythene bags, labelled, and stored in the refrigerator until further processing. Trichoderma isolates were isolated in a specific selective medium containing the following (g/litre): 1.0 Ca (NO3)2, 0.26 KNO3, 0.26 MgSO4∙7H2O, 0.12 KH (PO4), 1.0 CaCl2, 0.05 C6H8O7 (citric acid); 2.0 sucrose, 20.0 agar, 0.05C22H23ClN2O8 (chlortetracycline), and 0.04 captan (50% wettable powder) [24] .
2.3. Morphological Identification of Trichoderma
Trichoderma spp. isolates were tentatively identified on the basis of their morphological characteristics [25] . All measurements of morphological characters (size and shape of conidia, size of conidiophores, and presence or absence of sterile hairs) were taken from slide mounts prepared using the tape touch method in a drop of lacto fuchsin [26] . Plates were incubated at 28˚C ± 2˚C, and colonies were selected. T. harzianum TH12 and T. viride TV10 were obtained from 30 isolates from the pure fungi cultures in these experiments.
2.4. PCR Amplification of ITS Regions of Trichoderma Isolates
Total DNA was extracted according to the method described by Chakraborty et al. [27] , and the DNA samples were stored at −20˚C in small aliquots. The ITS regions were amplified using the universal primers ITS1 (5’-TCTGTAGGTGAACCTGCGG-3’) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) according to White et al. [28] . PCR was performed in a total volume of 100 μl, which contained 78 μl deionised water, 10 μl 10× Taq pol buffer, 1 μl 1-U Taq polymerase enzyme, 6 μl 2-mM dNTPs, 1.5 μl 100-mM reverse and forward primers, and 1 μl 50-ng template DNA. PCR was programmed with an initial denaturing at 94˚C for 5 min followed by 30 cycles of denaturation at 94˚C for 30 sec, annealing at 59˚C for 30 sec, extension at 70˚C for 2 min, and a final extension at 72˚C for 7 min. The PCR product (20 μl) was mixed with loading buffer (8 μ1) containing 0.25% bromophenol blue and 40% (w/v) sucrose in water and then loaded in 2% agarose gel with 0.1% ethidium bromide for examination via horizontal electrophoresis.
2.5. Production of Spore Suspensions, CF, and CFH
Fifteen mycelial 1-cm square disks of actively growing TH12 and TV10 were inoculated separately with 300 ml of PDB and incubated with a rotary shaker (85 rpm) at 25˚C ± 2˚C. After 20 days, the mycelial mat was harvested and grinded to form a spore suspension. The cell concentration was then adjusted to 1.5 × 107 CFU/ml (TH12) and 1 × 107 CFU/ml (TV10). Pure cultures of biotic elicitors were maintained on respective agar slants and stored at 4˚C for further use. Cell-free CF from 20-day-old T. harzianum TH12 and T. viride TV10 grown on PDB were prepared by centrifugation (12,000 × g for 15 min) followed by filter sterilisation with a 0.4-μm filter unit; the supernatants were collected and used as enzyme solutions. Some CF for biotic elicitors were heated to 100˚C for 15 min (i.e., CFH) to collect non-volatile metabolites produced by biotic elicitors.
2.6. Assay of Mycoparasitic and CF Effects
To determine the effects of isolates on mycelia growth of the targeted pathogen S. sclerotirum in dual-culture techniques, Trichoderma isolate suspensions and their cell free (CF) and CFH biotic elicitors were added to molten PDA media (40˚C ± 3˚C) to obtain a final concentration of 25% (v/v) each, which were mixed properly prior to plating. The media was poured in Petri dishes at 20 ml per plate. Plates were inoculated separately with 5-mm mycelia plugs of the pathogen S. sclerotiorum placed in the centres of the plates. The inoculated plates were incubated at 25˚C ± 1˚C for 5 days. Percent mycelial growth inhibition of each pathogen was calculated using the formula:
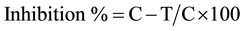
where C = control (radial growth of the pathogen) and T = treatment (radial growth of the pathogen after inhibition by the antagonist).
2.7. Plant Cultivation under Greenhouse Conditions
Both genotypes B. napus and Raphanus alboglabra were planted in 25-cm-diamter pots containinga 3-kg mixture of sand and peat moss at a ratio of 1:1 that had been autoclaved twice for 30 min within a 24-h interval. Five seeds of each genotype were sown into each pot. Both genotypes were grown under greenhouse conditions at 18/14 (±1) ˚C (day/night) temperature and a light intensity of 150 µE/m2/s1 for 12-h light/dark cycles for 35 to 40 days. Irrigation was applied by drenching twice a week.
2.8. Screening of Fungal Strains Based on ISR-Eliciting Potential and Effects on Plant Growth
To screen the fungal biotic elicitors capable of eliciting ISR, 15-day-old both B. napus and Raphanus albogla- bra plants were treated with water (control) or inoculated with suspensions 100 ml (1.5 × 107 CFU/ml) of T. harzianum TH12 and (1 × 107 CFU/ml) T. viride TV10 as well as their CF and CFH by spraying the leaves and stems, as opposed to soil drenching. The S. sclerotiorum tested was grown in PDB media for 4 weeks incubated with a rotary shaker (85 rpm) at 22˚C ± 2˚C. The pathogen propagules of each flask containing 30 ml media was blended and then mixed with the upper soil surface of each pot 1 day after application of the last biotic elicitor. Each treatment consisted of three replicates. Disease incidence and intensity were recorded at 15 days after treatment using a rating from 0 (no stem discolouration) to 5 (discolouration, sclerotia, premature ripening of the whole plant, and low seed production). Numerical values of 0, 1.25, 2.5, 3.75, and 5, respectively, were assigned to the 1 to 5 rating, and the disease index (DI) was calculated following the formula of Dueck et al. [29] :
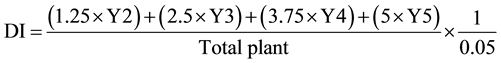
where DI = disease index and Y2, Y3, Y4, and Y5 = numbers of plants with scores of 2 to 5, respectively.
Based on the percent disease incidence and intensity, the mean degree of infection was calculated following the formula of Krüger [30] :
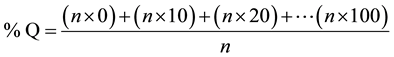
where Q = degree of infection and n = numbers of assessed plants whose percentages were 0 to 100.
Fresh and dry (70˚C for 48 h) weights of shoots and roots as well as plant height were determined. Five plants were chosen from each pot for these determinations at 45 days after sowing.
2.9. Statistical Analysis
Antagonistic effects of biotic elicitor isolates and their cell-free CF and CFH against S. sclerotiorum and all greenhouse experiments were analysed as completely randomised designs with three replications using GenStat software, and means were compared using least-significant difference tests [31] .
3. Results and Discussion
3.1. Morphological Identification of Trichoderma spp.
Thirteen isolates of T. harzianum and 17 of T. viride were isolated from rhizospheres of healthy rapeseed plants. Morphological characteristics have been used to characterise and discriminate Trichoderma species since 1969 [32] . Trichoderma isolated in PDA culture media after 7 days of incubation at 28˚C ± 2˚C showed differences in mycelial culture growth and patterns of sporulation (Figure 1). The characters of mycelia colour patterns varied noticeably between the Trichoderma isolates, from colourless to yellow and white; their shapes consisted of concentric rings, and hyphae were septated, smooth-walled, and hyaline. The conidia were green to yellow- green or dark green colour and their shapes were rough and subglobose. Conidia production was more intense in the centre but declined toward the margins. Morphological studies can provide information to identify Trichoderma species, but it is insufficient because Trichoderma species have relatively few morphological characters and limited disparity, which may cause interference and misidentification [33] . Furthermore, cultural conditions also can affect morphological characteristics [34] .
3.2. Molecular Characterisation of TH12 and TV10 Isolates
The Trichoderma genus is important from a practical perspective due to the ability of strains, belonging to various species, to inhibit and reduce effects of fungal plant pathogens. To use certain strains for plant protection as useful bio products, it is necessary to identify and characterise the species. The effects of environmental conditions on physiological and morphological characteristics have made accurate identification extremely difficult [35] . For better identification, molecular techniques are very useful. The ITS regions of rDNA of T. harzianum TH12 and T. viride TV10 were amplified using primers ITS1 and ITS4. ITS 5.8SDNA fragments approximately 625base pairs in length (according to estimates via agarose gel electrophoresis) were successfully amplified from T. harzianum TH12 and T. viride TV10 (Figure 2). Amplified PCR products of 5.8S rDNA genes from T. harzianum TH12 and T. viride TV10 isolates were sequenced; BLAST on National Centre for Biotechnology Information (NCBI) was then used to confirm the species identities of T. harzianum TH12 and T. viride TV10, which were morphologically identified earlier.
3.3. Detection Ability of Antagonists of Trichoderma Isolates in Vitro
The isolates were tested for their ability to reduce mycelia growth of S. sclerotiorum fungi using dual-culture
 (a) (b)
(a) (b)
Figure 1. Two different isolates of (a) T. harzianum TH12 and (b) T. viride TV10 on potato dextrose agar after 7 days of incubation.

Figure 2. Polymerase chain reaction (PCR) amplification products obtained from total DNA extracted from Trichoderma spp. using internal transcribed spacer (ITS) 1 and ITS4 primers. Lane 1 = T. viride TV10. Lane 2 = T. harzianum TH12. M = DNA marker.
techniques. The results revealed that the degree of inhibition varied greatly, ranging from 1.67% to 100% (Table 1), which facilitated of the biotic elecitors. On the basis of percent inhibition of the radial growth of the test pathogens, two specific isolates, T. harzianum TH12 and T. viride TV10, exhibited higher antagonistic potential were selected. These isolates (TH12 and TV10) reduced the growth of the pathogen S. sclerotiorum, with inhibition zones reaching 100 and 84.44%, respectively. Their cell-free CF caused the inhibition zones to reach 100 and 61.48%, respectively, and the inhibition zones of their CFH reached 50.00% and 35.18%, respectively (Figure 3). These results indicate that among the different strains of the same fungal genus isolated from the same ecological niche, a significant difference exists in terms of their antagonistic behaviour against a pathogen. Tricoderma is a genus of saprophytic fungi. These Trichoderma fungi are present in various soils and rhizo- sphere microflora, and they are well known as biological control agents against various plant pathogens. It should be noted that percent inhibition of radial S. sclerotiorum growth by different isolates of fungi varied widely in the present study (P < 0.05). These isolates overgrew the pathogen and completely suppressed it within 7 days of inoculation. These results indicate that T. harzianum and T. viride strongly can prevent mycelial growth of S. sclerotiorum in plates [36] -[38] . Generally, considerable variation in the inhibitory properties of resistant biotic elicitor isolates exists and is discernible. This could be due to differences in the quantity and quality of the material produced. Some strains inhibit the growth of fungal pathogens through production of volatile and non-volatile antibiotics and production of low-molecular-weight diffusible compounds or antibiotics [39] .
Trichoderma isolates are capable of producing many powerful plant-degrading enzymes and more than 200 types of antibiotics that are highly toxic to any macro- and microorganism [40] . Different mechanisms have been proposed for their mycoparasitic secretion of lytic enzymes, such as cellulase (β-1, 4-glucanase), proteases, and chitinases, which degrade cell walls [41] . Production of the antibiotic is likely located in the tips of growing hyphae, with components responsible for the antagonism secreted at the points of contact with the pathogen [42] . T. harzianum and T. viride CF to inhibit growth of Fusarium moniliforme pathogens might be due to the production of volatile compounds and liberation of extracellular enzymes, such as those with amylolytic, pectinolytic, proteolytic, and cellulolytic activities [43] .
3.4. ISR of Biotic Elicitors to S. sclerotiorum in B. napus and Raphanus alboglabra
The results showed that the pathogenic fungus S. sclerotiorum effect is significant in B. napus and Raphanus alboglabra plants compared to non-infected plants with the fungus pathogen (Figure 4). The results in Table 2 show that B. napus and Raphanus alboglabra plants grown in soils amended with TH12, TV10, CF, and CFH resulted in reductions of disease symptoms compared to control plants. The treatments were sprayed on rapeseed leaves and stems 1 day prior to inoculation with S. sclerotiorum in the greenhouse. The disease was evaluated by determining rates of disease intensity in plants after 15 days of challenge by examining the growth of the pathogen. Treatment with the biotic elicitors CF and CFH significantly ISR in treatment plants compared to control

Table 1. In-vitro screening of Trichoderma isolates against S. sclerotiorum.
 S. sclerotiorum (Control)
S. sclerotiorum (Control) TH12 + S. sclerotiorum Cell-free CF TH12 + S. sclerotiorum Cell-free CF with heat TH12 + S. sclerotiorum
TH12 + S. sclerotiorum Cell-free CF TH12 + S. sclerotiorum Cell-free CF with heat TH12 + S. sclerotiorum TV10 + S. sclerotiorum Cell-free CF TV10 + S. sclerotiorum Cell-free CF with heat TV10 + S. sclerotiorum
TV10 + S. sclerotiorum Cell-free CF TV10 + S. sclerotiorum Cell-free CF with heat TV10 + S. sclerotiorum
Figure 3. Antagonistic activities of T. harzianum TH12 and T. viride TV10 as well as their cell-free culture filtrates (CF) and culture filtrates treated with heat (CFH) against S. sclerotiorum. LSD = least significant difference.

Figure 4. Effects S. sclerotiorum on B. napus (AACC) and Raphanus oleracea (RRCC). (a) = non-pathogen (AACC). (b) = AACC infected. (c) = non-pathogen (RRCC). (d) = RRCC infected.
Table 2. Effect of T. harzianum TH12 and T. viride TV10 and their CF and CFH on B. napus and Raphanus alboglabra infected by S. sclerotiorum in greenhouse conditions.
Tables with the some treatments are not significantly different (P = 0.05). *CF = culture filter of resistant biotic elecitor. *CFH = culture filter of resistant biotic elecitor treated with heat. LSD = least significant difference.
plants (Table 2). There was no significant difference in inhibitory efficacy between the cell suspensions for the biotic elicitors CF and CFH. Suppression of SSR on plant leaves by cell suspensions was observed when biotic elicitor cells were applied to plant leaves before plants were inoculated with S. sclerotiorum. This indicates antifungal substances produced by biotic elicitor cells can penetrate well inside plant tissues to exhibit therapeutic effects and exert preventive effects against SSR disease. Therefore, once S. sclerotiorum infects plant tissue, it may be affected not at all or only slightly by antifungal substances produced in vitro by biotic elicitors [44] [45] .
Biotic elicitors in plant roots play a role in improving plant health. ISR and plant growth promotion are the important mechanisms by which selected mycorrhizal fungi and plant growth-promoting rhizobacteria in the rhizosphere prime the entire plant to promote defence against a broad range of pathogens [46] . A wide range of organisms associated root of plant, such as Trichoderma spp., prime plant immune systems for enhanced defence, indirectly activating the defensive system [22] [23] . Atagonistic activities of resistant biotic elicitors and their CF have been reported against several fungal plant pathogens. The present results show that the effects of the pathogenic fungus S. sclerotiorum are significant in rapeseed compared to untreated plants. The degree of infection and disease index in treatments were 81.40% and 83.50%, respectively, in genotype B. napus, and both were 0.00 in genotype and Raphanus alboglabra. The present results also indicate that the pathogenic fungus causes reduced root and shoot fresh and dry weights and reduced shoot height (Table 2).
The absence of resistance source in B. napus, AACC has dramatically restrained resistance breeding against phytopathogen. Newly, many of wild Brassica oleracea (B. incana) accessions (CC) identified with high level of resistance against S. sclerotiorum pathogen which causes severe crop losses in oilseed production all around the world [47] [48] .
A high level of resistance to S. sclerotium was reported in parental species of rapeseed such as B. oleracea cytodeme and was evaluated for resistance over 2 years. Relative to RS lines exhibited stronger resistance against S. sclerotiorum than “Zhongyou 821”, a cultivar of B. napus has partial resistant; RS lines are one of the most important sources of resistance in current rapeseed against Sclerotinia stem rot [49] . Significant differences were showed across the two growing seasons of B. incana and B. oleracea var. alboglabra between parents and the progeny for measures of both stem and leaf sclerotina resistance [50] .
The biotic elicitors significantly reduced (P ≤ 0.5) the SSR disease incidence in the rapeseed infected with S. sclerotiorum. The highest reduction in SSR degree of infection and DI were 7.22% and 6.67%, respectively, treated with TH12. The degree of infection and DI of the CF reached 20.00% and 16.67%, respectively (Figure 4), and the CFH treatment reached 35.00% and 21.67%, respectively, in B. napus. On the other hand, the degree of infection and DI reached 17.78% to 11.67% of that treated with TV10. Its CF reached 33.33% and 23.33% and CFH reached 37.78% and 28.33%, respectively, in B. napus. In and Raphanus alboglabra, values of 0.00% for both in all treatments were observed.
These results were consistent with those of Yoshioka et al. [51] , who described the ability of a cell-free CF of Trichoderma asperellum STK-1 to induce systemic resistance in A. thaliana, decrease lesion development and growth of the pathogen P. syringae pv. Tomato DC3000. Trichoderma harzianum T39, when applied before 48 to 72 h of inoculation, reduced downy mildew disease approximately 63% on susceptible grapevine cultivars caused by Plasmopara viticolain greenhouse conditions [52] . The PGPF Penicillium spp. GP16-2 and its cell- free CF were used to stimulate systemic resistance in A. thaliana against the bacterial leaf speck pathogen P. syringae [13] . They act against plant pathogens in several ways, either by lytic enzymes, antibiotic-mediated suppression, mycoparasitism and other by product production, induction of host resistance, or competition for nutrients [9] . SA accumulation and SA signalling have been demonstrated to facilitate resistance to the pathogens Colletotrichum higginsianum and P. syringae pv. maculicola in B. rapa and A. thaliana after treatment with plant activator HM [15] . Tested isolates of Trichoderma spp. can produce organic acids, such as gluconic and fumaric acids that permit the solubilisation of phosphates and decrease soil pH, micronutrients, and mineral cations, such as Mg, Mn, and Fe which are useful for plant metabolism [53] [54] . Jian et al. [55] also demonstrated that the same biogenic factors can provide protection in pea plants by causing increases in H2O2 (hydrogen peroxide) production, ascorbic acid, antioxidant enzymes, and lipid peroxidation against the pathogen S. sclerotiorum. The resistant biotic elicitor modes of action were not measured experimentally in this study. Amendment with biotic elicitors increased root and shoot fresh and dry weights and height of rapeseed plants and differed significantly compared to control plants infected by the pathogen (Table 2). The TH12 treatment ranked as most effective on B. napus with 2.390, 0.065, 5.745, and 0.880 g (root fresh weight, root dry weight, shoot fresh weight, and shoot dry weight, respectively) and 20.33 cm (height). This was followed by TV10, which reached 2.350, 0.058, 5.614, and 0.755 g and 21.11 cm, respectively, followed by CFH of TV10, which reached 2.324, 0.050, 4.437, and 0.677 g and 20.33 cm, respectively. The CFH of TV10 provided the lowest protection against the disease (2.226, 0.038, 4.228, and 0.539 g and 18.20 cm, respectively). The TH12 treatment ranked as most effective on Raphanus alboglabra, reaching3.00, 0.093, 5.95, and 1.30 g and 21.33 cm, respectively.
This was followed by TV10, which reached 2.98, 0.090, 5.90, and 1.28 g and 21.32 cm, respectively. The TH12 CF treatment reached 2.88, 0.090, 5.88, and 1.18 g and 20.67 cm, respectively, and the TH12 CFH treatment reached 2.75, 0.078, 5.59, and 1.25 g and 18.67 cm, respectively. Autoclaved T. harzianum T12 spores were not significantly different from living T12 spores at reducing uredial pustule percentage and reducing activity of bean rust disease caused by Uromyces appendiculatus, but autoclaving T. harzianum spores eliminated their ability to reduce rust spore infections [23] . Trichoderma species have the ability to suppress pathogens belonging to many other genera. This ability is due to the production of non-volatile and volatile organic compounds, extracellular lytic enzymes, high proliferation rates, and high competitive saprophytic abilities [56] . Their abilities to protect plantscan be attributed to the upregulation of a variety pathogenesis-related proteins and defence-related proteins (e.g., peroxidases, chitinases, glucanases, and specific phytoalexins), and enzymatic activities usually increase during a pathogenic attack [17] [57] . Protection of plants by Trichoderma spp. using ISR is considered of fundamental importance today compared with the “classical” direct, antagonistic, and competition methods that occur with a pathogen in host plants [2] [53] .
Finally, some plants do not have adequate protection to most potential pathogens that continually attack plants, such as fungi, bacteria, and viruses. This feature is referred to as “non-host resistance”, such as B. napus in this study. This genotype of plants needs biotic or abiotic elicitors to induce resistance against various phytopathogens. The T. harzianum and T. viride and their CF and CFH can be used as elicitors of resistance in B. napus. Moreover, some genotypes have the ability to resist various phytopathogens. This feature is referred to as “host resistance”, such as genotype Raphanus alboglabra; this study is the first to reveal the ability of Raphanus alboglabra to demonstrate resistance to SSR disease caused by S. sclerotiorum.
4. Conclusion
The application of useful resistant biotic elicitors for protection and increased rapeseed crop production is now practised in agriculture. The resistant biotic elicitors may better adapt to environmental changes, protect against a broader range of pathogens of plant varieties, increase the genetic diversity of biocontrol systems that continue longer in the rhizosphere without negatively impacting the environment, and utilise a wider array of ISR mechanisms.
References
- Dixon, G.R. (2007) Vegetable Brassicas and Related Crucifers. CAB International, Wallingford.
- Harman, G.E., Howell, C.R., Viterbo, A., Chet, I. and Lorito, M. (2004) Trichoderma Species-Opportunistic, Avirulent Plant Symbionts. Nature Reviews Microbiology, 2, 43-56. http://dx.doi.org/10.1038/nrmicro797
- Leflon, M., Eber, F., Letanneur, J.C., Chelysheva, V., Coriton, O., Huteau, V., Ryder, V., Barker, G., Jenczewski, E. and Chèvre, A.M. (2006) Pairing and Recombination at Meiosis of Brassica rapa (AA) Brassica napus (AACC) Hybrids. Theoretical and Applied Genetics, 113, 1467-1480. http://dx.doi.org/10.1007/s00122-006-0393-0
- Li, X., Guo, W., Wang, B., Li, X., Chen, H., Wei, L., Wang, Y., Wu, J. and Lon, H. (2010) Instability of Chromosome Number and DNA Methylation Variation Induced by Hybridization and Amphidiploid Formation between Raphanus sativus L. and Brassica alboglabra Bailey. BMC Plant Biology, 10, 207. http://dx.doi.org/10.1186/1471-2229-10-207
- Long, H., Chen, C., Wang, B. and Feng, Y. (2015) rDNA Genetic Imbalance and Nucleolar Chromatin Restructuring Is Induced by Distant Hybridization between Raphanus sativus and Brassica alboglabra. PLoS ONE, 10, e0117198. http://dx.doi.org/10.1371/journal.pone.0117198
- Dalili, A. (2013) Greenhouse and Field Evaluation of Rapeseed Cultivars and Lines for Resistance against Sclerotinia sclerotiorum (Lib.) de Bary. Journal of Oilseed Brassica, 4, 61-65.
- Mohammad, A. and Dalil, A. (2013) Evaluation of Rapeseed Genotypes Resistance to Sclerotinia sclerotiorum (Lib.) De Bary. American-Eurasian Journal of Agricultural Environmental Sciences, 13, 517-520.
- del Río, L.E., Bradlehl, C.A., Henson, R.A., Endres, G.J., Hanson, B.K., Mckay, K., Halvorson, M., Porter, P.M., Le Gare, D.G.H. and Lamey, A. (2007) Impact of Sclerotinia Stem Rot on Yield of Canola. Plant Disease, 91, 191-194. http://dx.doi.org/10.1094/PDIS-91-2-0191
- Pal, K.K and Gardener, M. (2006) Biological Control of Plant Pathogens. The Plant Health Instructor.
- Walters, D.R., Ratsep, J. and Havis, N.D. (2013) Controlling Crop Diseases Using Induced Resistance: Challenges for the Future. Journal of Experimental Botany, 64, 1263-1280. http://dx.doi.org/10.1093/jxb/ert026
- Spoel, S.H., Koornneef, A., Claessens, S.M., Korzelius, J.P., Van Pelt, J.A., Mueller, M.J., Buchala, A.J., Métraux, J., Brown, R., Kazan, K., Van Loon, L.C., Dong, X. and Pieterse, C.M.J. (2003) NPR1 Modulates Cross-Talk between Salicylate- and Jasmonate-Dependent Defense Pathways through a Novel Function in the Cytosol. The Plant Cell, 15, 760-770. http://dx.doi.org/10.1105/tpc.009159
- Verhagen, B.W., Glazebrook, J., Zhu, T., Chang, H.S., van Loon, L.C. and Pieterse, C.M. (2004) The Transcriptome of Rhizobacteria-Induced Systemic Resistance in Arabidopsis. Molecular Plant-Microbe Interactions Journal, 17, 895- 908. http://dx.doi.org/10.1094/MPMI.2004.17.8.895
- Hossain, Md.M., Sultana, F., Kubota, M. and Hyakumachi, M. (2008) Differential Inducible Defense Mechanisms against Bacterial Speck Pathogen in Arabidopsis thaliana by Plant-Growth Promoting-Fungus Penicillium sp. GP16-2 and Its Cell Free Filtrate. Plant and Soil, 304, 227-239. http://dx.doi.org/10.1007/s11104-008-9542-3
- Maksimov, I.V., Abizgil’dina, R.R. and Pusenkova, L.I. (2011) Plant Growth Promoting Rhizobacteria as Alternative to Chemical Crop Protectors from Pathogens (Review). Applied Biochemistry and Microbiology, 47, 373-385. http://dx.doi.org/10.1134/s0003683811040090
- Narusaka, M., Minami, T., Iwabuchi, C., Hamasaki, T., Takasaki, S., Kawamura, K. and Narusaka, Y. (2015) Yeast Cell Wall Extract Induces Disease Resistance against Bacterial and Fungal Pathogens in Arabidopsis thaliana and Brassica Crop. PLoS ONE, 10, e0115864. http://dx.doi.org/10.1371/journal.pone.0115864
- Schuster, A. and Schmoll, M. (2010) Biology and Biotechnology of Trichoderma. Applied Microbiology and Biotechnology, 87, 787-799. http://dx.doi.org/10.1007/s00253-010-2632-1
- Yedidia, I., Benhamou, N. and Chet, I. (1999) Induction of Defense Responses in Cucumber Plants (Cucumis sativus L.) by the Biocontrol Agent Trichoderma harzianum. Applied and Environmental Microbiology, 65, 1061-1070.
- Sharma, P. and Sain, S.K. (2004) Induction of Systemic Resistance in Tomato and Cauliflower by Trichoderma spp. against Stalk rot Pathogen, Sclerotinia sclerotiorum Lib de Bary. Journal of Biological Control, 18, 21-27.
- Jogaiah, S., Abdelrahman, M., Tran, L.P. and Shin-ichi, I. (2013) Characterization of Rhizosphere Fungi That Mediate Resistance in Tomato against Bacterial Wilt Disease. Journal of Experimental Botany, 64, 3829-3842. http://dx.doi.org/10.1093/jxb/ert212
- Elsharkawy, M.M., Shimizu, M., Takahashi, H., Ozaki, K. and Hyakumachi, M. (2013) Induction of Systemic Resistance against Cucumber Mosaic Virus in Arabidopsis thaliana by Trichoderma asperellum SKT-1. The Plant Pathology Journal, 29, 193-200. http://dx.doi.org/10.5423/PPJ.SI.07.2012.0117
- Umamaheswari, R., Sivakumar, M., Subramanian, S. and Samiyappan, R. (2004) Induction of Systemic Resistance by Trichoderma viride Treatment in Greengram (Vigna radiata) against Root-Knot Nematode Meloidogyne incognita. Current Nematology, 15, 1-7.
- Prasad, S.B., Kamala, G. and Ganesh, S.P. (2013) Efficacy of Trichoderma viride to Induce Disease Resistance and Antioxidant Responses in Legume Vigna Mungo Infested by Fusarium oxysporum and Alternaria alternate. International Journal of Agricultural Science and Research (IJASR), 3, 285-294.
- Burmeister, L. and Hau, B. (2009) Control of the Bean Rust Fungus Uromyces appendiculatus by Means of T. harzianum: Leaf Disc Assays on the Antibiotic Effect of Spore Suspensions and Culture Filtrates. BioControl, 54, 575-585. http://dx.doi.org/10.1007/s10526-008-9202-9
- Elad, Y. and Chet, I. (1983) Improved Selective Media for Isolation of Trichoderma spp. or Fusarium spp. Phytoparasitica, 11, 55-58. http://dx.doi.org/10.1007/BF02980712
- Askew, D.J. and Laing, M.D. (1993) An Adapted Selective Medium for the Quantitative Isolation of Trichoderma Species. Plant Pathology, 42, 686-690. http://dx.doi.org/10.1111/j.1365-3059.1993.tb01553.x
- Harris, J.L. (2000) Safe, Low-Distortion Tape Touch Method for Fungal Slide Mounts. Journal of Clinical Microbiology, 38, 4683-4684.
- Chakraborty, B.N., Chakraborty, U., Saha, A. and Dey, P.L. (2010) Molecular Characterization of Trichoderma viride and Trichoderma harzianum Isolated from Soils of North Bengal Based on rDNA Markers and Analysis of Their PCR-RAPD Profiles. Global Journal of Biotechnology & Biochemistry, 5, 55-61.
- White, T.J.T., Lee, B.S. and Taylor, J.W. (1990) Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: Innis, M.A., Gelfand, D.H., Sninsky, J.J. and White, T.J., Eds., PCR Protocols: A Guide to Methods and Applications, Academic Press, New York, 315-322. http://dx.doi.org/10.1016/b978-0-12-372180-8.50042-1
- Dueck, J., Morrall, R.A.A. and McKenzie, D.L. (1983) Control of Sclerotinia sclerotiorum in Rapeseed with Fungicides. Canadian Journal of Plant Pathology, 5, 289-293. http://dx.doi.org/10.1080/07060668309501613
- Krüger, W. (1983) Oilseed Rape, Pests and Diseases. Semundo, Hamburg, 119 p.
- Gomez, K.A. and Gomez, A.A. (1984) Statistical Procedures for Agricultural Research. 2nd Edition, John Wiley and Sons, Inc., London, 13-175.
- Gams, W. and Bissett, J. (2002) Morphology and Identification of Trichoderma. In: Kubicek, C.P. and Harman, G.E., Eds., Trichoderma and Gliocladium: Basic Biology, Taxonomy and Genetics, Taylor & Francis Ltd., London, 3-31.
- Anees, M., Tronsmo, A., Edel-Hermann, V., Hjeljord, L.G., Héraud, C. and Steinberg, C. (2010) Characterization of Field Isolates of Trichoderma Antagonistic against Rhizoctonia solani. Fungal Biology, 114, 691-701.
- Diguta, C.F., Vincent, B., Guilloux-Benatier, M., Alexandre, H. and Rousseaux, S. (2011) PCR ITS-RFLP: A Useful Method for Identifying Filamentous Fungi Isolates on Grapes. Food Microbiology, 28, 1145-1154. http://dx.doi.org/10.1016/j.fm.2011.03.006
- Siddiquee, S., Guan, F.A. and Aziz, E.R. (2007) Phylogenetic Relationship of Trichoderma harzianum Based on the Sequence Analysis of the Internal Transcribed Spacer Region-1 of the rDNA. Journal of Applied Sciences Research, 3, 896-903.
- Ojaghian, M.R. (2011) Potential of Trichoderma spp. and Talaromyces flavus for Biological Control of Potato Stem Rot Caused by S. sclerotiorum. Phytoparasitica, 39, 185-193. http://dx.doi.org/10.1007/s12600-011-0153-9
- Saxena, D., Tewari, A.K. and Rai, D. (2014) In Vitro Antagonistic Assessment of T. harzianum PBT 23 against Plant Pathogenic Fungi. Journal of Microbiology and Biotechnology Research, 4, 59-65.
- Troian, R.F., Steindorff, A.S., Ramada, M.H.S., Arruda, W. and Ulhoa, C.J. (2014) Mycoparasitism Studies of Trichoderma harzianum against Sclerotinia sclerotiorum: Evaluation of Antagonism and Expression of Cell Wall-Degrading Enzymes Genes. Biotechnology Letters, 36, 2095-2101. http://dx.doi.org/10.1007/s10529-014-1583-5
- Rini, C.R. and Sulochana, K.K. (2007) Usefulness of Trichoderma and Pseudomonas against Rhizoctonia solani and Fusarium oxysporum Infecting Tomato. Journal of Tropical Agriculture, 45, 21-28.
- Kubicek, C.P. and Harman, G.E. (1998) Trichoderma and Gliocladium. Vol. 1, Basic Biology, Taxonomy and Genetics. Taylor & Francis, London.
- Witkowsha, D. and Maj, A. (2002) Production of Lytic Enzymes by Trichoderma spp. and Their Effect on the Growth of Phytopathogenic Fungi. Folia Microbiologica, 47, 279-282. http://dx.doi.org/10.1007/BF02817652
- Michalikova, A. and Michrina, J. (1997) Regulatory Potential of T. harzianum B1 in Relation with Phytopathogenous Microorganisms. Proc. XIVth Slovak and Czech Plant Protect, Conference, Nitra, 291-292.
- Calistru, C., McLean, M. and Berjak, P. (1997) In Vitro Studies on the Potential for Biological Control of Aspergillus flavus and Fusarium moniliforme by Trichoderma Species: A Study of the Production of Extracellular Metabolites by Trichoderma Species. Mycopathologia, 137, 115-124. http://dx.doi.org/10.1023/A:1006802423729
- Ismail, E., Turkkan, M., Karaca, G.H. and Kılıc, G. (2011) Evaluation of in Vitro Antifungal Activity of Potassium Bicarbonate on Rhizoctonia solani AG 4 HG-I, Sclerotinia sclerotiorum and Trichoderma sp. African Journal of Biotechnology, 10, 8605-8612.
- Yang, R., Han, Y.C., Li, G.Q., Jiang, D.H. and Huang, H.C. (2007) Suppression of S. sclerotiorum by Antifungal Substances Produced by the Mycoparasite Coniothyrium minitans. European Journal of Plant Pathology, 119, 411-420. http://dx.doi.org/10.1007/s10658-007-9174-0
- Van Wees, S.C.M., Ent, S.V. and Pieterse, C.M.J. (2008) Plant Immune Responses Triggered by Beneficial Microbes. Current Opinion in Plant Biology, 11, 443-448. http://dx.doi.org/10.1016/j.pbi.2008.05.005
- Mei, J., Ding, Y., Lu, K., Wei, D., Liu, Y., Disi, J.O., Li, J., Liu, L., Liu, S., McKay, J. and Qian, W. (2013) Identification of Genomic Regions Involved in Resistance against Sclerotinia sclerotiorum from Wild Brassica oleracea. Theoretical and Applied Genetics, 126, 549-556. http://dx.doi.org/10.1007/s00122-012-2000-x
- Mei, J., Qian, L., Disi, J.O., Yang, X., Li, Q., Li, J., Frauen, M., Cai, D. and Qian, W. (2011) Identification of Resistant Sources against Sclerotinia sclerotiorum in Brassica Crops with Emphasis on B. oleracea. Euphytica, 177, 393-400. http://dx.doi.org/10.1007/s10681-010-0274-0
- Ding, Y., Mei, J., Li, Q., Liu, Y., Wan, H., Wang, L., Becker, H.C. and Qian, W. (2013) Improvement of Sclerotinia sclerotiorum Resistance in Brassica napus by Using B. oleracea. Genetic Resources and Crop Evolution, 60, 1615- 1619. http://dx.doi.org/10.1007/s10722-013-9978-z
- Disi, J., Mei, J., Wei, D., Ding, Y. and Qian, W. (2014) Inheritance of Leaf and Stem Resistance to Sclerotinia sclerotiorum in a Cross between Brassica incana and Brassica oleracea var. alboglabra. The Journal of Agricultural Science, 152, 146-152. http://dx.doi.org/10.1017/S0021859613000087
- Yoshioka, Y., Ichikawa, H., Naznin, H.R., Kogure, A. and Hyakumachi, M. (2012) Systemic Resistance Induced in Arabidopsis thaliana by Trichoderma asperellum SKT-1, a Microbial Pesticide of Seed Borne Disease of Rice. Pest Management Science, 68, 60-66. http://dx.doi.org/10.1002/ps.2220
- Perazzolli, M., Dagostin, S., Ferrari, A., Elad, Y. and Ilaria, P. (2008) Induction of Systemic Resistance against Plasmopara viticola in Grapevines by T. harzianum T39 and Benzothiadiazole. Biological Control, 47, 228-234. http://dx.doi.org/10.1016/j.biocontrol.2008.08.008
- Benitez, T., Rincon, A.M. and Condon, A.C. (2004) Biocontrol Mechanism of Trichoderma Strains. International Microbiology, 7, 249-260.
- Harman, G.E., Howell, C.R., Viterbo, A., Chet. I. and Lorito, M. (2004) Trichoderma Species―Opportunistic, Avirulent Plant Symbionts. Nature Reviews Microbiology, 2, 43-56. http://dx.doi.org/10.1038/nrmicro797
- Jain, A., Singh, A., Singh, S. and Singh, H.B. (2012) Microbial Consortium-Induced Changes in Oxidative Stress Markers in Pea Plants Challenged with Sclerotinia sclerotiorum. Journal of Plant Growth Regulation, 32, 388-398. http://dx.doi.org/10.1007/s00344-012-9307-3
- Elad, Y., David, D.R., Levi, T., Kapat, A. and Kirshner, B. (1999) In: Lyr, H., Russell, P.E., Dehne, H.W. and Sisler, H.D., Eds., Modern Fungicides and Antifungal Compounds II, Intercept Ltd., Andover, 459-467.
- Yedidia, I., Benhamou, N., Kapulnik, Y. and Chet, I. (2000) Induction and Accumulation of PR Proteins Activity during Early Stages of Root Colonization by the Mycoparasite Trichoderma harzianum Strain T-203. Plant Physiology and Biochemistry, 38, 863-873. http://dx.doi.org/10.1016/S0981-9428(00)01198-0
NOTES

*Corresponding authors.