Open Journal of Marine Science
Vol.4 No.3(2014), Article
ID:48418,9
pages
DOI:10.4236/ojms.2014.43018
Cryptic Behaviour of Juvenile Turbot Psetta maxima L. and European Flounder Platichthys flesus L.
Louise Dahl Kristensen1*, Claus Reedtz Sparrevohn1, Jens Tang Christensen2, Josianne Gatt Støttrup1
1Technical University of Denmark, National Institute of Aquatic Resources, Charlottenlund, Denmark
2Aarhus University, Department of Bioscience, Aquatic Biology, Aarhus, Denmark
Email: *LKR@aqua.dtu.dk
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 9 April 2014; revised 21 May 2014; accepted 18 June 2014
ABSTRACT
The aim of this study was to examine the burying behaviour of hatchery-reared European flounder Platichthys flesus and turbot Psetta maxima, and whether conditioning on a sandy substrate would improve burying efficiency. Both species buried shortly after release on a sandy substrate. However, the study revealed interspecies differences; the flounder buried immediately after release, while the turbot buried gradually. No significant difference in burying efficiency was observed between naïve and conditioned flounder and turbot. An effect of size on burial efficiency was observed for both flounder and turbot with a tendency for larger fish to bury more efficiently than smaller fish, despite previous conditioning. Size at settlement was found to be >2 cm for flounder and >3 cm for turbot.
Keywords:Juvenile Flatfish, Burying Behaviour, Conditioning, Stock Enhancement

1. Introduction
Post-release mortality is a major concern in stock enhancement since it often determines the success of stocking hatchery-reared flat fish [1] . Reared individuals lack behavioural skills and are thus more vulnerable to predators than their wild counterparts. Flatfish are reared in tanks with no sand on the bottom and have no experience of natural predators. Thus, they have a slower escape response [2] and are less efficient at burying into the sediment [3] . A shadow in the rearing tanks generally means food and causes the fish to leave the bottom to feed (personal observation, JG Støttrup), while in nature this behaviour would render the fish vulnerable to e.g. avian predation. Because pelleted feed is provided ad libitum in the rearing systems, they are inexperienced both in finding, capturing and eating live prey. Furthermore, the flatfish are starved to reduce metabolism during transportation and reduce mortality from handling stress during release. These factors all contribute to a higher mortality rate just after release [4] and a link between post-release mortality and increased vulnerability to predators have been demonstrated in laboratory studies [5] .
Predation was the major cause of post-release mortality in Japanese flounder Paralichthys olivaceus (Temminck & Schlegel) with daily mortality rates of 5% - 30% within the first five days of release. Subsequently, mortality dropped to 1% - 5% per day; the primary predators in this case were piscivorous fish [6] (in [7] ). Daily mortality of 14% was observed for released turbot Psetta maxima L. in the first few days after release, primarily caused by the great black-backed gull Larus marinus and herring gull Larus argentatus [4] . Temporary periods of low nutritional intake and unsuccessful adaptation to the natural environment also may increase post-release mortality [8] [9] , although the impact on mortality was not quantified in these studies. Therefore, conditioning hatchery-reared flatfish to the wild environment may be beneficial for the survival of the flatfish and thus for the cost-benefit of stock enhancement programmes [1] and [10] .
Adapting hatchery-reared flatfish to the natural environment with plenty of stimuli (e.g. live prey, larger habitat, sandy sediment, etc.) while protected from predators is called “conditioning”. This conditioning process increased survival by up to 50% for turbot [11] . There may be several reasons for this increased survival: First, flatfish not adapted to the natural environment (naïve) spend significantly more time off the bottom feeding than conditioned flatfish and since most predation probably happens off the bottom, this behaviour has the potential to make the flatfish more vulnerable to predators [12] [13] . Second, conditioning may improve the swimming performance of the flatfish and improve their skills to escape predators. However, since conditioning periods of a few days duration have been successful in increasing survival, this seems unlikely [13] . Third, cryptic behaviour improves when flatfish are conditioned even for short periods, both in terms of colour change in order to resemble the sediment and in terms of burying skill [3] .
Juvenile flounder Platichthys flesus L. and turbot make their appearance in nursery areas at a size (standard length, SL) between ~10 and 20 mm for flounder and ~20 and 30 mm for turbot (J. Martinsson & A. Nissling, unpubl. data in [14] ). In several flatfish species, metamorphosis is size-related but for others it depends on growth rate [15] [16] or temperature [17] . After complete metamorphosis the flatfish settle in nursery grounds and commence the benthic life stage [18] . The period shortly after settlement is crucial for many flatfish species, with high mortality due to predation by shrimp, crab, gadoids, and other piscivorous fish [19] -[21] . Thus, growth to obtain size-refuge from predators and the ability to bury efficiently are important factors in determining survival in the early juvenile stages. The burying ability of juvenile flatfish is strongly correlated with body size; Japanese flounder and plaice Pleuronectes platessa increase burying ability with size [22] [23] . A positive relationship between burying ability and sediment choice was demonstrated for Japanese flounder, European flounder, Pacific halibut Hippoglossus stenolepis (Schmith) and northern rock sole Lepidopsetta polyxystra (Orr and Matarese) [22] [24] [25] . Burying ability and efficiency have not previously been examined in European flounder and turbot. As these are two important species in marine fish releases in Denmark, there is a need to examine their behaviour.
The aim of this study was to examine burying behaviour in different sized juvenile European flounder and turbot. Furthermore we examined if conditioning fish to a sandy substrate could increase burying efficiency and cryptic behaviour in hatchery-reared fish.
2. Materials and Methods
Since burying behaviour of hatchery-reared winter flounder Pseudopleuronectes americanus (Walbaum) was observed to improve in efficiency after two days of conditioning [5] , we chose a three day conditioning period in this study.
Juvenile flounder and turbot were reared in fiberglass tanks at a commercial hatchery (Maximus A/S) in Denmark, and had thus never been exposed to sand (naïve). The fish were offspring of wild fish caught in the release area.
Two laboratory experiments were conducted comparing reared naïve and conditioned flounder (Exp A), and reared naïve and conditioned turbot (Exp B; Table 1).

Table 1. Overview of laboratory experiments.
N is the number of fish analysed.
2.1. European Flounder
All flounder were measured to the nearest lower mm and grouped into one cm size classes between 1 to 6 cm. Naïve flounder were kept in 20 L plastic tanks in conditions similar to rearing conditions, and densities of approximately 100 fish∙m−2. Sea water flow of 1 L∙min−1 was provided and the flounder were fed a diet of chopped mussels and commercial fish feed pellets during the daytime. Half the flounder were conditioned in the field in three aluminium bottomless cages (1.41 m × 1.41 m × 1.41 m) on a sandy substrate at a beach in Løgstør Broad, Limfjorden, Denmark. Conditioned flounder were held in these cages for three days at densities of approximately 100 fish∙m−2.
2.2. Turbot
All turbot were measured to the nearest lower mm and grouped into one cm size classes between 1 to 7 cm. Na- ïve turbot were kept under similar conditions as the naïve flounder. The turbot were conditioned for three days in 20 L plastic tanks at densities of approximately 100 fish∙m−2 with sufficient sand covering the bottom (3 cm) to allow burial behaviour.
2.3. Laboratory Experiments
The experiments were performed in round plastic tanks (h: 13 cm, d: 14.5 cm). Sediment was added to each tank covering the bottom by enough sand to allow burial (3 cm). The sediment was collected locally from an area, where both flounder and turbot naturally occur. The sediment was not sieved in order to make the test facility resemble a natural release site, although stones were removed. All tanks had an individual seawater flow of approximately 100 ml∙min−1. The water column in each container was 8 cm high with a volume of approximately 2 L.
A total of 62 flounder and 120 turbot were tested with 8 - 11 and 10 - 12 fish in each size group, respectively. Overhead strip lights and windows provided light for a photoperiod of 16:8 L:D. Temperature and oxygen levels were measured daily with an OxyGuard Handy Delta and salinity level was measured with a handheld refractometer.
One naïve or conditioned flounder or turbot (Exp. A1, A2, B1 or B2, Table 1) was gently released into a tank. Each fish was observed directly from above and the percentage of the fish’s dorsal surface covered by sediment was evaluated into one of four categories: 1 = not buried, 2 = less than 50% buried, 3 = more than 50% buried or 4 = completely buried. If the fish swam in the water column or adhered to the sides of the tanks this was noted. The behaviour was observed every five min during the first half hour and subsequently once every 30 min for a total of two hours. The tests were performed in daytime during the summer of 2007 and all fish were tested only once.
2.4. Statistical Analysis
To test whether there was any effect of size on burying score (efficiency), the Kruskal-Wallis One Way Analysis of Variance on Ranks (KW) was used for the naïve and conditioned flounder and turbot.
The Mann-Whitney Rank Sum Test (MW) was used to test if there was an effect of the conditioning period of three days for the durations of 5 and 120 min, and for the total experimental period. The effect of time on the burying frequency of the flounder and turbot was tested using KW. All fish burying to any degree, scoring 2 or higher was included in these tests. All tests were run in SigmaStat (Version 3.5, 2006) and significance levels were set at 5%.
3. Results
3.1. Abiotic Measurements
The temperature during the tests varied from 16.5˚C to 19.2˚C, with temperatures slightly higher in midsummer analysis than in the tests performed in late summer. The overall mean temperature was 17.6˚C. The mean oxygen content of the seawater was 7.3 mg O2 l−1 with measurements ranging from 5.6 to 8.0 mg O2 l−1. The salinity of the seawater was between 25 and 26 throughout the study with a mean salinity of 25.9.
3.2. Burying Behaviour
Significant interspecies differences between the average burying time for flounder and turbot were found, with flounder burying more rapidly than turbot (Figure 1). This difference was especially pronounced for the smaller (size 2.0 - 2.9 and 3.0 - 3.9 cm), with flounder burying within 5 - 15 min while turbot buried after approximately 60 min. For fish larger than 4.0 cm, again flounder buried more rapidly at approximately 5 min while the turbot buried at 30 min (Table 2).
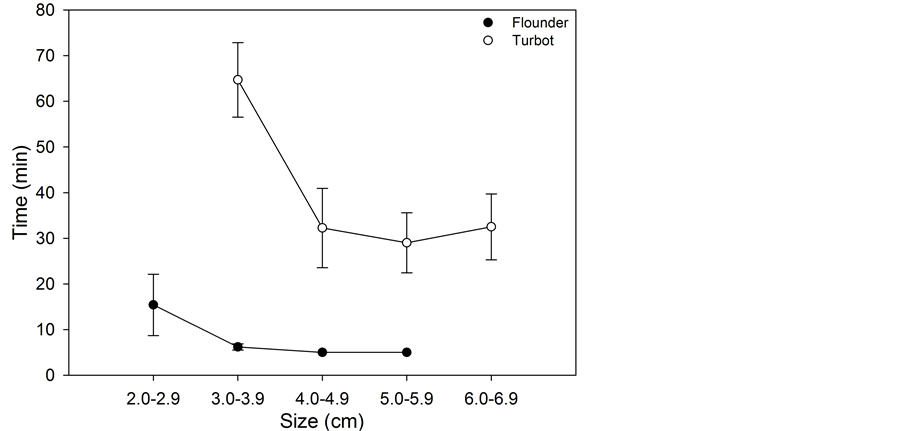
Figure 1. Average burying time for the different size classes in naïve and conditioned flounder (2 - 6 cm) and turbot (3 - 7 cm) with standard deviation.
Table 2. Percentage of fish buried (category 2 - 4, burial score: 1 = not buried, 2 = <50% buried, 3 = 50% buried, and 4 = 100% buried) at different time intervals (5 - 120 minutes) after release.
The frequency of buried flounder during the total experimental period divided into size classes is demonstrated in Figure 2(a). Except for the smallest group, all flounder buried quickly after release and the frequency of buried flounder generally increased with size. Time had no significant effect on the burial behaviour of flounder.
The number of burying turbot also increased with size (Figure 2(b)). However, in contrast to flounder, only a few of the turbot had buried at the beginning of the experiment. Time had a significant effect on burial behaviour in turbot (KW, df = 8, P < 0.001).
3.3. Settlement
For naïve 1.0 - 1.9 cm flounder, 63% of the fish never settled on the sediment while fish larger than 2.0 cm all settled for most of the observed period (Figure 2(a)). A similar pattern was observed for turbot (Figure 2(b)) in which 76 % of the naïve 1.0 - 2.9 cm fish never settled, but swam in the water column or adhered to sides of the tank during the observations.
3.4. Burying Efficiency
The average burying scores for the first 5 min of the experiment (Figure 3(a)) and for the total observation period of 120 min for the laboratory experiments (Figure 3(b)) increased with increasing fish size for both naïve and conditioned flounder (KW, df = 2, P < 0.01 and df = 3, P < 0.05). The case was the same for naïve and conditioned turbot (KW, df = 4, P < 0.001 and df = 4, P < 0.005).
Conditioned flounder and turbot did not score higher or, in other words, they did not bury more efficiently than naïve flounder and turbot (Figure 3). Except for turbot 4 - 5 cm (MW, 120 min: P < 0.05, total period: P = 0.005), none of the conditioned fish buried more efficiently than naïve fish (see P-values in Table 3).
4. Discussion
4.1. Is Burial Behaviour Innate?
The evolution of flatfishes has been a gradual event [26] , and it is believed that their unusual morphology is an adaptation to avoid predation and ambush prey [27] . Burying to reduce predation risk has been demonstrated experimentally in sole [3] . Further, more summer flounder Paralichthys dentatus have been shown to bury in the presence of predators [28] .
Winter flounder did not bury immediately, concluding that for this species, burying is a learned response and not innate behaviour [3] . In contrast naïve flounder and turbot buried immediately in this study; a result similar to those of other studies [3] [29] who found that cultured turbot buried immediately and 99% of the reared sole buried in sand within one minute after release, respectively. In this study, no differences in burying behaviour between naïve and conditioned flounder and turbot were observed. Furthermore, the flounder and turbot used in
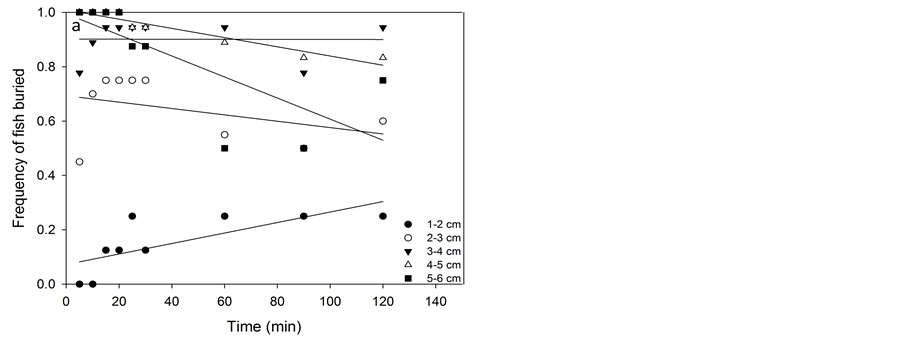

Figure 2. Frequency of naïve and conditioned fish (combined) buried over time. Flounder (a) and turbot (b) from Exp. A1, A2, B1 and B2. Linear regressions demonstrate trends in data.
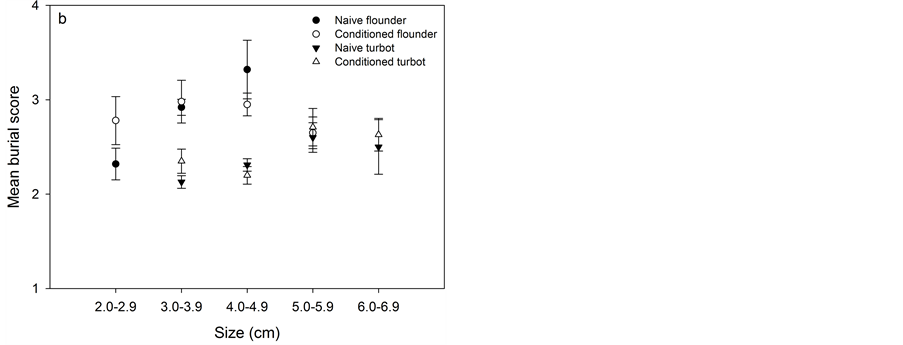

Figure 3. Mean burial score for different sized naïve and conditioned flounder (2 - 6 cm) and turbot (3 - 7 cm) from Exp. A1, A2, B1 and B2 at 5 min (a) and at end of the experiment after 120 min (b) with standard deviation. Burial score: 1 = not buried, 2 = <50% buried, 3 = 50% buried, and 4 = 100% buried.
Table 3. Statistical results of Mann-Whitney Rank Sum Test testing naïve vs conditioned flounder and turbot of the same size groups at 5 and 120 minutes after release. *P < 0.05, **P < 0.01.
the present study showed clear signs of burying; e.g. alternated beating of the head and tail against the bottom of the rearing tanks (pers. obs). The same behaviour has been described as typical burying behaviour in Japanese flounder [30] . Along with the results of sole [3] , this suggests that while the tendency for certain flatfish to bury is innate, this may not apply to all flatfish as in the case of winter flounder [5] .
4.2. Size and Cryptic Behaviour
Of the 1.0 - 1.9 cm naïve flounder, 63% never settled on the sediment during the observations, but adhered to the sides of the tank or swam in the water column with a tilted body posture. This suggests that flounder settle when they are larger than 2 cm and eye migration is completed. For naïve turbot less than 3 cm, 76% never settled on the sediment. Hence, the turbot are larger than flounder when they settle. This is consistent with the fact that turbot are larger than flounder at metamorphosis, 16 mm and 8 - 11 mm, respectively [31] [32] .
This study demonstrated a difference between burying behaviour in different sized fish; both the frequency and efficiency of burying increased with size. A positive relationship between the size of the flatfish and its ability to conceal itself has been demonstrated in other studies [22] [23] , and larger fish bury more efficiently in larger grain sizes than smaller fish did. It is assumed that larger fish bury more efficiently than smaller fish due to the greater force they can exert when concealing themselves [23] , suggesting that larger fish can inhabit more potential habitats than smaller fish [22] .
4.3. Differences between Conditioned and Naïve Fish
Conditioning fish before releasing into the wild is important for stocking success. The burial efficiency of similar sized hatchery-reared fish increased to the same level as that of wild fish after a conditioning period of 12 days for sole [3] , and 2 days for winter flounder [5] . As we did not find any difference in burying efficiency between naïve and conditioned flounder and turbot in this study, we suggest that turbot and flounder may need longer conditioning periods than winter flounder. Several studies have demonstrated lower mortality rates [10] [33] with as much as a 50% decrease in mortality [11] . The improved survival in those studies may not be due solely to improved burying efficiency, but also to changes in feeding behaviour and thus lower vulnerability to predators [12] [34] . Further investigations are needed to clarify the specific causes for improved post-release survival rates in hatchery-reared fishes, but there is sufficient evidence to warrant the need for longer conditioning periods for turbot and flounder before release.
4.4. Interspecies Differences
The turbot did not bury as quickly as the flounder in this experiment suggesting an interspecies difference in burying behaviour. While 77% of all settled flounder buried to some degree within the first five min after release, only 16% of all settled turbot had buried within this time. Fish tend to demonstrate fright response caused by transfer stress when released into a new environment [35] , so the observed behaviour may have been a “stress response” even though precautions were made to stress the fish as little as possible before and during the release. The stress response of flounder is to bury rapidly while the stress response of turbot is to lie motionless on the sediment before eventually burying, and these behaviours have been observed previously by the authors.
Another possible explanation for the observed differences in burial efficiency is differences in diel feeding behaviour between flounder and turbot. However, there seems to be some disagreement on the diel activity of flounder and turbot in the literature (flounder nocturnal: [36] -[40] , turbot nocturnal: [39] [41] , flounder diurnal: [42] , turbot diurnal: [42] [43] ). These disagreements may originate from the fact that the flatfish change feeding habits depending on size, area, and available prey [40] [44] -[46] . The present experiments were carried out during daytime and if it is assumed that the flounder is nocturnal and the turbot is diurnal, this could cause the flounder to stay buried during daytime while the turbot would be more likely to stay on the sediment ready to pursue prey. Repeating this experiment at night using infrared light or examining diel feeding behaviour in flatfish could help clarify this question.
Acknowledgements
The work was supported by Etatsråd C.G. Filtenborg and wife Marie Filtenborgs Scholarship and the Danish National Coastal Fisheries Management Program. The authors wish to thank Hanne Nicolajsen and Ole Madvig Larsen for assistance in the field.
References
- Sproul, J.T. and Tominaga, O. (1992) An Economic Review of the Japanese Flounder Stock Enhancement Project in Ishikari Bay, Hokkaido. Bulletin of Marine Science, 50, 75-88.
- Gibson, S. and Johnston, I.A. (1995) Scaling Relationships, Individual Variation and the Influence of Temperature on Maximum Swimming Speed in Early Settled Stages of the Turbot Scopthalmus maximus. Marine Biology, 121, 401-408. http://dx.doi.org/10.1007/BF00349449
- Ellis, T., Howell, B.R. and Hughes, R.N. (1997) The Cryptic Responses of Hatchery-Reared Sole to a Natural Sand Substratum. Journal of Fish Biology, 51, 389-401. http://dx.doi.org/10.1111/j.1095-8649.1997.tb01674.x
- Sparrevohn, C.R., Nielsen, A. and Stottrup, J.G. (2002) Diffusion of Fish from a Single Release Point. Canadian Journal of Fisheries and Aquatic Sciences, 59, 844-853. http://dx.doi.org/10.1139/f02-059
- Fairchild, E.A. and Howell, W.H. (2004) Factors Affecting the Post-Release Survival of Cultured Juvenile Pseudopleuronectes americanus. Journal of Fish Biology, 65, 69-87.
- Furuta, S., Watanabe, T. and Yamada, H. (1998) Predation by Fishes on Hatchery-Reared Japanese Flounder Paralichthys olivaceus Juveniles Released in the Coastal Area of Tottori Prefecture. Nippon Suisan Gakkaishi, 64, 1-7. (in Japanese) http://dx.doi.org/10.2331/suisan.64.1
- Yamashita, Y. and Yamada, H. (1999) Release Strategy for Japanese Flounder Fry in Stock Enhancement Programmes. In: Howell, B.R., Moksness, E. and Svasand, T., Eds., Stock Enhancement and Sea Ranching, Fishing News Books, Oxford, 191-204.
- Macquart-Moulin, C., Champalbert, G., Howell, B.R., Patriti, G. and Ranaivoson, C. (1991) La relation alimentation-fixation benthique chez les jeunes sole Soleasolea L. métamorphosées. Evidences expérimentales. Journal of Experimental of Marine Biology and Ecology, 153, 195-205. (in French)http://dx.doi.org/10.1016/0022-0981(91)90225-L
- Howell, B.R. and Yamashita, Y. (2005) Aquaculture and Stock Enhancement. In: Gibson, R.N., Ed., Flatfishes, Biology and Exploitation, Blackwell Publishing, Oxford, 416.
- Howell, B.R. (1994) Fitness of Hatchery-Reared Fish for Survival in the Sea. Aquaculture and Fisheries Management, 25, 3-17.
- Sparrevohn, C.R. and Stottrup, J.G. (2007) Post-Release Survival and Feeding in Reared Turbot. Journal of Sea Research, 57, 151-161. http://dx.doi.org/10.1016/j.seares.2006.08.010
- Furuta, S. (1996) Predation on Juvenile Japanese Flounder (Paralichthys olivaceus) by Diurnal Piscivorous Fish: Field Observations and Laboratory Experiments. In: Watanabe, Y., Yamashita, Y. and Oozeki, Y., Eds., Survival Strategies in Early Life Stages of Marine Resources, A. A. Balkema, Rotterdam, 285-294.
- Tsukamoto, K., Masuda, R., Kuwada, H. and Uchida, K. (1997) Quality of Fish for Release: Behavioural Approach. Bulletin of the National Research Institute of Aquaculture, 3, 93-99.
- Nissling, A., Jacobssen, M. and Hallberg, N. (2007) Feeding Ecology of Juvenile Turbot Scophthalmus maximus and Flounder Pleuronectes flesus at Gottland, Central Baltic Sea. Journal of Fish Biology, 70, 1877-1897. http://dx.doi.org/10.1111/j.1095-8649.2007.01463.x
- Gibson, R.N. (2005) The Behaviour of Flatfishes. In: Gibson, R.N., Ed., Flatfishes Biology and Exploitation, Blackwell Publishing, Oxford, 213-239.
- Geffen, A.J., van der Veer, H.W. and Nash, R.D.M. (2007) The Cost of Metamorphosis in Flatfish. Journal of Sea Research, 58, 35-45. http://dx.doi.org/10.1016/j.seares.2007.02.004
- Benoit, H.P. and Pepin, P. (1999) Individual Variability in Growth Rate and the Timing of Metamorphosis in Yellowtail Flounder Pleuronectes ferrugineus. Marine Ecology Progress Series, 184, 231-244. http://dx.doi.org/10.3354/meps184231
- Gibson, R.N. (1997) Behaviour and the Distribution of Flatfishes. Journal of Sea Research, 37, 241-256. http://dx.doi.org/10.1016/S1385-1101(97)00019-1
- van der Veer, H.W. and Bergman, M.J.N. (1987) Predation by Crustaceans on a Newly Settled 0-Group Plaice Pleuronectes platessa Population in the Western Wadden Sea. Marine Ecology Progress Series, 35, 203-215. http://dx.doi.org/10.3354/meps035203
- Ellis, T. and Gibson, R.N. (1995) Size Selective Predation of 0-Group Flatfishes on Scottish Coastal Nursery Ground. Marine Ecology Progress Series, 127, 27-37. http://dx.doi.org/10.3354/meps127027
- Fairchild, E.A. and Howell, W.H. (2000) Predator-Prey Size Relationship between Pseudopleuronectes americanus and Carcinus maenas. Journal of Sea Research, 44, 81-90. http://dx.doi.org/10.1016/S1385-1101(00)00037-X
- Tanda, M. (1990) Studies on Burying Ability in Sand and Selection to the Grain Size for Hatchery-Reared Marbled Sole and Japanese Flounder. Nippon Suisan Gakkaishi, 56, 1543-1548. http://dx.doi.org/10.2331/suisan.56.1543
- Gibson, R.N. and Robb, L. (1992) The Relationship between Body Size, Sediment Grain Size and the Burying Ability of Juvenile Plaice, Pleuronectes platessa L. Journal of Fish Biology, 40, 771-178. http://dx.doi.org/10.1111/j.1095-8649.1992.tb02623.x
- Gibson, R.N. and Robb, L. (2000) Sediment Selection in Juvenile Plaice and its Behavioural Basis. Journal of Fish Biology, 56, 1258-1275. http://dx.doi.org/10.1111/j.1095-8649.2000.tb02138.x
- Stoner, A.W. and Ottmar, M.L. (2003) Relationship between Size-Specific Sediment Preferences and Burial Capabilities in Juveniles of Two Alaska Flatfishes. Journal of Fish Biology, 282, 85-101.
- Friedman, M. (2008) The Evolutionary Origin of Flatfish Asymmetry. Nature, 454, 209-212. http://dx.doi.org/10.1038/nature07108
- Bock, W. and von Wahlert, G. (1965) Adaptation and the Form-Function Complex. Evolution, 19, 20-69. http://dx.doi.org/10.2307/2406439
- Keefe, M.L. and Able, K.W. (1994) Contributions of Abiotic and Biotic Factors to Settlement in Summer Flounder, Paralichthys dentatus. Copeia, 1994, 458-465. http://dx.doi.org/10.2307/1446993
- Iglesias, J. and Rodríguez-Ojea, G. (1994) Fitness of Hatchery-Reared Turbot, Scophthalmus maximus L., for Survival in the Sea: First Year Results on Feeding, Growth and Distribution. Aquaculture and Fisheries Management, 25, 179-188.
- Kawabe, R., Nashimoto, K., Hiraishi, T., Naito, Y. and Sato, K. (2003) A New Device for Monitoring the Activity of Freely Swimming Flatfish, Japanese Flounder Paralichthys olivaceus. Fisheries Science, 63, 3-10. http://dx.doi.org/10.1046/j.1444-2906.2003.00581.x
- Osse, J.W.M. and Van den Boogart, J.G.M. (1997) Size of Flatfish Larvae at Transformation, Functional Demands and Historical Constraints. Journal of Sea Research, 37, 229-239. http://dx.doi.org/10.1016/S1385-1101(97)00025-7
- Munk, P. and Nielsen, J.G. (2005) Eggs and Larvae of North Sea Fishes. Biofolia, Frederiksberg.
- Hossain, M.A.R., Tanaka, M., and Masuda, R. (2002) Predator-Prey Interaction between Hatchery-Reared Japanese Flounder Juvenile, Paralichthys olivaceus, and Sandy Shore Crab, Matuta lunaris: Daily Rhythms, Anti-Predator Conditioning and Starvation. Journal of Experimental Marine Biology and Ecology, 267, 1-14. http://dx.doi.org/10.1016/S0022-0981(01)00340-9
- Howell, B.R. and Baynes, S.M. (1993) Are Hatchery-Reared Sole Equipped for Survival in the Sea? ICES CM 1993/F:33 SESS R.
- Henderson, H.F. (1980) Behavioural Adjustments of Fishes to Release into a New Habitat. In: Bardach, J.E., Magnuson, J.J., May, R.C. and Reinhart, J.M., Eds., Fish Behaviour and Its Use in the Capture and Culture of Fishes, ICLARM, Manila, 512.
- Bregnballe, F. (1962) Plaice and Flounder as Consumers of the Microscopic Bottom Fauna. Meddelelser fra Danmarks Fiskeriog Havundersogelser, 3, 133-182.
- Muus, B.J. (1967) The Fauna of Danish Estuaries and Lagoons. Distribution and Ecology of Dominating Species in the Shallow Reaches of the Mesohaline Zone. Meddelelser fra Danmarks Fiskeriog Havundersogelser, 5, 173-208.
- Verheijen, F.J. and De Groot, S.J. (1967) Diurnal Activity Pattern of Plaice and Flounder (Pleuronectidae) in Aquaria. Netherlands Journal of Sea Research, 3, 383-390. http://dx.doi.org/10.1016/0077-7579(67)90011-7
- Nash, R.D.M., Santos, R.S., Geffen, A.J., Hughes, G. and Ellis, T.R. (1994) Diel Variability in Catch Rate of Juvenile Flatfish on Two Small Nursery Grounds (Port Erin Bay, Isle of Man and Porto Pim Bay, Faial, Azores). Journal of Fish Biology, 44, 35-45.
- Miyazaki, T., Masuda, R., Furuta, S. and Tsukamoto, K. (1997) Laboratory Observations on the Nocturnal Activity of Hatchery-Reared Juvenile Japanese Flounder Paralichthys olivaceus. Fisheries Science, 62, 205-210.
- Champalbert, G. and Le Direach-Boursier, L. (1998) Influence of Light and Feeding Conditions on Swimming Activity Rhythms of Larval and Juvenile Turbot Scophthalmus maximus L.: An Experimental Study. Journal of Sea Research, 40, 333-345. http://dx.doi.org/10.1016/S1385-1101(98)00031-8
- Holmes, R.A. and Gibson, R.N. (1983) A Comparison of Predatory Behaviour in Flatfish. Animal Behaviour, 31, 1244-1255. http://dx.doi.org/10.1016/S0003-3472(83)80031-1
- Imsland, A.K., Folkvord, A. and Stefansson, S.O. (1995) Growth, Oxygen Consumption and Activity of Juvenile Turbot (Scophthalmus maximus L.) Reared under Different Temperatures and Photoperiods. Netherlands Journal of Sea Research, 34, 149-159. http://dx.doi.org/10.1016/0077-7579(95)90023-3
- Summers, R.W. (1979) The Diet and Feeding Behaviour of the Flounder Platichthys flesus (L.) in the Ythan Estuary, Aberdeenshire, Scotland. Estuarine and Coastal Marine Science, 11, 217-232. http://dx.doi.org/10.1016/S0302-3524(80)80042-9
- Bos, A.R. (1999) Aspects of the Life History of the European Flounder (Pleuronectes flesus L. 1758) in the Tidal River Elbe. Ph.D. Thesis, Hamburg University, Hamburg.
- Vinagre, C., Cabral, H. and Costa, M.J. (2008) Prey Selection by Flounder, Platichthys flesus, in the Douro Estuary, Portugal. Journal of Applied Ichthyology, 24, 238-243. http://dx.doi.org/10.1111/j.1439-0426.2008.01055.x
NOTES

*Corresponding author.