Natural Resources
Vol.06 No.02(2015), Article ID:54257,9 pages
10.4236/nr.2015.62012
Growth Characteristic of the Oleaginous Microalga Chlorella ellipsoidea SD-0701 with Lipid Accumulation
Wenyu Luo1, Wenya Du1, Yi Su1, Jiejie Hui1, Jing Zhuang1, Lili Liu1,2*
1College of Life Science, Tianjin Normal University, Tianjin, China
2Tianjin Key Laboratory of Animal and Plant Resistance, Tianjin, China
Email: *liulilisd@126.com
Copyright © 2015 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 4 February 2015; accepted 23 February 2015; published 26 February 2015
ABSTRACT
Microalgae have great advantages as a new biomass source for fuel production. But microalgae are photosynthetic microorganisms, which normally grow in the light. Because of this growth condition, the commercial viability of microalgal biofuel is limited by current production systems. To obtain microalgal biofuel, fermentation is a more convenient, more economical and practical industry model. In this study, we asked whether and why the dark fermentation of C. ellipsoidea SD-0701 could be achieved by changing the culture medium formula. We focused the research on carbon-containing compounds and the initial pH of media. The results indicated that glucose was the optimum carbon-containing compound, which provided C. ellipsoidea SD-0701 with energy and carbon skeleton for accumulating organic compounds including lipids. When C. ellipsoidea SD-0701 was cultivated in the add-nutrition medium containing glucose, the optimum initial pH for the growth of C. ellipsoidea SD-0701 was pH 7.71. Therefore, if the suitable medium is used, C. ellipsoidea SD-0701 can grow normally in the dark, which is the same condition as the fermenter, and high microalgal biomass (0.50 g∙L−1) and lipid yield (232.90 mg∙L−1) can be achieved.
Keywords:
Oleaginous Microalga, Chlorella ellipsoidea, Growth Characteristic, Lipid Accumulation

1. Introduction
The development of renewable and eco-friendly sources of energy as alternative energy is urgently needed. Bioenergy is an alternative, carbon neutral, renewable source of energy produced from biomass [1] . The most common biofuels are usually produced from oleaginous crops and crop stover, such as rapeseed [2] -[7] , soybean [8] -[12] , sunflower [13] -[17] , corn [18] -[21] and palm [22] -[24] , through transesterification of vegetable oils with short chain alcohols [25] ; Ethanol was produced from crop stover via microorganism fermentation. The production of biofuels not only causes food shortages but also is of high production cost.
Microalgae have recently gained a lot of attention as a new biomass source for biofuel production [26] -[28] . Many strains of microalgae, such as Phaeodactylum tricornutum [29] -[32] , Haematococcus pluvialis [33] -[36] , Mychonastes afer HSO-3-1 [37] , Chlorella vulgaris [38] -[41] and Dunaliella tertiolecta [42] -[45] , have been reported as potential new sources of biodiesel. Unlike soybean and crops such as corn used for biofuels, most of algae do not need fresh water or agricultural land, and therefore do not compete with food supplies [46] . Algae grow incredibly fast, doubling their numbers every few hours, can be harvested daily, and have the potential to produce a volume of biomass many times greater than the most productive land crops [47] . The lipid content of algae is far greater than that of the vegetable oil crops, and under certain conditions some algal species can accumulate up to 70% of their dry weight as oil [48] . In addition, the efficiency of these photosynthetic algae in converting carbon dioxide into carbon-rich lipids, which is only a step or two away from biodiesel, greatly exceeds that of agricultural oleaginous crops. The high oil content together with rapid biomass production makes algae an excellent source for biodiesel production [49] .
Chlorella ellipsoidea is a single-celled, oil-rich alga. It is not only an oleaginous microalga but also a nutritious food with high protein content [50] [51] . In the production of biofuel with C. ellipsoidea, after the oil is extracted, the residue can be used as feed, leaving no industrial waste, so C. ellipsoidea is a high quality biomass for producing algal oil.
Before production of microalgal-derived diesel-fuel substitutes can become a large-scale commercial reality, the potential of microalgal biofuels need to be discussed, and the technical and economic barriers need to be overcome [52] . C. ellipsoidea is a photosynthetic microorganism, which normally grows in the light. The commercial viability of algal biofuel is limited by current production systems. Typical open ponds are inexpensive, but they often fail to maintain selected species for more than a few weeks or a few months due to contamination by invading organisms, including both competitors and predators [53] . The current industrial fermentation technology is very mature. Microorganisms can grow quickly in fermenters, and the production efficiency is greater than that of light conditions. Other advantages of fermentation include small operation space and high degree of automation. However, fermentative production is the process that the microorganisms are cultured in the dark and in a closed fermenter to produce a large amount of microbial products. In this study, we investigated whether and why changing the medium composition can achieve the dark fermentation of C. ellipsoidea. We focused the research on carbon-containing compounds and initial pH in the media. C. ellipsoidea was cultivated in dark and closed conditions. By adjusting carbon-containing compounds and initial pH of the medium, the high algae biomass and lipid content were achieved. We analyzed the effects of these factors on the cell metabolism of C. ellipsoidea, and provided scientific basis for fermenting oleaginous microalgae by traditional industrial dark fermentation to produce microalgae biofuels.
2. Materials and Methods
2.1. Oleaginous Microalga
Chlorella ellipsoidea SD-0701: Chlorella ellipsoidea SD-0701 was screened from YAOSHI Lake and maintained in the microbiology laboratory of Tianjin Normal University.
2.2. Growth Medium
SE medium: 0.25 g∙L−1 NaNO3, 0.075 g∙L−1 K2HPO4∙3H2O, 0.075 g∙L−1 MgSO4∙7H2O, 0.025 g∙L−1 CaCl2∙2H2O, 0.175 g∙L−1 KH2PO4, 0.025 g∙L−1 NaCl, 0.005 g∙L−1 FeCl3∙6H2O, 40 ml soil solution, 1 ml Fe-EDTA, 1 ml A5 solution, 958 ml ddH2O [54] . The medium was sterilized by autoclave at 121˚C for 20 min.
2.3. Determination of Microalgal Biomass
1.23 × 109 microalgal cells were inoculated in each 250-ml flask containing 100 ml medium. All the experiments were kept under continuous illumination at a light intensity of 60 - 80 μmol∙m−2∙s−1 at 25˚C for 7 days, and were shaken once every 4 hours (Illuminated incubator, LRH-250Z, Guangdong Province Medical Devices Factory). The algal cells were then harvested from the culture and centrifuged at 5000 rmin−1 for 10 min at 4˚C to produce a cellular pellet. The cellular pellet was dried in incubator at 80˚C until constant weight was reached, and the microalgal biomass was quantified.
2.4. Determination of the Lipid Content in the Microalgal Cells
To make a standard curve, total lipids were extracted from 10 g microalgae powder by acid hydrolysis method [55] . Lipids of 0.0, 0.1, 0.2, 0.5, 1.0, 2.0 and 5.0 mg were weighed and they were dissolved in 1 ml chloroform, respectively. 1 ml of H2SO4 was added into each chloroform solution, mixed thoroughly and incubated for 10 min at 100˚C. The solutions were cooled, and 5 ml of vanillin-phosphoric acid reagent (0.1978 g vanillin, 20 ml distilled water, 80 ml 85% phosphoric acid) was added into the solutions, respectively. The tubes were then incubated for 2 h at room temperature. The OD528 of each chloroform solution was read at 528 nm using a spectrophotometer [56] . The standard curve was plotted with the weight of the lipid on the x axis and the OD528 readings on the y axis, and linear regression analysis was performed on the standard curve.
The algal cells were harvested from 100 ml culture and centrifuged at 5000 rmin−1 for 10 min at 4˚C. Cellular deposit was frozen at −80˚C (Ultra-low Freezer, UF 3410, Heto-Holten) and thawed for 5 times in 24 h, and dried at 80˚C until constant weight was reached to produce dry microalgae powder. To determine the lipid content, 1 mg of dry microalgae powder was weighed and was treated according to the vanillin colorimetric method. Then lipid yields were calculated. The lipid content is the lipid weight per mg of dry microalgae powder. The lipid yield is the lipid weight per liter of the cultured-alga media.
2.5. The Comparison Method for Microalgae Growth and Lipid Accumulation in Media Supplemented with Different Carbon-Containing Compounds
The Basic Medium (BM) was SE medium. The effects of different carbon-containing compounds on microalgae growth were tested by adding them into the BM. The compounds were Glucose 25 mmol∙L−1, Citric Acid 25 mmol∙L−1, D-sorbitol 25 mmol∙L−1, Tris-acetic acid 25 mmol∙L−1, NaHCO3 20 mmol∙L−1, Na2CO3 20 mmol∙L−1 [57] . The control group was SE medium. The microalgal biomasses and lipid contents were measured according to the above methods, the lipid yields of cultures were calculated according to the lipid contents. Each experiment and the subsequent analysis were repeated three times.
2.6. The Comparison Method for Microalgae Growth and Lipid Accumulation in Media with Different Initial pH
The add-nutrition media (glucose 3 g∙L−1, equal to 25 mmol∙L−1, yeast extract 6 g∙L−1, KH2PO4 0.025 g∙L−1, Ca (NO3)2 0.1 g∙L−1, MgSO4∙7H2O 0.025 g∙L−1, KCl 0.012 g∙L−1, FeCl3 0.001 g∙L−1) were prepared. The pH of the media was buffered to different levels: 4.71, 5.71, 6.71, 7.71, 8.71 and 9.71. Microalgae were cultivated in Erlenmeyer flasks to the exponential phase of growth, and then 1.23 × 109 cells were transferred to 250-ml flasks containing 100 ml add-nutrition medium with different initial pH, static cultured for 7 days in a light intensity of 60 - 80 μmol∙m−2∙s−1, and shaken once every 4 hours. After cultivation, the microalgal biomasses and lipid contents were measured, the lipid yields of cultures were calculated according to the lipid contents. Each experiment and the subsequent analysis were repeated three times.
2.7. The Comparison Method for Microalgae Growth and Lipid Accumulation in Dark or Light Incubators
1.23 × 109 microalgal cells were inoculated into 250-ml flasks containing 100 ml add-nutrition medium confirmed by us, and cultured in different incubators at 25˚C. 1) Microalgae were cultured in the dark incubator for dark cultivation. 2) Microalgae were cultured in transparent incubator which was placed under sunlight. The experiment was done during seven consecutive sunny days. 3) Microalgae were cultured in illuminated incubator with daylight fluorescent tubes (T54-765, Philips) at 60 - 80 μmol∙m−2∙s−1. They were incubated for 7 days and shake once every 4 hours. The microalgal biomasses and lipid contents were measured according to the above methods, the lipid yields of cultures were calculated according to the lipid contents. Each experiment and the subsequent analysis were repeated three times.
2.8. Determination of Residual Carbohydrates in Microalgal Cultures
The residual carbohydrates in the microalgal cultures were determined by the anthrone method [58] . A particular operation is shown below.
Making the standard curve, the glucose was dried at 80˚C until constant weight was reached. Serial glucose standard solutions were prepared with concentrations of 0, 10, 20, 30, 40, 50, 60, 70, 80, 90 and 100 µg∙ml−1 in water. Add 5 ml of anthrone-H2SO4 reagent (0.1 mg anthrone, 70 ml H2SO4, 30 ml ddH2O) into the tubes containing 1 ml of glucose standard solution. The tubes were then incubated for 10 min at 100˚C. The solutions were cooled, and their absorbance was read at 625 nm using a spectrophotometer. The standard curve was plotted with the concentrations of glucose on x axis and the OD625 readings on y axis.
After the microalgae in media supplemented with nutrients were incubated, the cultures were centrifuged. 1 ml of the supernatant was transferred to a tube and mixed with 5 ml of anthrone-H2SO4 reagent. The tubes were then incubated for 10 min at 100˚C. The extracts were cooled, and their absorbance was read at 625 nm using a spectrophotometer. The content of residual carbohydrates in cultured-alga medium was then quantified according to the standard curve.
2.9. Statistical Analysis
All the experiments were conducted in three replicates. The data were analyzed using one-way analysis of variance (ANOVA) with SPSS software (version 18.0). A confidence of 95% (probability limit of p < 0.05) was chosen.
3. Results
3.1. Effects on C. ellipsoidea SD-0701 Growth and Lipid Accumulation in Media Supplemented with Different Carbon-Containing Compounds
After the C. ellipsoidea SD-0701 cells were cultivated in media supplemented with different carbon-containing compounds for 7 days, the cell concentrations of cultured-alga media in flasks have great differences (Figure 1(a)). Then the microalgal biomasses (Figure 1(a)) and lipid contents (Figure 1(b)) were determined and the lipid yields (Figure 1(c)) of the cultured-alga media were calculated. The highest microalgal biomass (1.15 g∙L−1) and lipid content (63.88%) were achieved in medium supplemented glucose and the lipid yield (734.4 mg∙L−1) of this group was also the highest (Figure 1). This result indicated that carbon-containing compounds had significant impact on the biomass and lipid content of C. ellipsoidea SD-0701. After the carbon-containing compounds were supplemented into SE media, algae biomass and lipid content have been significantly improved. And among the tested carbon-containing compounds, organic carbon-containing compounds were likely better than inorganic carbon-containing compounds. Among the tested organic carbon-containing compounds, glucose was the optimum, followed by Tris-acetic acid and D-sorbitol. C. ellipsoidea SD-0701 did not grow in medium containing citric acid.
We reformed the medium formula according to the above experimental results. A add-nutrition medium containing glucose was formulated and used in the subsequent experiments.
 (a) (b) (c)
(a) (b) (c)
Figure 1. Algal biomasses (a), lipid contents (b) and lipid yields (c) of C. ellipsoidea SD-0701 cultured in media supplemented with different carbon-containing compounds (p < 0.05, n = 3). CK, control check; S, sorbierite; G, glucose; CA, citric acid; TA, Tris-acetate; NH, NaHCO3; NA, Na2CO3.
3.2. Effects on C. ellipsoidea SD-0701 Growth and Lipid Accumulation in Add-Nutrition Media with Different Initial pH Levels
C. ellipsoidea SD-0701 grew in the add-nutrition media with six different initial pH levels. The effects on the growth and lipid accumulation of C. ellipsoidea SD0701 were researched (Figure 2). The microalgal biomass were about 0.34 g∙L−1 (initial pH 4.71), 0.40 g∙L−1 (initial pH 5.71), 0.64 g∙L−1 (initial pH 6.71), 0.95 g∙L−1 (initial pH 7.71), 0.77 g∙L−1 (initial pH 8.71) and 0.56 g∙L−1 (initial pH 9.71), respectively. The highest microalgal biomass (0.95 g∙L−1) was achieved in initial pH 7.71 medium. But the lipid content of this group was slightly lower than that of pH 8.71. The lipid yield were about 0.126 g∙L−1 (initial pH 4.71), 0.172 g∙L−1 (initial pH 5.71), 0.318 g∙L−1 (initial pH 6.71), 0.554 g∙L−1 (initial pH 7.71), 0.479 g∙L−1 (initial pH 8.71) and 0.294 g∙L−1 (initial pH 9.71), respectively (Figure 2(c)). The highest lipid yield (0.277 g/L) was also achieved in initial pH 7.71. So the optimum initial pH for C. ellipsoidea SD0701 growth and lipid accumulation was about 7.71.
3.3. Growth and Lipid Accumulation of C. ellipsoidea SD-0701 in Dark or Light Incubators
C. ellipsoidea SD-0701 were cultured with the add-nutrition medium at initial pH 7.71 in different incubators (Figure 3). The effects on microalgal biomasses, lipid contents and lipid yields of C. ellipsoidea SD-0701 in different incubators were investigated and the residual carbohydrate contents in the cultured-algae media, were also determined. The microalgal biomasses were 1.00 g∙L−1 (dark), 0.98 g∙L−1 (sunlight) and 0.96 g∙L−1 (fluorescent light), respectively (Figure 4(a)). The lipid yields were 467.9 mg∙L−1 (dark), 388.3 mg∙L−1 (sunlight) and 541.2 mg∙L−1 (fluorescent light) (Figure 4(c)). The results indicated that there was no significant difference in microalgal biomass and lipid accumulation when C. ellipsoidea SD-0701 was cultured in the dark or light incubators. The residual carbohydrates content in the cultured medium was the highest under sunlight and the lowest was in the dark (Figure 4(d)).
 (a) (b) (c)
(a) (b) (c)
Figure 2. Algal biomasses (a), lipid contents (b) and lipid yields (c) of C. ellipsoidea SD-0701 cultivated in media at different initial pH levels (p < 0.05, n = 3).
 (a) (b) (c)
(a) (b) (c)
Figure 3. C. ellipsoidea SD-0701 cultured in dark incubator (a), transparent incubator placed in sunlight (b) and illuminated incubator with fluorescent light (c), respectively.
 (a) (b)
(a) (b) Light source Light source(b) (c)
Light source Light source(b) (c)
Figure 4. Algal biomasses (a), lipid contents (b) and lipid yields (c) of C. ellipsoidea SD-0701 in dark or light incubators and the residual carbohydrates contents of cultured-alga media after cultivation (d) (p < 0.05, n = 3).
4. Discussion
Different carbon-containing compounds have different transport mechanisms and metabolic pathways. And they also provide different amounts of energy for cell growth. Glucose is transported into C. ellipsoidea SD-0701 cells by facilitated enzyme, and its transport rate is relatively high. After glucose in medium enters C. ellipsoidea l SD-0701 cells, it is phosphorylated by a family of enzymes called hexokinases to form glucose 6-phos- phate (G6P). One G6P is then oxidized completely through the TCA cycle, in the process approximate 32 additional molecules of ATP produced. In addition, its decomposition products provide carbon skeleton for constructive metabolism and lipid accumulation. So the microalgal biomass (1.15 g∙L−1) was the highest when glucose was supplemented for the growth of C. ellipsoidea l SD-0701.
Tris-acetic acid is transported by active transport. One molecule of Tris-acetic acid is catalyzed to acetyl-CoA by acetyl-CoA synthase after it enters the cell. Acetyl-CoA is oxidized completely through the TCA cycle and produces 9 molecules of ATP for C. ellipsoidea SD-0701. Therefore, glucose produces more ATP and is more beneficial for C. ellipsoidea SD-0701 growth than Tris-acetic acid. Sorbitol is transported into C. ellipsoidea SD-0701 cells by active transport, which needs to consume ATP. After entering the cell, sorbitol oxidase catalyzes the conversion of sorbitol to glucose which then enters TCA cycle after glycolysis. Sorbitol oxidase is the rate-limiting enzyme, whose level in the cell limits the metabolic rate of sorbitol. So the metabolic rate of C. ellipsoidea SD-0701 in medium with sorbitol is much lower than that with glucose. This has also been verified in the results of microalgal biomass. The microalgal biomass was 0.31 g∙L−1 when sorbitol was used as carbon- containing compound, and it was 1.15 g∙L−1 when glucose was used as carbon-containing compound.
The result indicated that C. ellipsoidea SD-0701 did not grow in medium with citric acid. The reasons might be: 1) The activity of citrate synthase was feedback inhibited by citric acid, which could affect TCA circle; 2) Cells could be blocked in G1 phase by citric acid, which could block DNA synthesis and inhibit cell growth [59] .
Cheng et al. [60] has reported that C. ellipsoidea can only fix two forms of dissolved inorganic carbon―CO2 and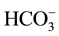 . And only when
. And only when 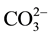 is converted into CO2 and
is converted into CO2 and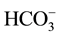 , can they be fixed and utilized by C. ellipsoidea SD-0701. However, the pH of the medium increases with the increase of CO2-consuming, which is not favor condition for the conversion from
, can they be fixed and utilized by C. ellipsoidea SD-0701. However, the pH of the medium increases with the increase of CO2-consuming, which is not favor condition for the conversion from 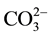 to CO2 and
to CO2 and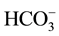 . Therefore, C. ellipsoidea SD-0701 grow faster in medium supplemented with NaHCO3 than with Na2CO3. NaHCO3 and Na2CO3 provide inorganic carbon-containing compounds for C. ellipsoidea SD-0701. They could not provide both energy and decomposition products for cells, lower microalgal biomass was achieved.
. Therefore, C. ellipsoidea SD-0701 grow faster in medium supplemented with NaHCO3 than with Na2CO3. NaHCO3 and Na2CO3 provide inorganic carbon-containing compounds for C. ellipsoidea SD-0701. They could not provide both energy and decomposition products for cells, lower microalgal biomass was achieved.
NaHCO3 and Na2CO3 provide CO2 for C. ellipsoidea SD-0701 when they are used as carbon-containing com- pounds. C. ellipsoidea SD-0701 cannot utilize CO2 for photosynthesis in the dark. Therefore, it is not suitable to use NaHCO3 and Na2CO3 as carbon-containing compounds in traditional dark fermentation.
A add-nutrition medium that can effectively promote the growth and lipid accumulation of C. ellipsoidea SD-0701 was formulated according to above experimental results. And the pH level of the medium was researched. Because the initial pH level of medium affects the available fraction of nutrients in media, the membrane stability and absorptive capacity of different nutrient elements. The highest microalgal biomass (0.95 g∙L−1) was achieved in initial pH 7.71. Its lipid content was slightly lower than that of pH 8.71. But the highest lipid yield was also achieved in pH 7.71. So pH 7.71 is optimal initial pH level of medium.
The residual carbohydrate contents in the cultured-alga media in different light conditions were markedly different. The residual carbohydrate content with dark treatment was the lowest (Figure 4(d)), and the microalgal biomass of this group was the highest (Figure 4(a)). It suggests that photoreaction of C. ellipsoidea SD-0701 was stopped and dark metabolism was activated under the dark treatment condition. In dark C. ellipsoidea SD-0701 absorbed nutrients directly, glucose was mainly utilized to provide energy and carbon skeleton for synthesizing organic compound, and to finally achieve high microalgal biomass and lipid accumulation. Therefore, the existing fermentation equipment can be applied to the industrial production of C. ellipsoidea SD-0701 oil, like the traditional production of microbial fermentation products.
5. Conclusion
In conclusion, after the SE medium was supplemented with suitable concentration of glucose, the microalgal biomass and lipid yield have been increased 4.11 and 7.70 times, respectively. Adding different carbon-con- taining compounds into the media, the growth rate of C. ellipsoidea SD-0701 was changed. The highest microalgal biomass and lipid content were achieved when glucose was applied in the medium and the lipid yield of this group was also the highest. The optimum pH of the add-nutrition medium was pH 7.71. Although C. ellipsoidea SD-0701 is a photoautotrophic microorganism, microalgal biomass and lipid yield can be enhanced in a dark incubator being filled with the add-nutrition medium. It is feasible that the modern fermentation equipment can be applied to the industrial production of C. ellipsoidea SD-0701 oil, just like the traditional production of microbial fermentation products.
Acknowledgements
This work was supported in part by the Natural Science Foundation of China under grant (#31172029), Tianjin Natural Science Foundation under grant (#07JCYBJC15400), and a grant (#0802220) from Tianjin Agriculture and Rural Affairs Committee, as well as a grant (#12ZCZDNC00600) from Tianjin Municipal Science and Technology Program.
References
- Tushar, K.G. and Mark, A.P. (2011) Bioenergy. In: Energy Resources and Systems, 327-418.
- Herrmann, I.T., Jorgensen, A. and Bruun, S. (2012) Potential for Optimized Production and Use of Rapeseed Biodiesel. International Journal of Life Cycle Assessment, 18, 418-430. http://dx.doi.org/10.1007/s11367-012-0486-8
- Tutunea, D. (2012) Thermal Investigation of Biodiesel Blends Derived from Rapeseed Oil. Journal of Thermal Analysis and Calorimetry, 111, 869-875. http://dx.doi.org/10.1007/s10973-012-2213-x
- Firrisa, M.T., Iris, V.D. and Voinov, A. (2013) Energy Efficiency for Rapeseed Biodiesel Production in Different Farming Systems. Energy Efficiency, 7, 79-95. http://dx.doi.org/10.1007/s12053-013-9201-2
- Kamzolova, S.V., Dedyukhina, E.G. and Samoilenko, V.A. (2013) Isocitric Acid Production from Rapeseed Oil by Yarrowia lipolytica Yeast. Applied Microbiology and Biotechnology, 97, 9133-9144. http://dx.doi.org/10.1007/s00253-013-5182-5
- Kreivaitis, R., Gumbyte, M., Kazancev, K., Padgurskas, J. and Makarevitiene, V. (2013) A Comparison of Pure and Natural Antioxidant Modified Rapeseed Oil Storage Properties. Industrial Crops and Products, 43, 511-516. http://dx.doi.org/10.1016/j.indcrop.2012.07.071
- Qi, D.H., Bae, C. and Feng, Y.M. (2013) Preparation, Characterization, Engine Combustion and Emission Characteristics of Rapeseed Oil Based Hybrid Fuels. Renew Energy, 60, 98-106. http://dx.doi.org/10.1016/j.renene.2013.05.008
- Pessoa, F.L.P., Magalhaes, S.P. and Pedro, W.C.F. (2010) Production of Biodiesel via Enzymatic Ethanolysis of the Sunflower and Soybean Oils: Modeling. Applied Biochemistry and Biotechnology, 161, 238-244. http://dx.doi.org/10.1007/s12010-009-8878-3
- Simasatitkul, L.P., Siricharnsakunchai, Patcharavorachot, Y., Assabumrungrat, S. and Arpornwichanop, A. (2011) Re- active Distillation for Biodiesel Production from Soybean Oil. Korean Journal of Chemical Engineering, 28, 649-655. http://dx.doi.org/10.1007/s11814-010-0440-z
- Go, A.-R., Lee, Y., Kim, Y.H., et al. (2013) Enzymatic Coproduction of Biodiesel and Glycerol Carbonate from Soybean Oil in Solvent-Free System. Enzyme and Microbial Technology, 53, 154-158. http://dx.doi.org/10.1016/j.enzmictec.2013.02.016
- Oliveira, S.O., Maruyama, S.A. and Claus, T. (2013) A Novel Response Surface Methodology Optimization of Base- Catalyzed Soybean Oil Methanolysis. Fuel, 113, 580-585. http://dx.doi.org/10.1016/j.fuel.2013.06.011
- Wan, L., Liu, H. and Skala, D. (2014) Biodiesel Production from Soybean Oil in Subcritical Methanol Using MnCO3/ ZnO as Catalyst. Applied Catalysis B: Environmental, 152, 352-359. http://dx.doi.org/10.1016/j.apcatb.2014.01.033
- Santos, A.G.D., Caldeira, V.P.S., Farias, M.F., Araujo, A.S., Souza, L.D. and Barros, A.K. (2011) Characterization and Kinetic Study of Sunflower Oil and Biodiesel. Journal of Thermal Analysis and Calorimetry, 106, 747-751. http://dx.doi.org/10.1007/s10973-011-1838-5
- Tavares, M.L.A., Queiroz, N., Santos, I.M.G., Souza, A.L., Cavalcani, E.H.S., Barros, A.K.D., Rosenhaim, R., Sole- dade, L.E.B. and Souza, A.G. (2011) Sunflower Biodiesel Use of P-DSC in the Evaluation of Antioxidant Efficiency. Journal of Thermal Analysis and Calorimetry, 106, 575-579. http://dx.doi.org/10.1007/s10973-011-1357-4
- Alenezi, R., Santos, R.C.D., Raymahasay, S. and Leeke, G.A. (2013) Improved Biodiesel Manufacture at Low Temperature and Short Reaction Time. Renewable Energy, 53, 242-248. http://dx.doi.org/10.1016/j.renene.2012.11.019
- Sanchez, B.S., Mendow, G. and Levrand, P.G. (2013) Optimization of Biodiesel Production Process Using Sunflower Oil and Tetramethyl Ammonium Hydroxide as Catalyst. Fuel, 113, 323-330. http://dx.doi.org/10.1016/j.fuel.2013.05.085
- Spinelli, D., Jez, S. and Pogni, R. (2013) Environmental and Life Cycle Analysis of a Biodiesel Production Line from Sunflower in the Province of Siena (Italy). Energy Policy, 59, 492-506. http://dx.doi.org/10.1016/j.enpol.2013.04.009
- Min, J.Y. and Lee, E.Y. (2011) Lipase-Catalyzed Simultaneous Biosynthesis of Biodiesel and Glycerol Carbonate from Corn Oil in Dimethyl Carbonate. Biotechnology Letters, 33, 1789-1796. http://dx.doi.org/10.1007/s10529-011-0627-3
- Raul, D., Jose, G.R. and Natalia, G. (2013) Energy Valorisation of Crude Glycerol and Corn Straw by Means of Slow Co-Pyrolysis: Production and Characterisation of Gas, Char and Bio-Oil. Fuel, 112, 31-37. http://dx.doi.org/10.1016/j.fuel.2013.05.005
- Shi, A.M., Du, Z.Y. and Ma, X.C. (2013) Production and Evaluation of Biodiesel and Bioethanol from High Oil Corn Using Three Processing Routes. Bioresource Technology, 128, 100-106. http://dx.doi.org/10.1016/j.biortech.2012.10.007
- Ciftci, O.N. and Temelli, F. (2014) Continuous Bioconversion of the Lipids of Corn, Wheat, and Triticale Distiller’s Dried Grains with Solubles to Biodiesel in Supercritical Carbon Dioxide and Characterization of the Products. BioEnergy Research, 7, 702-710. http://dx.doi.org/10.1007/s12155-013-9394-4
- Silalertruksa, T. and Gheewala, S.H. (2012) Environmental Sustainability Assessment of Palm Biodiesel Production in Thailand. Energy, 43, 306-314. http://dx.doi.org/10.1016/j.energy.2012.04.025
- Cho, H.J., Kim, J.K. and Ahmed, F. (2013) Life-Cycle Greenhouse Gas Emissions and Energy Balances of a Biodiesel Production from Palm Fatty Acid Distillate (PFAD). Applied Energy, 111, 479-488. http://dx.doi.org/10.1016/j.apenergy.2013.05.038
- Liaquat, A.M., Masjuki, H.H. and Kalam, M.A. (2013) Impact of Palm Biodiesel Blend on Injector Deposit Formation. Applied Energy, 111, 882-893. http://dx.doi.org/10.1016/j.apenergy.2013.06.036
- Khan, S.A., Rashmi, Hussain, M.Z., Prasad, S. and Banerjee, U.C. (2009) Prospects of Biodiesel Production from Microalgae in India. Renewable and Sustainable Energy Reviews, 13, 2361-2372. http://dx.doi.org/10.1016/j.rser.2009.04.005
- Satyanarayana, K.G., Mariano, A.B. and Vargas, J.V.C. (2011) A Review on Microalgae, a Versatile Source for Sustainable Energy and Materials. International Journal of Energy Research, 35, 291-311. http://dx.doi.org/10.1002/er.1695
- Nwachukwu, A.N. and Chukwu, M.A. (2012) The Potential of Macroalgae as a Source of Carbohydrates for Use in Bioethanol Fermentation. International Journal of Energy and Environmental, 3, 761-774.
- Wu, Y.H., Hu, H.Y., Yu, Y., Zhang, T.Y., Zhu, S.F., Zhuang, L.L., Zhang, X. and Lu, Y. (2014) Microalgal Species for Sustainable Biomass/Lipid Production Using Wastewater as Resource: A Review. Renewable and Sustainable Energy Reviews, 33, 675-688. http://dx.doi.org/10.1016/j.rser.2014.02.026
- Gong, Y. and Jiang, M. (2011) Biodiesel Production with Microalgae as Feedstock: From Strains to Biodiesel. Biotechnology Letters, 33, 1269-1284. http://dx.doi.org/10.1007/s10529-011-0574-z
- Burrows, E.H., Bennette, N.B., Carrieri, D., Dixon, J.L., Brinker, A., Frada, M., Baldassano, S.N., Falkowski, P.G. and Dismukes, G.C. (2012) Dynamics of Lipid Biosynthesis and Redistribution in the Marine Diatom Phaeodactylum tricornutum under Nitrate Deprivation. Biotechnology Letters, 5, 876-885. http://dx.doi.org/10.1007/s12155-012-9201-7
- Gardner, R.D., Cooksey, K.E., Mus, F., Macur, R., Moll, K., Eustance, E., et al. (2012) Use of Sodium Bicarbonate to Stimulate Triacylglycerol Accumulation in the Chlorophyte Scenedesmus sp. and the Diatom Phaeodactylum tricornutum. Journal of Applied Phycology, 24, 1311-1320. http://dx.doi.org/10.1007/s10811-011-9782-0
- Fields, M.W., Hise, A., Lohman, E.J., Bell, T., Gardner, R.D., Corredor, L., et al. (2014) Sources and Resources: Im- portance of Nutrients, Resource Allocation, and Ecology in Microalgal Cultivation for Lipid Accumulation. Journal of Applied Phycology, 98, 4805-4816. http://dx.doi.org/10.1007/s00253-014-5694-7
- Luis, F.R. (2011) Net Energy Calculations for Production of Biodiesel and Biogas from Haematococcus pluvialis and Nannochloropsis sp. Green Energy and Technology, 83-91.
- Lei, A., Chen, H., Shen, G.M., Hu, Z.L., Chen, L. and Wang, J.X. (2012) Expression of Fatty Acid Synthesis Genes and Fatty Acid Accumulation in Haematococcus pluvialis under Different Stressors. Biotechnology for Biofuels, 5, 18. http://dx.doi.org/10.1186/1754-6834-5-18
- Saha, S.K., McHugh, E. and Hayes, J. (2012) Effect of Various Stress-Regulatory Factors on Biomass and Lipid Production in Microalga Haematococcus pluvialis. Bioresource Technology, 128, 118-124. http://dx.doi.org/10.1016/j.biortech.2012.10.049
- Gerhard, K. (2013) Production and Properties of Biodiesel from Microalgal Oils. Developments in Applied Phycology, 5, 207-221.
- Yuan, C., Liu, J.H., Fan, Y., Ren, X.H., Hu, G.R. and Li, F.L. (2011) Mychonastes afer HSO-3-1 as a Potential New Source of Biodiesel. Biotechnology for Biofuels, 4, 47. http://dx.doi.org/10.1186/1754-6834-4-47
- Cheirsilp, B., Kitcha, S. and Torpee, S. (2011) Co-Culture of an Oleaginous Yeast Rhodotorula glutinis and a Microalga Chlorella vulgaris for Biomass and Lipid Production Using Pure and Crude Glycerol as a Sole Carbon Source. Annals of Microbiology, 62, 987-993. http://dx.doi.org/10.1007/s13213-011-0338-y
- Pribyl, P., Cepak, V. and Zachleder, V. (2012) Production of Lipids in 10 Strains of Chlorella and Parachlorella, and Enhanced Lipid Productivity in Chlorella vulgaris. Applied Microbiology and Biotechnology, 94, 549-561. http://dx.doi.org/10.1007/s00253-012-3915-5
- Talebi, A.F., Thidfar, M. and Tabatabaei, M. (2013) Genetic Manipulation, a Feasible Tool to Enhance Unique Characteristic of Chlorella vulgaris as a Feedstock for Biodiesel Production. Molecular Biology Reports, 40, 4421-4428. http://dx.doi.org/10.1007/s11033-013-2532-4
- Al-Lwayzy, S.H., Yusaf, T. and Al-Juboori, R.A. (2014) Biofuels from the Fresh Water Microalgae Chlorella vulgaris (FWM-CV) for Diesel Engines. Energies, 7, 1829-1851. http://dx.doi.org/10.3390/en7031829
- Demirbas, A. and Demirbas, M.F. (2010) Green Energy and Technology, Algae Energy, Algae as a New Source of Biodiesel. Springer, London, 139-157.
- Tang, H., Abunasser, N. and Garcia, M.E.D. (2011) Potential of Microalgae Oil from Dunaliella tertiolecta as a Feed- stock for Biodiesel. Applied Energy, 88, 3324-3330. http://dx.doi.org/10.1016/j.apenergy.2010.09.013
- Moheimani, N.R. (2012) Long-Term Outdoor Growth and Lipid Productivity of Tetraselmis suecica, Dunaliella tertiolecta and Chlorella sp (Chlorophyta) in Bag photobioreactors. Journal of Applied Phycology, 25, 167-176. http://dx.doi.org/10.1007/s10811-012-9850-0
- Chow, Y.S., Goh, S.J.M., Su, Z.H., Ng, D.H.P., Lima, C.Y., Lim, N.Y.N., et al. (2013) Continual Production of Gly- cerol from Carbon Dioxide by Dunaliella tertiolecta. Bioresource Technology, 136, 550-555. http://dx.doi.org/10.1016/j.biortech.2013.03.040
- Mario, R. and Tredici (2010) Photobiology of Microalgae Mass Cultures: Understanding the Tools for the Next Green Revolution. BioFuels, 1, 143-162.
- Dismukes, G.C., Carrieri, D., Bennette, N., Ananyev, G.M. and Posewitz, M.C. (2008) Aquatic Phototrophs: Efficient Alternatives to Land-Based Crops for Biofuels. Current Opinion in Biotechnology, 19, 235-240. http://dx.doi.org/10.1016/j.copbio.2008.05.007
- Perego, C. and Ricci, M. (2012) Diesel Fuel from Biomass. Catalysis Science & Technology, 2, 1776-1786. http://dx.doi.org/10.1039/c2cy20326j
- Demirbas, A. (2010) Biodiesel from Oilgae, Biofixation of Carbon Dioxide by Microalgae: A Solution to Pollution Problems. Applied Energy, 88, 3541-3547. http://dx.doi.org/10.1016/j.apenergy.2010.12.050
- Liang, S.Z., Liu, X.M., Chen, F. and Chen, Z.J. (2004) Current Microalgal Health Food R & D Activities in China. Hydrobiologia, 512, 45-48. http://dx.doi.org/10.1023/B:HYDR.0000020366.65760.98
- Pauline, S., Claire, J.C., Elie, D. and Arsène, I. (2006) Commercial Applications of Microalgae. Journal of Bioscience and Bioengineering, 101, 87-96. http://dx.doi.org/10.1263/jbb.101.87
- Pienkos, P.T. and Darzins, A.L. (2009) The Promise and Challenges of Microalgal-Derived Biofuels. Biofuels, Bio- products and Biorefining, 3, 431-440. http://dx.doi.org/10.1002/bbb.159
- Rodolfi, L., Chini Zittelli, G., Bassi, N., Padovani, G., Biondi, N., Bonini, G. and Tredici, M.R. (2008) Microalgae for Oil: Strain Selection, in Duction of Lipid Synthesis and Outdoor Mass Cultivation in a Low-Cost Photobioreactor. Biotechnology and Bioengineering, 102, 100-112. http://dx.doi.org/10.1002/bit.22033
- Zhang, C.M., Pan, W.B. and Chen, Y.Z. (2009) The Toxic Effects of the Extracellular Active Components from One Algae-Lysing Bacteria on Chlorella pyrenoidosa. Microbiology, 36, 821-825.
- Ma, S., Fu, L.L., Wang, M., Hu, X.W., Tan, D.G. and Zhang, J.M. (2010) Comparison of Extraction Methods for Crude Fat from Microalgae. China Oils and Fats, 35, 77-79.
- Gunnel, A. and Leonardo, M. (1991) Lipid Analysis of Freshwater Microalgae: A Method Study. Archiv Fur Hydrobiologie, 121, 295-306.
- Zheng, H.L., Gao, Z., Zhang, Q. and Huang, H. (2011) Effect of Inorganic Carbon Source on Lipid Production Withautotrophic Chlorella vulgaris. Chinese Journal of Biotechnology, 27, 436-444.
- Xue, Y.L. (1985) Determination of Soluble Sugar in the Plant Material. In: Xue, Y.L., Ed., Lab Manual of Plant Phy- siology, 134-138.
- Huang, J.F. and Zhang, Y.X. (2006) Influence of Citrate on Growth and Metabolism of CHO Cells. Journal of East China University of Science and Technology (Natural Science Edition), 32, 556-560.
- Cheng, L.H., Zhang, L., Chen, H.L. and Gao, C.K. (2005) Advances on CO2 Fixation by Microalgae. Chinese Journal of Biotechnology, 1, 177-181.
NOTES

*Corresponding author.