Open Journal of Medical Microbiology
Vol.04 No.04(2014), Article ID:51878,7 pages
10.4236/ojmm.2014.44025
Antileishmanial Potential of Piper nigrum Seed Extracts against Leishmania donovani
Garima Chouhan1, Mohammad Islamuddin1, Farnaz Ahmad1, Dinkar Sahal2, Farhat Afrin1,3*
1Parasite Immunology Laboratory, Department of Biotechnology, Faculty of Science, Jamia Hamdard (Hamdard University), New Delhi, India
2Malaria Research Group, International Centre for Genetic Engineering and Biotechnology, Aruna Asaf Ali Marg, New Delhi, India
3Department of Medical Laboratories Technology, Faculty of Applied Medical Sciences, Taibah University, Medina, KSA
Email: *farhatafrin@gmail.com, *afrin_farhat@yahoo.co.in
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 21 September 2014; revised 21 October 2014; accepted 20 November 2014
ABSTRACT
Visceral leishmaniasis (VL) is a pestilent form of leishmaniasis that chiefly impinges the poorest sections of the society. The prototypical therapeutic interventions in vogue are handicapped due to toxicity and alarming increase in drug resistance. In the absence of vaccines, progressive emergence of HIV-VL co-infection and relapse in the form of post kala-azar dermal leishmaniasis, have fuelled the quest for alternative therapies. Herein, we report antileishmanial activity of Piper nigrum, which is endowed with multifarous medicinal properties. Hexane (PNH) and ethanolic (PNE) extracts of P. nigrum substantially inhibited the growth of Leishmania donovani promastigotes with 50% inhibitory concentration (IC50) of 31.6 and 37.8 µg・ml−1, respectively. Growth reversibility analysis revealed the leishmanicidal effect of PNH which caused cell shrinkage and flagellar disruption. In contrast, PNE treated promastigotes showed partial effect. PNH and PNE also abrogated the growth of intra-macrophagic Leishmania amastigotes with IC50 of 14.6 and 18.3 µg・ml−1, respectively. Anti-amastigote efficacy of PNH was accompanied by higher selectivity over host macrophages than PNE. Gas-Chromatography-Mass Spectrometry showed the presence of several secondary metabolites such as trans-β-caryophyllene, piperine, β-bisabolene and other sesquiterpenes in PNH and piperine, δ-(sup 9)-cis oleic acid and piperyline in PNE. Conclusively, our work revealed discernible antileishmanial activity of P. nigrum extracts.
Keywords:
Visceral Leishmaniasis, Leishmania donovani, Piper nigrum, Antileishmanial, Plant Extracts

1. Introduction
Leishmaniasis is a neglected, usually poverty associated complex vector-borne disease that causes huge morbidity and economic losses all over the world. Visceral leishmaniasis (VL), which is the fatal form of leishmaniasis, is responsible for an annual death toll of 60,000 and incidence of 500,000 cases per annum. It is estimated that around 200 million people living in 109 districts in India, Bangladesh, and Nepal are at risk of developing visceral infection. The antiquated chemotherapeutics in use for antileishmanial therapy either cause noxious effects or are rendered ineffective due to growing parasite resistance. Escalation in leishmaniasis burden owing to multiple factors such as co-infection with HIV, occurrence of Post Kala-Azar Dermal Leishmaniasis (PKDL) as relapse infection, lack of vaccine based approaches and difficulties in controlling vectors, have left chemotherapeutic intervention as the sole alternative [1] .
Plants offer ample sources to develop new antileishmanials and replace synthetic drugs. Herbal remedies have been utilized by human population residing in leishmaniasis endemic regions since ancient times. Plants of Piperaceae family have long served the world civilization as wealthy source of ingredients for food spices and traditional medicines. Piper nigrum is a well-known medicinal plant that possesses secondary metabolites with antibacterial, immunomodulatory, cytotoxic, and antimalarial activities [2] . Plant extracts from various species of the genus Piper have demonstrated potent antileishmanial efficacy and leishmanicidal molecules apart from piperine have also been identified [3] - [6] . This prompted us to evaluate the antileishmanial efficacy of P. nigrum seed extracts against L. donovani and to check the presence of plant secondary metabolites other than piperine, which could be responsible for the leishmanicidal effect.
2. Materials
2.1. Parasite Culture and Maintenance
L. donovani strain AG83 (MHOM/IN/83/AG83) was maintained in vivo in BALB/c mice. The promastigote form of the parasites was cultured in Medium 199 (M199), pH 7.4 supplemented with penicillin G sodium (100 U・ml−1), streptomycin sulfate (100 µg・ml−1), HEPES (25 mM) and 10% heat-inactivated FBS (complete M199). Log phase promastigotes were subcultured every 72 - 96 h, in the same medium at 22˚C, inoculum being 2 × 106 cells・ml−1 [7] .
2.2. Plant Material and Preparation of Extracts
P. nigrum dried fruits were purchased locally, and authenticated by Dr. H.B. Singh (voucher no. NISCAIR/ RHMD/Consult/-2010-11/1440/38), Taxonomist, NISCAIR, CSIR, New Delhi. The dried fruits were thoroughly washed with water, shade dried and ground to powder form. The powdered plant material (100 g) was sequentially extracted with solvents of increasing polarity, including hexane, ethanol and water by percolation. The powder was initially soaked in hexane (500 ml) and four washes (250 ml each, at 24 h interval) were performed, followed by extraction in ethanol and water in a similar way. Filtrate at each step was passed through Whatman filter paper (No. 1), pooled and then concentrated to dryness under vacuum by rotary evaporation at 35˚C for hexane (PNH) and ethanolic (PNE) extracts and lyophilized to obtain the aqueous (PNA) extract. The dried extracts were stored at −20˚C until used for bioassay.
2.3. Evaluation of Anti-Promastigote Potential of P. nigrum Extracts and Assessment of Growth Reversibility
Growth kinetics assay was performed to appraise the anti-leishmanial efficacy of P. nigrum extracts. Freshly transformed L. donovani promastigotes (2 × 106 cells・ml−1) were incubated at 22˚C in the presence or absence of test extracts (PNH, PNE and PNA) at 500 µg・ml−1. Piperine, one of the known constituents of Piper species and pentamidine, a standard antileishmanial drug, (positive control used for in vitro studies) were also tested at 500 µg・ml−1 for all the experiments. DMSO (0.5%), which was used to solubilize the extracts, was taken as solvent control to determine any unspecific parasite death. The bioactivity of the extracts was assessed by enumerating the viable parasites under phase contrast microscope (40×) after every 24 h for 7 days. Erythrosin B stain (0.2%, 1:1) was used to distinguish live and dead cells. In a parallel set of experiment, untreated and treated parasites were harvested, washed twice with M199 and finally resuspended in fresh complete M199 at 22˚C. After 96 h, the cell viability was determined by counting the motile, viable, flagellated cells under the microscope [8] .
2.4. Determination of 50% Promastigote Growth Inhibitory Concentration (IC50) of the Bioactive Extracts and Study of Cellular Morphology
Exponential phase promastigotes (2 × 106 ml−1) were incubated in triplicates with the bioactive extracts, piperine, pentamidine or medium alone at serial three fold dilutions (500 to 2.05 µg・ml−1). After 96 h, parasite survival was assessed at different concentrations of the drugs by enumerating the live promastigotes and percent viability calculated according to the formula:
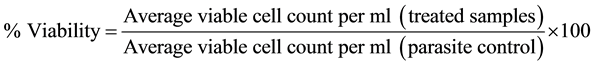
IC50, the concentration that decreased parasite growth by 50% was determined by graphical extrapolation. In parallel, at 96 h, the parasites were harvested, washed twice (3000 ×g, 10 min, 4˚C) with phosphate buffered saline (PBS, 0.02 M, pH 7.4), fixed in 80% chilled ethanol, stained with erythrosin B and observed under Nikon Eclipse 80i microscope (100× objective) for morphological alterations. Since the cells were fixed prior to staining, erythrosin B was not used to distinguish between live and dead cells but for staining the cells for optimal imaging.
2.5. Evaluation of Anti-Amastigote Activity ex Vivo and Determination of Nitric Oxide (NO) Production
Leishmania infected macrophage cells (RAW 264.7) were used to test intracellular efficacy of the bioactive extracts. Macrophages (5 × 106 ml−1, 100 µl per well) in RPMI-1640 medium were plated on round cover slips in 24-well plates. The cells were allowed to adhere for 24 h in carbon dioxide incubator with 5% CO2 at 37˚C. Non adherent cells were removed followed by infection with stationary phase promastigotes, maintaining a Leishmania: macrophage ratio of 10:1. After 24 h, the non-phagocytosed promastigotes were washed with serum-free RPMI 1640 and the infected macrophages were incubated with RPMI 1640 (infected control) or with serial four-fold dilutions of PNH, PNE, piperine (200 to 3.12 µg・ml−1) or pentamidine (100 to 0.39 µg・ml−1) in triplicates at 37˚C. After 48 h, the cover slips were washed with PBS, dried, fixed with chilled methanol (analytical grade) and giemsa-stained. At least 200 macrophages per cover slip from triplicate cultures were counted to calculate the percentage of infected macrophages. Percent amastigote infectivity was determined using the formula:
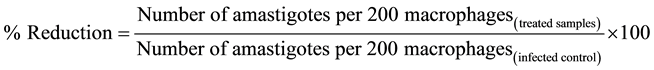
IC50, the concentration of drug that is cytotoxic to 50% of the amastigotes, was obtained by plotting the graph of percent infectivity versus drug concentrations tested [9] . NO was estimated from supernatants of treated and untreated macrophages by Griess reaction [10] .
2.6. Cytotoxic Potential of the Bioactive Extracts and Determination of Selectivity Index
To assess the cytotoxicity, the macrophages (2 × 106 ml−1) in RPMI 1640 were seeded (200 µl・well−1) and left for adherence. After 24 h, the non-adherent macrophages were removed by washing with serum-free RPMI and the adherent macrophages were incubated with the bioactive extracts at serial two fold dilutions (500 to 7.81 µg・ml−1) in triplicates for 48 h. MTT reagent (5 mg・ml−1, 50 µl per well) was added for a further 4 - 6 h, following which the formazan crystals were dissolved in isopropanol:dimethylsulfoxide (1:1) and the absorbance taken in ELISA plate reader at 570 nm. The absorbance by macrophages in the absence of any treatment was considered to be 100%. CC50, the cytotoxic concentration at which 50% of macrophages are viable was calculated according to the formula:
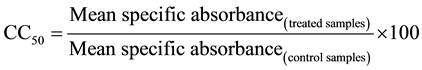
Selectivity index (SI) as a measure of specificity of the leishmanicidal effect of the extracts against internalized parasites over the host macrophages was determined as the ratio of CC50 to IC50 against intramacrophagic amastigotes.
2.7. Gas Chromatography-Mass Spectrometry (GC-MS) of Secondary Metabolites in Bioactive Extracts
The secondary metabolites from PNH and PNE were characterized by GC-MS using a Shimadzu QP2010 with a DB-5 column (30 m, film 0.25 mm, ID 0.25 mm). The temperature of the column was increased gradually from 45˚C to 270˚C at 5˚C・min−1, and the injector and detector temperature for the analysis was about 250˚C. Helium was used as the carrier gas at a flow rate of 1.5 ml・min−1. Identification of the chemical constituents was based on correlation of the recorded mass spectra with those obtained from WILEY8.LIB and NIST08.LIB library spectra provided with the software of the GC-MS system [8] .
2.8. Statistical Analysis
All experiments were performed at least thrice in triplicate. The results shown are from one of three independent experiments and expressed as mean ± standard error of mean (SEM) of the samples in triplicate. Statistical analysis was performed using Graph-Pad software and statistical significance was calculated by one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison test. Differences were considered statistically significant at P < 0.05.
3. Results
3.1. Antiproliferative Effect of PNH and PNE on L. donovani Promastigotes
PNH was the most potent in inducing parasite death. PNE also exhibited killing of L. donovani promastigotes but was less effectual than PNH. Pentamidine, completely arrested the parasite growth after 72 h of incubation (Figure 1(a)). No cell death was apparent in PNA, parasite control, and 0.5% DMSO (solvent control) treated parasites. Piperine initially induced marginal reduction in the promastigote proliferation rate after which the parasites expanded normally.
3.2. Growth Inhibitory Effect of PNH on L. donovani Promastigotes Is Leishmanicidal
The cytocidal (lethal) or cytostatic (growth inhibitory) effect of P. nigrum extracts was confirmed in a growth reversibility assay. Analysis of parasite count after 96 h revealed negligible reversion of growth in PNH treated parasites, indicating that the mode of PNH induced killing was essentially leishmanicidal (Figure 1(b)). However, low parasite count observed in case of PNE treatment reflected its cytostatic effect. High parasite counts were observed in parasites receiving no treatment (parasite control) or upon treatment with 0.5% DMSO (solvent

Figure 1. (a) Antiproliferative effects of P. nigrum extracts against L. donovani promastigotes. Promastigotes (2 × 106 ml−1) were incubated with 500 µg・ml−1 of extracts or 0.5% DMSO (solvent control) and viable cells enumerated at different time points; (b) Growth reversibility analysis of P. nigrum extracts after 96 h. PNH and pentamidine treated promastigotes exhibit no reversion in parasite growth upon drug withdrawal. *P < 0.001 with respect to parasite control.
control) or PNA whereas in pentamidine treated parasites, there was absolutely no reversion in growth of L. donovani promastigotes.
3.3. PNH Induced Morphological Alterations in L. donovani Promastigotes
Morphological changes in L. donovani promastigotes in the presence or absence of Piper extracts were examined microscopically (100×) after 96 h. The photomicrographs depicted that the control untreated parasites retained their long flagella, slender and elongated shape, and were highly motile (Figure 2(a)). PNH treated promastigotes however, had an atypical appearance owing to shortening of flagella, cell shrinkage and consequent circularization. Similar observations were found after pentamidine treatment whereas such changes were less pronounced in PNE treated samples. No such morphological alterations occurred in piperine treated promastigotes.
3.4. IC50 of Bioactive Extracts against L. donovani Promastigotes
To assess IC50 concentration of the bioactive extracts of P. nigrum, parasites were incubated for 96 h with different concentrations of the extracts (2.05 to 500 µg・ml−1) and control was set without any treatment. PNH and PNE induced dose dependent inhibition of parasite growth with IC50 value corresponding to 31.6 and 37.83 µg・ml−1, respectively (Figure 2(b), Table 1).
3.5. P. nigrum Extracts Abrogated Intracellular Amastigote Form of L. donovani in ex Vivo Macrophage―Amastigote Model
The anti-leishmanial activity of PNH and PNE was tested against intracellular amastigotes in L. donovani
 (a) (b)
(a) (b)
Figure 2. (a) Morphological alterations in L. donovani promastigotes after treatment with P. nigrum extracts. Changes in cellular morphology 96 h post treatment with the bioactive extracts, piperine and pentamidine as observed under microscope (100×); (b) Dose-dependent antipromastigote activity of bioactive extracts. Promastigotes (2 × 106 ml−1) were incubated with increasing concentrations of test extracts (2.05 to 500 µg・ml−1) and the IC50 was determined as described in methods. Inset shows mean IC50 values of treated promastigotes.

Table 1. Anti-promastigote and amastigote activity of P. nigrum extracts and determination of Selectivity Index (SI).
SI of bioactive extracts was determined as a measure of their toxicity against RAW 264.7 macrophages. SI = CC50 against macrophages/IC50 against amastigotes, SE = Standard error.
infected RAW 264.7 cells in comparison to pentamidine (100 to 0.39 µg・ml−1), owing to its cytotoxicity. Percent inhibition for each group (PNH, PNE, piperine, and pentamidine) was plotted in a concentration-response curve to calculate the IC50 as elaborated in methods. PNH and PNE exhibited a dose dependent reduction in parasite load with IC50 of 14.63 and 18.83 µg・ml−1, respectively. The IC50 of pentamidine was found to be 0.84 µg・ml−1 and that of piperine was achieved at 41.33 µg・ml−1 (Figure 3, Table 1), which is indicative of its moderate antileishmanial activity.
3.6. Antileishmanial Activity of P. nigrum Extracts Is Independent of NO Generation
Since NO is one of the major effector microbicidal molecules responsible for clearance of Leishmania amastigotes, we probed the role of NO in PNH and PNE induced death. We found that our bioactive extracts along with piperine and pentamidine failed to generate NO in L. donovani infected RAW macrophages (P > 0.05, data not shown).
3.7. Cytotoxicity of P. nigrum Extracts towards Mammalian Macrophages
To assess the safety of bioactive extracts of P. nigrum against macrophages, the RAW 264.7 cells were incubated with increasing concentrations of PNH and PNE (0 to 500 µg・ml−1). In order to examine the specificity of these extracts against Leishmania amastigotes over host macrophages, the SI was calculated as a ratio of CC50 against macrophages to IC50 against amastigotes. PNH was found to be most selective with SI of 25.95 (Table 1). PNE exhibited SI = 10.91, whereas piperine was non-toxic against RAW macrophages.
3.8. GC-MS Analysis of PNH and PNE
GC-MS profiling of PNH and PNE was performed to identify the leishmanicidal plant secondary metabolites in these bioactive extracts. PNH constituted 52 compounds, the most abundant being trans-β-caryophyllene (22.28%). The other constituents included β-bisabolene (6.08%), tetrahydropiperine (5.66%), 9Z-9-tetra-decen- al (5.32%), α-copaene (4.76%), 3-amino-4-piperonyl-5-pyrazolone (4.14%), 1,1-dimethyldecahydronap-thalene (3.42%), 14-methyl-8-hexadcyn-1-ol (3.04%), δ-cadinol (2.21%), 1-methyldecahydronapthalene (2.75%), α-caryophyllene (2.06%), 1-(12-octadecanoyl) pyrrolidine (2.03%), δ-cadinene (1.81%), α-selinene (1.80%), β-elemene (1.62%) and elemol (1.37%). Piperine content of PNH was observed to be 8.65% (Supplementary

Figure 3. Antiamastigote efficacy of PNH and PNE. Antiamastigote efficacy was evaluated in RAW macrophages parasitized with L. donovani promastigotes as described in methods. *P < 0.001 with respect to infection control.
File 1). However, PNE was dominated by piperine (70.36%) though other components such as δ-(sup 9)-cis oleic acid (3.71%), piperoylpiperidine (3.70%), piperyline (3.02%), 3-amino-4-iperonyl-5-pyrazolone (2.13%), tetrahydropiperine (1.64%), trans-β-caryophyllene (1.33%) were also detected (Supplementary File 2).
4. Discussion
In the present work, we found that hexane (PNH) and ethanolic (PNE) extracts of P. nigrum were found to be more effectual in reducing in vitro proliferation of L. donovani promastigotes and amastigotes in comparison to piperine. Our data elaborated the cytocidal effect of PNH that is preferred over cytostatic mode of cell death as evident with PNE, for advantages such as rapid elimination of parasites, lesser risk of resistance development or resurgence of infection. Alterations in parasite morphology were also observed and the irreversible changes in morphology accompanying minimal growth reversion further corroborated the leishmanicidal action of PNH in comparison to PNE. Pentamidine was used as a known antileishmanial, that has been earlier reported by us [8] [9] and others [11] for in vitro studies.
Further, the antiamastigote efficacy of the extracts was independent of NO generation. Trans-β-caryophyllene, a major plant secondary metabolite found in PNH has also been reported to be active against L. amazonensis in NO independent manner [12] . Piperine also failed to induce NO production and similar observation has also been demonstrated by others [13] . SI values were calculated to assess the adverse cytotoxicity of bioactive extracts and SI values above 10 were considered specific in accordance with those reported for antiprotozoal compounds including Leishmania [14] . In accordance, PNH was found to be highly specific and least toxic, whereas PNE was found to be moderately specific.
GC-MS analysis of PNH revealed that the major portion of the extract was constituted by cyclic sesquiterpenes. Leishmanicidal activity of this class of compounds has been well documented [15] . Tetrahydropiperine, a derivative of piperine present in PNH (5.66%) and to a lesser extent in PNE (1.64%) has exhibited antileishmanial activity against L. amazonensis [13] . In addition to being antileishmanial, trans-β-caryophyllene has also been reported to be antitrypanosomal [16] and antitumoricidal [17] . Leishmanicidal potential of piperine was reported as early as 1993 against an attenuated L. donovani UR6 strain where it completely inhibited its growth after 5 days of incubation at 1.2 mg・ml−1 [18] . IC50 of piperine against L. donovani promastigotes has been reported to be 2.558 mM [19] . We also found that piperine was moderately active against L. donovani promastigotes even at 500 µg・ml−1. Thus, the GC-MS data further demonstrates that P. nigrum extracts are a source of potent leishmanicidal plant secondary metabolites besides piperine.
5. Conclusion
Conclusively, our studies have demonstrated promising antileishmanial potential of P. nigrum extracts. Hexane (PNH) and ethanolic (PNE) extracts exhibited antileishmanial activity with PNH being highly selective towards intracellular stage of the parasite. Thus, the active principles present in P. nigrum seeds may be a source of promising leishmanicidal compounds and may serve as structural templates for synthesis of various synthetic and semi-synthetic derivatives with improved antileishmanial activity.
Acknowledgements
This work received financial assistance from Department of Biotechnology, Central Council for Research in Unani Medicine and Department of Science and Technology, Government of India. G.C. is a recipient of Basic Science Research Fellowship from University Grants Commission. We are obliged to Dr. N. Ali, Indian Institute of Chemical Biology, Kolkata, India, for providing us L. donovani strain. We are thankful to Dr. A. Kumar, Jawaharlal Nehru University, Delhi, India, for his valuable help in carrying out GC-MS analysis.
References
- Chouhan, G., Islamuddin, M., Sahal, D. and Afrin, F. (2014) Exploring the Role of Medicinal Plant Based Immunomodulators for Effective Therapy of Leishmaniasis. Frontiers in Immunology, 5, 193. http://dx.doi.org/10.3389/fimmu.2014.00193
- Rahmatullah, M., Hasan, A., Parvin, W., Moniruzzaman, M., Khatun, A., Khatun, Z., Jahan, F.I. and Jahan, R. (2012) Medicinal Plants and Formulations Used by the Soren Clan of the SantalTribe in Rajsahi District, Bangladesh for Treatment of Various Ailments. African Journal of Traditional, Complementary and Alternative Medicine, 9, 350-359. http://dx.doi.org/10.4314/ajtcam.v9i3.8
- Hermoso, A., Jiménez, I.A., Mamani, Z.A., Bazzocchi, I.L., Piñero, J.E., Ravelo, A.G. and Valladares, B. (2003) Antileishmanial Activities of Dihydrochalcones from Piper elongatum and Synthetic Related Compounds. Structural Requirements for Activity. Biorganic and Medicinal Chemistry, 11, 3975-3980. http://dx.doi.org/10.1016/S0968-0896(03)00406-1
- Carrara, V.D.S., Cunha-Júnior, E.F., Torres-Santos, E.C., Corrêa, A.G., Monteiro, J.L., Demarchi, I.G., Lonardoni, M.V.C. and Cortez, D.A.G. (2013) Antileishmanial Activity of Amides from Piper amalago and Synthetic Analogs. Revista Brasiliera de Farmacognosia, 23, 447-454. http://dx.doi.org/10.1590/S0102-695X2013005000022
- Sahi, S., Tewatia, P. and Ghosal, S. (2012) Leishmania donovani Pteridine Reductase 1: Comparative Protein Modeling and Protein-Ligand Interaction Studies of the Leishmanicidal Constituents Isolated from the Fruits of Piper longum. Journal of Molecular Modeling, 18, 5065-5073. http://dx.doi.org/10.1007/s00894-012-1508-y
- Sen, R. and Chaterjee, M. (2011) Plant Derived Therapeutics for the Treatment of Leishmaniasis. Phytomedicine, 18, 1056-1069. http://dx.doi.org/10.1016/j.phymed.2011.03.004
- Afrin, F., Rajesh, R., Anam, K., Gopinath, M., Pal, S. and Ali, N. (2002) Characterization of Leishmania donovani Antigens Encapsulated in Liposomes That Induce Protective Immunity in BALB/c Mice. Infection Immunity, 70, 6697- 6706. http://dx.doi.org/10.1128/IAI.70.12.6697-6706.2002
- Islamuddin, M., Farooque, A., Dwarakanath, B.S., Sahal, D. and Afrin, F. (2012) Extracts of Artemisia annua Leaves and Seeds Mediate Programmed Cell Death in Leishmania donovani. Jounal of Medical Microbiology, 61, 1709-1718. http://dx.doi.org/10.1099/jmm.0.049387-0
- Islamuddin, M., Sahal, D. and Afrin, F. (2014) Apoptosis-Like Death in Leishmania donovani Promastigotes Induced by Eugenol-Rich Oil of Syzygium aromaticum. Journal of Medical Microbiology, 63, 74-85. http://dx.doi.org/10.1099/jmm.0.064709-0
- Githinji, E.K., Irungu, L.W., Tonui, W.K., Rukunga, G.M., Mutai, C., Muthaura, C.N., Lugalia, R., Gikandi, G., Wainaina, C.W., Ingonga, J.M. and Wanjoya, A. (2010) In Vitro Effects of Warburgia ugandensis, Psiadia punctulata and Chasmanthera Dependes on Leishmania Major Promastigotes. African Journal of Traditional, Complementary and Alternative Medicine, 7, 264-275. http://dx.doi.org/10.4314/ajtcam.v7i3.54791
- Benítez, J., Cavalcanti de Queiroz, A., Correia, I., Alves, M.A., Alexandre-Moreira, M.S., Barreiro, E.J., Lima, L.M., Varela, J., González, M., Cerecetto, H., Moreno, V., Pessoa, J.C. and Gambino, D. (2013) Newoxivanadium (IV) N- Acylhydrazone Complexes: Promising Antileishmanial and Antitrypanosomal Agents. European Journal of Medicinal Chemistry, 62, 20-27. http://dx.doi.org/10.1016/j.ejmech.2012.12.036
- Soares, D.C., Portella, N.A., De S. Ramos, M.F., Siani, A.C. and Saraiva, E.M. (2013) Trans-β-Caryophyllene: An Effective Antileishmanial Compound Found in Commercial Copaiba Oil (Copaifera spp.). Evidence-Based Complementary and Alternative Medicine, 2013, Article ID: 761323. http://dx.doi.org/10.1155/2013/761323
- Ferreira, C., Soares, D.C., Barreto-Junior, C.B., Nascimento, M.T., Freire-de-Lima, L., Delorenzi, J.C., Lima, M.E., Atella, G.C., Folly, E., Carvalho, T.M., Saraiva, E.M. and Pinto-da-Silva, L.H. (2011) Leishmanicidal Effects of Piperine, Its Derivatives, and Analogues on Leishmania amazonensis. Phytochemistry, 72, 2155-2164. http://dx.doi.org/10.1016/j.phytochem.2011.08.006
- Osorio, E., Arango, G.J., Jiménez, N., Alzate, F., Ruiz, G., Gutiérrez, D., Paco, M.A., Giménez, A. and Robledo, S. (2007) Antiprotozoal and Cytotoxic Activities in Vitro of Colombian Annonaceae. Journal of Ethnopharmacology, 111, 630-635. http://dx.doi.org/10.1016/j.jep.2007.01.015
- Odonne, G., Herbette, G., Eparvier, V., Bourdy, G., Rojas, R., Sauvain, M. and Stien, D. (2011) Antileishmanial Sesquiterpene Lactones from Pseudelephantopus spicatus, a Traditional Remedy from the Chayahuita Amerindians (Peru). Part III. Journal of Ethnopaharmacology, 137, 875-879.
- Izumi, E., Ueda-Nakamura, T., Veiga Jr., V.F., Pinto, A.C. and Nakamura, C.V. (2012) Terpenes from Copaifera De- monstrated in Vitro Antiparasitic and Synergic Activity. Journal of Medicinal Chemistry, 55, 2994-3001. http://dx.doi.org/10.1021/jm201451h
- Amiel, E., Ofir, R., Dudai, N., Soloway, E., Rabinsky, T. and Rachmilevitch, S. (2012) β-Caryophyllene, a Compound Isolated from the Biblical Balm of Gilead (Commiphora gileadensis), Is a Selective Apoptosis Inducer for Tumor Cell Lines. Evidence-Based Complementary and Alternative Medicine, 2012, 872394. http://dx.doi.org/10.1155/2012/872394
- Kapil, A. (1993) Piperine: A Potent Inhibitor of Leishmania donovani Promastigotes in Vitro. Planta Medica, 59, 474. http://dx.doi.org/10.1055/s-2006-959737
- Singh, I.P., Jain, S.K., Kaur, A., Singh, S., Kumar, R., Garg, P., Sharma, S.S. and Arora, S.K. (2010) Synthesis and Antileishmanial Activity of Piperoyl-Amino Acid Conjugates. European Journal of Medicinal Chemistry, 45, 3439- 3445. http://dx.doi.org/10.1016/j.ejmech.2010.04.033
NOTES

*Corresponding author.