American Journal of Plant Sciences
Vol.4 No.7A1(2013), Article ID:34785,3 pages DOI:10.4236/ajps.2013.47A1004
Kuschakewiczia versus Solenanthus—Palynological Data and the Generic Position of Kuschakewiczia turkestanica Regel and Smirnov (Boraginaceae)
![]()
Department of Botany, Moravian Museum, Brno, Czech Republic.
Email: ksutory@mzm.cz
Copyright © 2013 Karel Sutorý. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received May 15th, 2013; revised June 17th, 2013; accepted July 3rd, 2013
Keywords: Central Asia; Kuschakewiczia; Solenanthus
ABSTRACT
The genus Kuschakewiczia, Regel and Smirnov [1], was described including only one species, K. turkestanica. Later it was joint with the genus Solenanthus, although there are characters (corolla having narrow long lobes, filaments inserted on the corolla margin, low gynobasis, small scar on nutlets after disconnecting without any strip of tissue, reduced number of nutlets in the ripe fruit, etc.) which distinguish the two genera sufficiently. Palynological data, although not significantly different, support its separate positions on the discrete generic level.
1. Introduction
The genus Kuschakewiczia was described by Regel and Smirnov [1] including only one species, K. turkestanica, from the neighborhoods of Tashkent in Tajikistan. Differential characters of the new genus should be the position of the stamen insertion (stamina fauce inserta), the structure of the nutlet surface (facie antice areola concava saepe marginata) and the number of nutlets in imamture flowers (maturae semper solitariae). Later Lipsky [2] considered these characters to be insufficient and classified this species into the genus Solenanthus under the name S. kuschakewiczii. He did not find differences between both genera except for the position of the stamens. The name of the species was brought in accordance with nomenclatural rules by Kuznetzow [3] and three years later illegimately also by Macbride [4]. Meling [5] found five characters distinguishing the genus Kuschakewiczia from the genus Solenanthus: 1) corolla having narrow long lobes, 2) filaments inserted on the corolla margin, 3) low gynobasis, 4) small scar on nutlets after perfect disconnecting (no rest of tissue strip), 5) inconspicuous thickness of the cells in the endocarp walls. Further the reduced number of nutlets in ripe fruits, the structure of the fruit surface and other characters are to be added. Palynological data had not yet been used to solve the problem of the generic level of the genus Kuschakewiczia. Similarly as in other cases in Boraginaceae they can be useful. Already Avetisyan [6] started to use palynological data for this purpose and later also many others, such as Clarke et al. [7], Sahay [8], Díaz & Valdés [9], Popova et al. [10], Khatamzas [11], Liu et al. [12], and lately Bigazzi et al. [13]. For differentiation of newly created genera they were used by Barbier et Mathez [14] and Sutorý [15].
2. Materials and Methods
Pollen material of four specimens collected in south Tajikistan and preserved at the V. L. Komarov Botanical Institute of the Russian Academy of Sciences in St. Petersburg was used (Michelson A. I., No. 1245, 4.5.1913, Liičevskij I.A. No. 46, 30.4.1938, Varivceva E. A., Nepli G. N., No. 269, 4.5. 1948, Filatov E. M., Saprunova R. M., No. 30, 27.3.1964). Pollen grains were measured on the photographs of pollen grains. SEM microphotographs were produced using the Tescan Mira 3 LMU electron microscope.
3. Results and Discussion
Only three previously published sources can be partly utilized for the problem under discussion. These are consistent in the shape and structure of the pollen grains but different in their dimensions. Avetisyan [6] mentions measurements of 10.5 × 9.8 μm, Barbier et Mathez [14] 9 × 8.2 μm, and Bigazzi et al. [13] 6.4 × 7.4 µm in size. The discrepancies are probably due to different methods they used. My preliminary measurements (15 pollen grains) carried out on material from the four different samples come close to the results of Bigazzi et al. [13], namely with an average of 6.9 × 7.6 µm (the amplitude being 6.2 - 7.2 × 7.1 - 8.0 µm). In any case, all sources agree that the pollen grains are small, on the lower end of the range of other species in Solenanthus (7.5 × 6.3 µm to 11.0 × 9.9 µm). They have a very low ratio (1.2) of polar axis: equatorial diameter, thus they are almost spherical in shape, which is not different from most other species of Solenanthus. The ratio in my measurements was 1.1. The pollen grains are tricolporate, i.e. having three compound apertures (colpori) altering with three simple apertures (pseudoapertures, colpi) and in the equatorial region they are connected with a distinct belt (ectocingulus). Bigazzi et al. [13] call this type of pollen the “Cynoglossum officinale type” and they describe the pollen in Solenanthus as “essentially uniform” and the apertures in Solenanthus with the following words: “The ectoapertures [composed apertures, colpori] are rhombic in outline with lalongate [= transversely elongate] endoapertures (1.2 × 3.2 μm), situated at about the equator, the colporate apertures are equal to or shorter than the fusiform simple colpi [pseudocolpi]”. Liu et al. [12] give a similar description: “colpi… slightly shorter than pseudocolpi, rhomboid, with narrow endocingulus”. Avetisyan [6] gives the length of colpi and colpori as being identical. This is in accordance with published pictures by Díez et Valdés [9], Bigazzi et al. [13], and Liu et al. [12] (Figure 1(A)). In the genus Kuschakewiczia the type of pollen grains is identical but the shape and character of apertures are reciprocal. The compound apertures (ectoapertures, colpori) are generally narrower (sometimes longer or equally long as to colpi), the simple colpi is almost rhombic, endoapertures are narrower and longer, about 1 × 4 (-5) μm (Figure 1(B)).
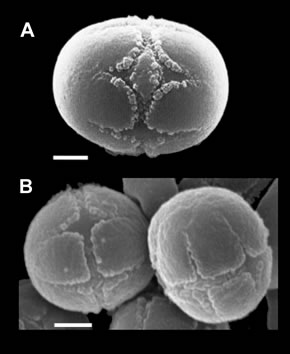
Figure 1. Pollen grains of Solenanthus biebersteinii (A) (Bigazzi et al. [13]) and Kuschakewiczia turkestanica (B) (Scale bars 2 µm).
4. Acknowledgements
This paper appears through financial support provided to the Moravian Museum by the Ministry of Culture of the Czech Republic as part of its long-term conceptual development program for research institutions (ref. MK000094862). My thanks are due to the curators of the herbaria in St. Petersburg for enabling study of their rich material. I also wish to thank Ladislav Ilkovics (Masaryk University, Medical Faculty, Department of Histology and Embryology, Brno, Czech Republic) for providing the SEM microphotographs.
REFERENCES
- E. Regel, “Descriptiones Plantarum Novarum et Minus Cognitarum 6,” Trudy Imperatorskago S.-Peterburgskogo Botanicheskago Sada, Vol. 5, No. 2, 1877, pp. 577-645.
- V. I. Lipsky, “Materialy dla Flory Srednej Azii 2,” Trudy Imperatorskago S.-Peterburgskogo Botanicheskago Sada, Vol. 23, No. 1, 1904, pp. 1-247.
- N. I. Kuznetzow, “Boraginaceae,” In: N. I. Kuznetzow, N. A. Bush and A. V. Fomin, Eds., Flora Caucasica Critica, Vol. 4, No. 2, K. Mattisena, Jurijev, 1913, pp. 66-400.
- J. F. Macbride, “Certain Borraginaceae [sic!], New or Transferred,” Proceedings of the American Academy of Arts and Sciences,” Vol. 51, No. 10, 1916, pp. 541-548. doi:10.2307/20025598
- E. V. Meling, “Polozhenie Solenanthus Turkestanicus (Boraginaceae) v Sisteme Roda, (The Position of Solenanthus Turkestanicus (Boraginaceae) in the System of the Genus),” Botanicheskii Zhurnal, Vol. 67, No. 3, 1982, pp. 348-353.
- E. M. Avetisyan, “Morfologia Microspor Buratschnikovych,” Trudy Botanicheskogo Instituta Akademii Nauk Armyanskoi SSR, Vol. 10, 1956, pp. 7-66.
- G. C. S. Clarke S. Chanda and S. Sahay, “Pollen Morphology in the Genus Pardoglossum (Boraginaceae) with Some Observations on Heterocolpate Pollen,” Review of Paleobotany and Palynology, Vol. 28, No. 3-4, 1979, pp. 301-309. doi:10.1016/0034-6667(79)90030-7
- S. K. Sahay, “Palynotaxonomy of Boraginaceae and Some Other Families of Tubuliflorae,” Biological Memories, Vol. 4, No. 1-2, 1979, pp. 117-205.
- M. J. Díez and B. Valdés, “Pollen Morphology of the Tribes Eritrichieae and Cynoglosseae (Boraginaceae) in the Iberian Peninsula and Its Taxonomic Significance,” Botanical Journal of the Linnean Society, Vol. 107, No. 1, 1991, pp. 49-66. doi:10.1111/j.1095-8339.1991.tb00214.x
- T. Popova and E. A. Zemskova, “Palynomorphology Study of Some Species of Boraginaceae (Subfamily Boraginoidae),” Botanicheskii Zhurnal, Vol. 80, No. 10, 1995, pp. 1-13.
- M. Khatamzas, “Pollen Morphology of Iranian Boraginaceae Family and Its Taxonomic Significance,” Iranian Journal of Botany, Vol. 9, No. 1, 2001, pp. 27-40.
- J.-X. Liu, Y.-L. Zhang, J.-Ch. Ning, Y.-Y. Zhao, Y.-X. Li, J.-M. Zhang and X.-H. Sun, “Pollen Morphology of the Tribe Cynoglosseae of Boraginoideae (Boraginaceae) in China,” Acta Phytotaxonomica Sinica, Vol. 39, No. 6, pp. 515-522.
- M. Bigazzi, E. Nardi and F. Selvi, “Palynological Contribution to the Systematics of Rindera and the Allied Genera Paracaryum and Solenanthus (Boraginaceae-Cynoglosseae),” Willdenowia, Vol. 36, 2006, pp. 37-46. doi:10.3372/wi.36.36103
- E. Barbier and J. Mathez, “Contribution à l’étude des Cynoglossés (Boraginacées): Pardoglossum, Genre Nouveau du Basin Méditerranéen Occidental,” Candollea, Vol. 28, No. 2, 1973, pp. 281-323.
- K. Sutorý, “Oncaglossum, a New Genus of the Boraginaceae, Tribe Cynoglossae, from Mexico,” Novon, Vol. 20, No. 4, 2010, pp. 463-469. doi:10.3417/2008059