Open Journal of Nephrology
Vol.4 No.1(2014), Article ID:44087,8 pages DOI:10.4236/ojneph.2014.41004
No Correlation between AVPR1A Promoter Polymorphisms and Prepulse Inhibition in Patients with Nocturnal Enuresis
Sebastian Schulz-Juergensen1*, Philipp von Bismarck1, René Santer2, Paul Eggert1
1Department of General Pediatrics, University Hospital Schleswig-Holstein, Campus Kiel, Kiel, Germany
2Department of Pediatrics, University Medical Center Eppendorf, Hamburg, Germany
Email: *schulz-juergensen@pediatrics.uni-kiel.de
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/


Received 28 January 2014; revised 25 February 2014; accepted 10 March 2014
ABSTRACT
Introduction: A correlation between AVPR1A promoter polymorphisms and prepulse inhibition (PPI) of startle reflexes has been described in healthy adults. Many children with nocturnal enuresis (NE) have a reduced PPI and treatment with desamino arginine vasopressin (dDAVP), a ligand of the arginine vasopressin receptor 1A (AVPR1A), and both improve clinical symptoms and significantly increase PPI. Methods: In 17 children (median 9.1 years, range 6.4 - 17.3) with NE, promoter repeats within the RS1 and RS3 regions of AVPR1A were quantified and correlated to PPI (native and age-adjusted). Results: No direct correlation was found between the number of promoter repeats at RS1 and PPI (correlation coefficient—0.240, p = 0.346) or RS3 and PPI (correlation coefficient—0.0192, p = 0.936), with no change through age-adjustment of PPI. The different RS3 length subgroups did not show differences in PPI, nor did differentiation of NE according to clinical subtype or treatment response to dDAVP show differences in the number of promoter repeats. Conclusion: The missing reproducibility of the correlation between AVPR1A promoter polymorphisms and PPI in a group with wide range of PPI suggests a more complex interaction. Therefore, further investigations are needed to analyze this very plausible interaction. Conditions with a reduced PPI, such as enuresis, schizophrenia or autism, are particularly interesting for this research.
Keywords:AVPR1A Promoter; Prepulse Inhibition; Nocturnal Enuresis; Correlation

1. Introduction
Both prepulse inhibition (PPI) and blood concentrations of arginine vasopressin (AVP) have been associated with modulation of social behaviour in different species including humans [1] -[4] . PPI, measured as the attenuation of the startle reflex by a shortly preceding prepulse, is used as a marker for sensorimotor gating [5] , and has repeatedly been shown to be impaired in psychiatric disorders, e.g., schizophrenia or autism [6] [7] .
In recent years, modulation of expression of the gene of the central arginine vasopressin receptor 1a (AVPR1a, gene symbol AVPR1A) and of the effect of AVP on this receptor has been attributed to differences in the promoter region of AVPR1A [8] . Particularly, different numbers of repeats in the regions RS1 and RS3 have shown to have an influence on social behavior in animals [9] and humans [10] .
At present, only one study has been published that describes correlations of the number of repeats within AVPR1A and PPI [11] . Possibly limited by a limited range of PPI in a population-based approach, these correlations could only be demonstrated by using sophisticated and non-conservative statistics.
In this context, patients with nocturnal enuresis (NE) represent an interesting group with a specific phenotype: a large proportion has been repeatedly shown to have reduced PPI [12] [13] , and show both clinical improvement as well as improvement of PPI through desamino arginine vasopressin (dDAVP) [14] , suggesting an effect of dDAVP on central reflex control via the central AVP-receptor AVPR1a.
Additionally, the effect of dDAVP is correlated to the baseline PPI. Only patients with a reduced baseline PPI (clinically those with a non-monosymptomatic enuresis, NMNE) benefit from dDAVP, while patients with normal baseline PPI (clinically those with strictly monosymptomatic enuresis, MNE) do not show further improvement of PPI and also have no clinical improvement [15] .
Therefore, this study was designed to investigate a possible correlation of the number of repeats in the RS1 and RS3 promoter regions of AVPR1A and PPI in a cohort of clinically well-defined patients with nocturnal enuresis. Furthermore, the number of repeats was correlated with clinical subtypes defined by daytime symptoms, family history and treatment response to dDAVP.
2. Materials and Methods
2.1. Patients
Seventeen patients with a diagnosis of nocturnal enuresis were recruited through our Pediatric Nephrology clinic between 04/2008 and 02/2010. Inclusion criteria were a diagnosis of MNE or NMNE, with the exclusion of malformations of the urinary tract as well as neurological or psychological impairments. The group consisted of 13 boys and 4 girls with a median age of 9.1 years (range 6.4 - 17.3 years). The individual patient characteristics are included in the summarizing table of results (Table 1). A family history regarding parental enuresis was also obtained.
2.2. Stimulation/Startle Reaction and Procedure
A tone generator and EMG measurement instruments (Contact Precision Instruments®) were used. A white noise of 50 dB SPL was continuously applied as a background hum and was replaced without interruption by the presented stimuli. The startle stimulus consisted of a white-noise burst (105 dB, 40 ms duration, 20 - 20,000 Hz, <1 ms rise and fall time) and the prepulse stimulus of a 1000 Hz-tone (75 dB, 25 ms duration, <1 ms rise and fall time). The stimuli were presented binaurally through SONY-MDR-CD470-earphones. During each session startle stimuli were presented as follows: 1) startle alone and 2) startle proceeded by prestimulation tone 60 ms (“prepulse + startle”) before the startle stimulus. After two minutes of adaptation to the recording room three “startle alone” stimuli were administered to reach a stable individual habituation level of startle reactivity. Then a total of 20 impulses followed in a pseudo-randomized sequence of which ten were “startle alone” stimuli (1) and ten “prepulse” (2). The startle amplitude of these 20 stimuli was used for computing the startle response and the startle reflex control. The average inter-stimulus interval was 20 seconds. The probands were informed that they would hear noises over headphones while watching a silent cartoon, but had no instructions to pay attention or to react to the stimuli (unattended startle and prepulse stimuli).
2.3. EMG Measurement and Calculation of PPI
The startle response was measured by electromyography (EMG) of the M. orbicularis oculi. The EMG was rec

Table 1. Summary of patient characteristics and results. Age is given in years. Symptoms are categorized into “only night” with strictly monosymptomatic NE, without day symptoms or urge, and “day and night” in case of day symptoms with only urge or day wetting. dDAVP response was graded into “good” for reduction of wet nights ≥ 90%, “moderate” for reduction of wet nights between 50% - 89% and “poor” for reduction of wet nights ≤ 49%. “Resolution” indicates the age at which the patient was dry under therapy. “Family history” indicates eventual parent also affected by NE. PPI is given as percentage as indicated in the methods. The lengths of PCR products of RS1 and RS3 promoter repeats are given, with long repeat number being marked in bold lettering. 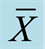 RS1 and
RS1 and 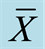 RS3 indicate the average repeat numbers, respectively.
RS3 indicate the average repeat numbers, respectively.
orded continuously bipolarly from silver/silver chloride electrodes taped to the skin (4 mm² for EMG, 8 mm² for reference electrode). Two EMG electrodes were fixed 15 mm apart from each other (centre to centre) and 12 mm below the left lower eyelid and a reference electrode on the forehead. The EMG raw signal was amplified, filtered (high and low pass filters at 30 - 500 Hz) and converted to a digital signal by an analogue/digital converter. The signal was recorded at a sampling rate of 1000 Hz and stored on a PC.
The digital EMG signal was integrated by an automatic signal centering system to remove any DC offset, rectified and smoothed (running average with a time constant of 4 ms) before measuring the startle response (startle amplitude). The EMG base line and the maximum amplitude after a startle stimulus with or without prepulse were calculated from the integrated, smoothed EMG signal. The base line was calculated as the average amplitude of the 200 ms before onset of the startle stimulus. A startle reaction was defined as an increase above mean baseline of the EMG starting between 20 and 80 ms after the stimulus and reaching a maximum within 20 - 120 ms. The startle amplitude was calculated from the difference between the peak amplitude and the baseline of each startle stimulus. The individual startle response was calculated as the average startle reactivity of the available trials for the respective condition. Following the “Committee report for human startle eyeblink EMG studies” [16] , a test person was classified as a responder when at least one startle reaction to one of the first three startle alone stimuli was above two standard deviations of the EMG baseline, and the following trials were used for data analysis independent of individual startle amplitude. If no startle reaction was registered, the test person was classified as a non-responder and excluded from the data analysis. A single startle response to a startle stimulus defined as written above during one session was considered as invalid when there were blinks or artefacts 200 ms before and 20 ms after the startle stimulus. These single responses were classified as “invalids”. The individual startle reactivity was not computed when there were >3 invalid responses (“invalids”) to startle or prestimulation stimuli.
The startle reflex control (PPI) was quantified as the percentage reduction of the averaged startle response in the pre-stimulation condition (“prepulse + startle” stimuli) compared to the control condition (“startle alone” stimuli). It was calculated using the following formula:
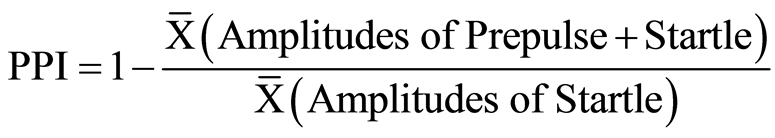
2.4. Genetic Investigations
DNA was obtained from all family members and extracted according to standard methods. PCR amplification of the polymorphic RS1 and RS3 regions of the AVPR1A promoter was achieved using the following pair of primers, originally recommended by Levin et al. [11] :
RS1: sn-(fluorescent)-5’-AGG GAC TGG TTC TAC AAT CTG C-3’ and asn-5’-ACC TCT CAA GTT ATG TTG GTG G-3’; RS3: sn-(fluorescent)-5’-CCT GTA GAG ATG TAA GTG CT-3’ and asn-5’-TCT GGA AGA GAC TTA GAT GG-3’.
The PCR products were analyzed on an ABI/Hitachi 3500 Genetic Analyser (Life Techniologies, Applied Biosystems, Darmstadt, Germany).
2.5. Presentation and Analysis of Data
Box plots are used to present PPI data with median (line), interquartile range (box), 10th and 90th percentile (whiskers), and outliers (dots).
In order to describe the AVPR1A promoter polymorphisms at RS1 and RS3, the lengths of PCR products obtained with the primers described above are presented.
Correlations were investigated using the non-parametric Spearman rank order correlation test.
Comparisons between subgroups were analyzed using the non-parametric Mann-Whitney test, with p < 0.05 considered significant.
Sigma Plot 11 was used as statistical software.
2.6. Ethical Approval and Informed Consent
The study was approved by the ethics committee of the Christian-Albrechts-University, Kiel. The patients and their parents gave informed consent to participate in the study.
3. Results
Table 1 gives an overview of the individual patient’s results. Findings of PPI testing are presented together with the length of the PCR products, an indirect measure of the repeat numbers in the RS1 and RS3 regions, and with clinical information regarding age, symptoms, treatment response, and family history.
The following correlations were evaluated:
1) RS1/RS3 repeats and native PPI There is no correlation between PPI and the average number of repeats within the RS1 region or PPI and average number of repeats within the RS3 region of AVPR1A, see Table 2. Using lower or higher repeat number within the RS1 and RS3 regions instead of average repeat number, there is also no correlation between PPI and number of repeats, respectively (data not shown).
2) RS1/RS3 repeats and age-adjusted PPI If individual PPI is calculated as percentage of age-related normal value [17] , this age-adjusted PPI shows no significant correlation with the average number of RS1 and RS3, see Table 2.
3) PPI in RS3 subgroups If the RS3 repeats are graded into short (S) and long (L) with the median of 326 repeats as cut-off and the patients assigned to SS-, SLand LL-subgroups (as suggested by Knafo et al. [18] and Levin et al. [11] , PPI is different in the subgroups, with lowest PPI in the SL group (see Figure 1). This difference is not statistically significant. The same applies to a differentiation according to RS1 repeats with the median of 311 repeats as cut-off. The median PPIs for the SS, SL and LL subgroups were 54%, 53% and 18%, respectively, again without statistically significant difference (data not shown).
4) RS1/RS3 repeats and PPI in clinical subgroups (MNE vs. NMNE)
The patients were clinically classified into patients with strictly nocturnal, monosymptomatic enuresis (MNE
Table 2. Correlations between PPI (without and with age-adjustment) and RS1 as well as RS3 repeats, respectively. P-values are calculated using the Spearman rank order correlation..

Figure 1. PPI in the subgroups with repeats classified in the subgroups short/short (SS), short/long (SL) and long/long (LL). Data are given as median and range, p values are calculated using the non-parametric Wilcoxon-signed-rank test.
n = 9) and those with daytime symptoms, non-monosymptomatic enuresis (NMNE, n = 8). The comparisons of median RS1 and RS3 repeats as well as median PPI are given in Table 3.
5) RS1/RS3-Repeats and PPI differentiated according to treatment response to dDAVP The patients were clinically classified into patients with good response to dDAVP (reduction of wet nights ≥ 90%, n = 3) and moderate/poor response to dDAVP (reduction of wet nights < 90%, n = 14). The comparisons of median RS1 and RS3 lengths as well as median PPI are given in Table 4.
6) RS1/RS3-Repeats and PPI differentiated according to family history The patients were clinically classified into patients with a positive family history (father and/or mother suffering from NE in childhood, n = 9) and negative family history (no history of NE in family, n = 8). The comparisons of median RS1 and RS3 repeats as well as median PPI are given in Table 5.
4. Discussion
A genetic background of nocturnal enuresis has long been postulated (e.g., Frary [19] ), with a relevant proportion of patients having a positive family history for this disturbance [20] . More recently, interesting gene loci for NE have been identified by linkage analyses as there are ENUR1 in the chromosomal region 13q13 - q14.3 [21] , ENUR2 in 12q13-q21 [22] and ENUR3 in 22q11 [23] . Within ENUR2 lies AVPR1A, the gene encoding the arginine vasopressin receptor type 1a, which has been located to chromosomal region 12q14.2 [24] .
While dDAVP is known to be a ligand of the main renal vasopressin receptor (type 2, AVPR2), two studies could demonstrate that it also has clinical effects in patients with diabetes insipidus and mutations in the respective gene [25] [26] , suggesting that other AVP receptors are involved in the mode of action of dDAVP.
The described association of the number of AVPR1A promoter repeats and PPI [11] and the effect of dDAVP on PPI in nocturnal enuresis [14] therefore strongly suggested genetic variations of the AVPR1a receptor as pathogenetic factors in nocturnal enuresis.
This association was investigated in a pediatric cohort of enuresis patients with the advantage of a wide range of PPIs and at least some therapeutic effect of dDAVP, on the other hand, the reduced sample size was definitively the main limiting factor. The missing correlation of PPI and the number of RS1 or RS3 repeats in this cohort can possibly be explained by the small sample size, but the correlation coefficients do not suggest a mere lack of number of investigated subjects. Given the clinical heterogeneity of NE, this association could be present in one of the subgroups defined by clinical subtype, treatment response to dDAVP, or family history. The small sample size in this study, however, prevents formation of subgroups with sufficient statistical power, reflected in the non-significant p values in these analyses.
Table 3. Median RS1 and RS3 length in base-pairs (bp) and PPI in the clinical subgroups with monosymptomatic (MNE) or non-monosymptomatic (NMNE) enuresis. In the last column, the p value for the difference calculated with WilcoxonSigned-Rank test is given.
Table 4. Median RS1 and RS3 length in base-pairs (bp) and PPI in the clinical subgroups with good (n = 3) and poor (n = 14) response to dDAVP. In the last column, the p-value for the difference calculated with Wilcoxon-Signed-Rank test is given.
Table 5. Median RS1 and RS3 length in base-pairs (bp) and PPI in the clinical subgroups with positive and negative family history. In the last column, the p-value for the difference calculated with Wilcoxon-Signed-Rank test is given.
Looking at the trends found in the subgroup analyses, the lower PPI in the NMNE subgroup and in the patients with good dDAVP response is consistent with previous findings [15] [27] . However, the trend towards higher number of RS3-repeats in the NMNE subgroup is not in accordance with the expectations from the study by Levin et al. [11] . This subgroup would rather be expected to have lower number of RS3 repeats associated with a lower PPI. This finding should be further investigated in a larger cohort, also focusing on exact clinical classifications of NE subgroups. In view of the missing overall association of number of RS3 repeats and PPI in this cohort with a wide range of PPIs, the findings of Levin et al. [11] , based on explicitly non-conservative statistics, need to be reproduced in a larger sample.
5. Conclusion
In conclusion, a simple and direct association of the number of repeats within the RS3 region of AVPR1A and different PPIs in NE or its subgroups (MNE, NMNE) could not be found in this relatively small cohort. However, the approach of investigating microsatellite polymorphisms within RS1 and RS3 of AVPR1A in a disturbance strongly was associated with reduced PPI and arginine vasopressin agonists like dDAVP, possibly also including further analyses in the coding regions of AVPR1A or of markers of AVPR1A gene expression. In view of PPI changes during childhood and ethical concerns about genetic investigations in children, other clinical conditions that affect adults and are known to be associated with reduced PPI (e.g., schizophrenia, autism and Gilles de la Tourette syndrome) may also help to further understand this interesting association. The idea of transferring correlations between genetic parameters and neurophysiologic parameters found in healthy subjects to cohorts with more specific and “extreme” phenotypes may generally be useful to reconfirm such correlations.
Acknowledgements
We thank Dr. Julia Gebhardt who performed the startle and PPI measurements.
Conflict of Interest
The authors declare that they have no conflict of interest.
References
- Naert, A., Callaerts-Vegh, Z. and D’Hooge, R. (2012) Nocturnal Hyperactivity, Increased Social Novelty Preference and Delayed Extinction of Fear Responses in Post-Weaning Socially Isolated Mice. Brain Research Bulletin, 85, 354- 362. http://dx.doi.org/10.1016/j.brainresbull.2011.03.027
- Giakoumaki, S.G. (2012) Cognitive and Prepulse Inhibition Deficits in Psychometrically High Schizotypal Subjects in the General Population: Relevance to Schizophrenia Research. Journal of the International Neuropsychological Society, 18, 643-656.
- Semsar, K., Kandel, F.L. and Godwin, J. (2001) Manipulations of the AVT System Shift Social status and Related Courtship and Aggressive Behavior in the Bluehead Wrasse. Hormones and Behavior, 40, 21-31. http://dx.doi.org/10.1006/hbeh.2001.1663
- Meyer-Lindenberg, A., Domes, G., Kirsch, P. and Heinrichs, M. (2011) Oxytocin and Vasopressin in the Human Brain: Social Neuropeptides for Translational Medicine. Nature Reviews Neuroscience, 12, 524-538. http://dx.doi.org/10.1038/nrn3044
- Braff, D.L. and Geyer, M.A. (1990) Sensorimotor Gating and Schizophrenia. Human and Animal Model Studies. Archives of General Psychiatry, 47, 181-188. http://dx.doi.org/10.1001/archpsyc.1990.01810140081011
- Parwani, A, Duncan, E.J., Bartlett, E., Madonick, S.H., Efferen, T.R., Rajan, R., Sanfilipo, M., Chappell, P.B., Chakravorty, S., Gonzenbach, S., Ko, G.N. and Rotrosen, J.P. (2000) Impaired Prepulse Inhibition of Acoustic Startle in Schizophrenia. Biological Psychiatry, 47, 662-669. http://dx.doi.org/10.1016/S0006-3223(99)00148-1
- Perry, W., Minassian, A., Lopez, B., Maron, L. and Lincoln, A. (2007) Sensorimotor Gating Deficits in Adults with Autism. Biological Psychiatry, 61, 482-486. http://dx.doi.org/10.1016/j.biopsych.2005.09.025
- Hammock, E.A. and Young, L.J. (2002) Variation in the Vasopressin V1a Receptor Promoter and Expression: Implications for Interand Intraspecific Variation in Social Behaviour. European Journal of Neuroscience, 16, 399-402. http://dx.doi.org/10.1046/j.1460-9568.2002.02083.x
- Hopkins, W.D., Donaldson, Z.R. and Young, L.J. (2012) A Polymorphic Indel Containing the RS3 Microsatellite in the 5’ Flanking Region of the Vasopressin V1a Receptor Gene Is Associated with Chimpanzee (Pan troglodytes) Personality. Genes, Brain and Behavior, 11, 552-558. http://dx.doi.org/10.1111/j.1601-183X.2012.00799.x
- Walum, H., Westberg, L., Henningsson, S., Neiderhiser, J.M., Reiss, D., Igl, W., Ganiban, J.M., Spotts, E.L., Pedersen, N.L., Eriksson, E. and Lichtenstein, P. (2008) Genetic Variation in the Vasopressin Receptor 1a Gene (AVPR1A) Associates with Pair-Bonding Behavior in Humans. Proceedings of the National Academy of Sciences, 105, 14153-14156. http://dx.doi.org/10.1073/pnas.0803081105
- Levin, R., Heresco-Levy, U., Bachner-Melman, R, Israel, S., Shalev, I. and Ebstein, R.P. (2009) Association between Arginine Vasopressin 1a Receptor (AVPR1a) Promoter Region Polymorphisms and Prepulse Inhibition. Psychoneuroendocrinology, 34, 901-908. http://dx.doi.org/10.1016/j.psyneuen.2008.12.014
- Ornitz, E.M., Russell, A.T., Hanna, G.L., Gabikian, P., Gehricke, J.G., Song, D. and Guthrie, D. (1999) Prepulse Inhibition of Startle and the Neurobiology of Primary Nocturnal Enuresis. Biological Psychiatry, 45, 1455-1466.
- Baeyens, D., Roeyers, H., Naert, S., Hoebeke, P. and VandeWalle, J. (2007) The Impact of Maturation of Brainstem Inhibition on Enuresis: A Startle Eye Blink Modification Study with 2-Year Followup. Journal of Urology, 178, 2621- 2625. http://dx.doi.org/10.1016/j.juro.2007.07.061
- Schulz-Juergensen, S., Rieger, M., Schaefer, J., Neusuess, A. and Eggert, P. (2007) Effect of 1-Desamino-8-D-Arginine Vasopressin on Prepulse Inhibition of Startle Supports a Central Etiology of Primary Monosymptomatic Enuresis. Journal of Pediatrics, 151, 571-574. http://dx.doi.org/10.1016/j.jpeds.2007.05.024
- Eggert, P., Freischmidt, S., Bismarck, P.V. andSchulz-Jürgensen, S. (2012) Differentiation of Treatment Options for Enuresis by Measurement of Prepulse Inhibition of Startle Reflex. Acta Paediatrica, 101, e304-e308.
- Blumenthal, T.D., Cuthbert, B.N., Filion, D.L., Hackley, S., Lipp, O.V. and van Boxtel, A. (2005) Committee Report: Guidelines for Human Startle Eyeblinkelectromyographic Studies. Psychophysiology, 42, 1-15. http://dx.doi.org/10.1111/j.1469-8986.2005.00271.x
- Gebhardt, J., Schulz-Juergensen, S. and Eggert, P. (2012) Maturation of Prepulse Inhibition (PPI) in Childhood. Psychophysiology, 49, 484-488. http://dx.doi.org/10.1111/j.1469-8986.2011.01323.x
- Knafo, A., Israel, S., Darvasi, A., Bachner-Melman, R., Uzefovsky, F., Cohen, L., Feldman, E., Lerer, E., Laiba, E., Raz, Y., Nemanov, L., Gritsenko, I., Dina, C., Agam, G., Dean, B., Bornstein, G. and Ebstein, R.P. (2008) Individual Differences in Allocation of Funds in the Dictator Game Associated with Length of the Arginine Vasopressin 1a Receptor RS3 Promoter Region and Correlation between RS3 Length and Hippocampal mRNA. Genes, Brain and Behavior, 7, 266-275. http://dx.doi.org/10.1111/j.1601-183X.2007.00341.x
- Frary, L.G. (1935) Enuresis: A Genetic Study. American Journal of Diseases in Children, 49, 557-578. http://dx.doi.org/10.1001/archpedi.1935.01970030003001
- Hallgren, B. (1957) Enuresis: A Clinical and Genetic Study. Acta Psychiatrica et Neurologica Scandinavica, Supplementum, 114, 1-159.
- Eiberg, H., Berendt, I. and Mohr, J. (1995) Assignment of Dominant Inherited Nocturnal Enuresis (ENUR1) to Chromosome 13q. Nature Genetics, 10, 354-356. http://dx.doi.org/10.1038/ng0795-354
- Arnell, H., Hjälmås, K., Jägervall, M., Läckgren, G., Stenberg, A., Bengtsson, B., Wassén, C., Emahazion, T., Annerén, G., Pettersson, U., Sundvall, M. and Dahl, N. (1997) The Genetics of Primary Nocturnal Enuresis: Inheritance and Suggestion of a Second Major Gene on Chromosome 12q. Journal of Medical Genetics, 34, 360-365. http://dx.doi.org/10.1136/jmg.34.5.360
- Eiberg, H. (1998) Total Genome Scan Analysis in a Single Extended Family for Primary Nocturnal Enuresis: Evidence for a New Locus (ENUR3) for Primary Nocturnal Enuresis on Chromosome 22q11. European Urology, 33, 34-36.
- Thibonnier, M., Auzan, C., Madhun, Z., Wilkins, P., Berti-Mattera, L. and Clauser, E. (1994) Molecular Cloning, Sequencing, and Functional Expression of a cDNA Encoding the Human V1a Vasopressin Receptor. The Journal of Biological Chemistry, 269, 3304-3310.
- Jonat, S., Santer, R., Schneppenheim, R., Obser, T. and Eggert, P. (1999) Effect of DDAVP on Nocturnal Enuresis in a Patient with Nephrogenic Diabetes Insipidus. Archives of Disease in Childhood, 81, 57-59. http://dx.doi.org/10.1136/adc.81.1.57
- Robben, J.H., Sze, M., Knoers, N.V., Eggert, P., Deen, P. and Müller, D. (2007) Relief of Nocturnal Enuresis by Desmopressin Is Kidney and Vasopressin Type 2 Receptor Independent. Journal of the American Society of Nephrology, 18, 1534-1539.
- Meir, J. and Eggert, P. (2011) Prepulse Inhibition of the Startle Reflex for the Differentiation of Enuresis in Children. Pediatric Nephrology, 26, 939-943.
Abbreviations
AVPR1A: arginine vasopressin receptor 1a
dDAVP: desamino arginine vasopressin
MNE: monosymptomatic nocturnal enuresis
NMNE: non-monosymptomatic nocturnal enuresis
NE: nocturnal enuresis
PPI: prepulse inhibition
RS: repeat sequence
NOTES

*Corresponding author.