Open Journal of Immunology
Vol. 3 No. 1 (2013) , Article ID: 28675 , 9 pages DOI:10.4236/oji.2013.31004
In vitro activity and function of B7-H4-Ig fusion protein
![]()
Laboratory of Experimental Immunology, Department of International Health, Immunology and Microbiology, The Faculty of Health Sciences, University of Copenhagen, Copenhagen, Denmark; #Corresponding Author: Claesson@sund.ku.dk
Received 9 October 2012; revised 22 November 2012; accepted 2 December 2012
Keywords: CD28 Family; B7-H4; Fusion Protein; MLC
ABSTRACT
B7-H4 has been shown to inhibit T cell proliferation, cytokine production and cell cycle in vitro. B7-H4 deficient mice develop exacerbated disease in the mouse models of Rheumatoid Arthritis (RA), Type 1 Diabetes (T1D) and Experimental Autoimmune Encephalomyelitis (EAE). On the other hand, B7-H4-Ig fusion protein has been documented to assuage the symptoms in mouse models of RA, T1D, and multiple sclerosis in vivo. In the present study, B7-H4-Ig bound to the majority of human peripheral blood monocytes and NK cells, but not to either normal or activated T cells. B7-H4-Ig fusion protein was assayed for its effects in allogeneic mixed lymphocyte culture (MLC) systems. Soluble B7-H4- Ig had no significant effect in the MLC, but with a tendency to promote allogeneic response. Immobilized, but not soluble B7-H4-Ig inhibited plastic bound anti-CD3 mediated activation of T cells. This inhibition however was largely due to B7-H4-Ig mediated displacement of anti-CD3 antibody from the plastic plate. Finally, B7-H4-Ig had no effect on the cytotoxicity mediated by NK and LAK cells in PBMC. Our findings thus caution against the interpretation of suppressive effect observed solely in plate-bound anti-CD3 mediated T cell co-stimulation in vitro.
1. INTRODUCTION
The B7 family member B7-H4, discovered ten years ago, has been shown to inhibit TCR-mediated proliferation, cell-cycle progression and IL-2 production by CD4+ and CD8+ T cells in murine [1-3] and human [4]
in vitro systems. Regulatory T cells (Treg) have been shown to induce IL-10 secretion and upregulation of B7-H4 in APCs [5]. B7-H4 mRNA is broadly expressed in many tissue and cell types, including tumors [6]. B7-H4 protein , however, is limited to activated T cells, B cells, monocytes, dendritic cells [2] activated hepatic stellar cells, synovia cells in rheumatoid arthritis, islet beta cell, renal tubular and lung epithelial cells [7,8]. A number of cancers have been shown to over-express B7-H4 protein, including human ovarian, breast, prostate and lung cancer, renal-cell carcinoma and uterine adenocarcinoma [9]. In ovarian carcinoma, B7-H4 expressing tumor-associated macrophages inhibit Tumor Associated Antigen (TAA)-specific T-cell effector function [10]. B7-H4 blockade with a monoclonal antibody in the EAE mouse model led to exacerbated disease and to an accumulation of CD8 + T cells and macrophages in the brain [1]. In vivo administration of B7-H4Ig in a murine graft-versus-host disease model reduced both T cell proliferation, CTL activity and improved animal survival [2]. A soluble form of B7-H4 has been implicated in pathogenesis of rheumatoid arthritis and genetic deletion of B7-H4 accelerated the progression of collagen-induced arthritis in mice [11]. B7-H4 knockout mice suffer from a more severe experimental diabetes due to increased islet infiltration of Th1 and Th17 cells, while mice overexpressing B7-H4 were protected from diabetes by reducing IFN-production in CD4 T cells, without skewing towards Th2 phenotype or activation of Tregs [12]. Treating NOD mice with B7-H4-Ig resulted in reduced incidence of diabetes through a transient increase in Tregs number and suppressive activity [13]. The receptor for B7-H4 has yet to be identified, but is believed to be expressed on subsets of immune cells, including activated T cells and neutrophils. The B7-H4 receptor most probably belongs to an unknown member of the co-inhibitory CD28 superfamily that includes BTLA, CTLA-4 and PD-1 [14].
In the present paper, we studied a recently developed recombinant human B7-H4-Ig fusion protein, B7-H4-Ig (provided by AmplimmunInc.U.S.A.) in human in vitro systems. Binding of B7-H4-Ig to normal and activated human peripheral blood mononuclear cells (PBMC) was examined. Whereas the majority of monocytes and NK cells were shown to bind B7-H4-Ig, less than 10% of normal B cells and neither naive, nor activated T cells were found to bind B7-H4-Ig. In a series of allogeneic mixed lymphocyte culture (MLC) experiments, B7-H4-I g showed a slight, but insignificant enhancing effect on proliferation. An inhibitory effect of B7-H4-Ig on immobilized anti-CD3 induced T cell proliferation was observed, but proved to be caused by B7-H4-Ig mediated displacement of anti-CD3 antibody from the culture well. Finally, neither the activity of NK nor LAK cells was influenced by exposure to B7-H4-Ig.
2. MATERIALS AND METHODS
2.1. Antibodies and Control Reagents
Anti-CD3 antibody (F101.01) recognizes a conformational determinant on the TcR/CD3 complex [15]. The antibody was a kind gift from Professor C. Geisler, University of Copenhagen and was used for T cell activation. The following mAbs were purchased from Becton-Dicki nson,New Jersey, U.S.A. as mouse anti-human Abs: FITC-conjugated anti-CD3; APC-conjugated anti -CD8, anti-CD14, and anti-CD56. Mouse anti-human FITCconjugated anti-CD4 and APC-conjugated anti C-D19 an tibodies were purchased from Elektra and PE-con jugated mouse anti-human B7-H4 (clone H74) was purchased from eBioscience (California, U.S.A.). B7 -H4-Ig (human B7-H4 extracellular domain fused with human IgG1) fusion proteins were produced in suspension culture in an animal protein-free-adapted CHOK- 1SV (Lon za Biologics, Allendale, NJ) cell line utilizing the glutamine synthetase gene expression system, and purified using Amplimmune’s protein purification process. KS is an inactive mutant of B7-DC-Ig (provided by Amplimmune Inc.), a fusion protein consisting of the Fc portion of human IgG1 protein and extracellular domain of B7-DC, in which a missense mutation in the active site changed lysine residue at position 113 to serine. The KS mutant was used as a negative, isotype control for B7- H4-Ig. Synagis (also known as palivizumab) was used as another negative isotype control for B7-H4-Ig, as well as a Fc-blocker for flow cytometry experiments. Synagis is a humanized (IgG1 Fc domain) monoclonal antibody against respiratory syncytial virus.
2.2. Cell Cultures
Human PBMCs were obtained from buffy coats of healthy donors in Danish Blood Donor Corps by Ficoll gradient centrifugation and stored at −140˚C. Thawing of the cells followed a standard protocol. Cells were cultured on 96-well round (MLCs) or flat-bottomed plates (anti-CD3 stimulated T cells) in 220 μl of RPMI-1640 medium supplied with Glutamax, 10% human serum (Valley Biomedical), 0.5% 2-mercaptoethanol and 0.5% penicillin/streptomycin. All cultures were conducted with a total of 0.2 million cells per well in an incubator at 37˚C, 5% CO2. One-way MLCswere prepared by irradiating one set of cells (“stimulator”) with 2000 rad from Cs-137 for 10 minutes. CD8 T cells were depleted using Dynabeads-CD8 (Invitrogen) following manufacturer’s manual. T cells were enriched 2-3 fold by two-step nonadherence to plastic. All cells were cultured for 72 hours and their proliferation assessed by tritium thymidine incorporation assay.
Immobilization of B7-H4-Ig, KS, anti-CD3 antibody and Synagis or a mixture of them was carried out with 40 μl of the agent incubated in flat-bottomed plates overnight at 4˚C and washed twice. In case of experiments shown in Figures 5-8 the indicated agent (B7-H4-Ig, KS or Synagis) was either added as “immobilized” (incubated overnight together with anti-CD3) or “soluble” (added together with the cells after the plate had been washed). In experiments shown in Figures 9 and 10 the plate was washed and B7-H4-Ig was added at 40 μl for two hours at 37˚C followed by washing and plating of cells essentially as described by Sica et al. [2].
2.3. Flow Cytometry
Cells for FACS analysis were incubated with 300 μg/ml Synagis for 15 min on ice (to block Fc receptors) followed by incubation for 1h on ice with either 100 μg/ml of B7-H4-Ig or PBS. After wash 1 x 106 Cells were stained with fluorochrome labeled antibodies containing 1% bovine serum albumin (BSA) in PBS for 20 minutes on ice. After washing twice, cells were fixed with 1% paraformaldehyde. Isotype control antibodies were included to define appropriate cut-off levels. For labelling of non-adherent, anti-CD3 activated T cells, these were incubated for 15 minutes with 300 μg/ml Synagis, washed, and incubated for 1 hour with 200 μg/ml B7-H4-Ig or PBS, then washed and incubated for 30 minutes in FITC-conjugated mouse anti-CD3 (BD Pharmingen) and PE-conjugated mouse anti-B7-H4 antibodies (eBioscience,clone H74), in PBS supplied with 0,5% BSA. Cells were then washed, resuspended in either FACS buffer or 1% paraformaldehyde and analysed using a FACScalibur flow cytometer (Becton-Dickinson). Data were analysed using cellquest software (BectonDickinson).
2.4. Simultaneous ELISA and Proliferation Assays
Two plates were incubated with a range of anti-CD3 concentrations overnight at 4˚C. They were washed twice with PBS, incubated with 40 μl of 50 μg/ml of B7-H4-Ig or PBS for 2 hours at 37˚C and washed twice with PBS. ELISA assay for anti-CD3 binding was performed on the first plate. To the second plate enriched human T cells were added and incubated at 37˚C/5% CO2 for 3 days. Tritium thymidine incorporation was performed to assess the proliferation.
2.5. 51Cr-Release Assay
Freshly harvested PBMCs from healthy donors were washed twice and resuspended in medium containing 10% FCS, at 5 × 106 cells/ml and directly tested for cytotoxicity or incubated for 24 h min at 37˚C in the atmosphere of 5% CO2 in air in the presence of a stimulatingcytokine cocktailof (IL-12 and IL-15: 50 ng/ml, IL-2: 2000 U/ml and IFN-α: 10.000 U/ml) (ImmunoTools GmbH; Germany) to generate LAK cells. B7-H4 -Ig was added to the effector cells at 50µg/ml and the formulation buffer and PBS was added as controls.K562 and Daudi cells were used as targets for NK and LAK respectively. Target cells were labeled with 30μCi/mil of 51Cr (Perkin Elmer, Billerica, MA) for 1 h at 37˚C. The labeled cells were washed and resuspended in medium. Cells were co-incubated at effector to target (E:T) ratios at 100:1 - 12.5:1. Co-cultures were set up in 96 well V-bottom plates (Nunc; Denmark) in four replicates and incubated for 4h at 37˚C in the atmosphere of 5% CO2 in air. Controls included target cells incubated in medium alone for spontaneous release or lysed with 10% triton X-100 for maximal release. Radioactivity was measured by Wallac Wizard 1470 Automatic Gamma Counter. Specific release was calculated using the following formula:
% specific lysis = (sample cpm − spontaneous cpm)/ (maximal cpm − spontaneous cpm) × 100%.
2.6. Data Treatment and Statistics
Data were analysed in R, using ggplot2 package for visualization. Statistical significance was assessed by onesample t-test on medium control normalized data. A posthoc Benjamini-Hochberg correction was employed to control for multiple testing. The results of simultaneous ELISA and proliferation assays were analysed by fitting a four-point logistic model over a range of anti-CD3 concentrations and response values (c.p.m. and OD values in proliferation and ELISA assays, respectively) in control samples. Using this model anti-CD3 concentrations corresponding to response values in B7-H4-Ig treated samples were reversely predicted. Ratio of predicted and originally applied anti-CD3 antibody concentrations was calculated (equalling 1 for control samples) and plotted.
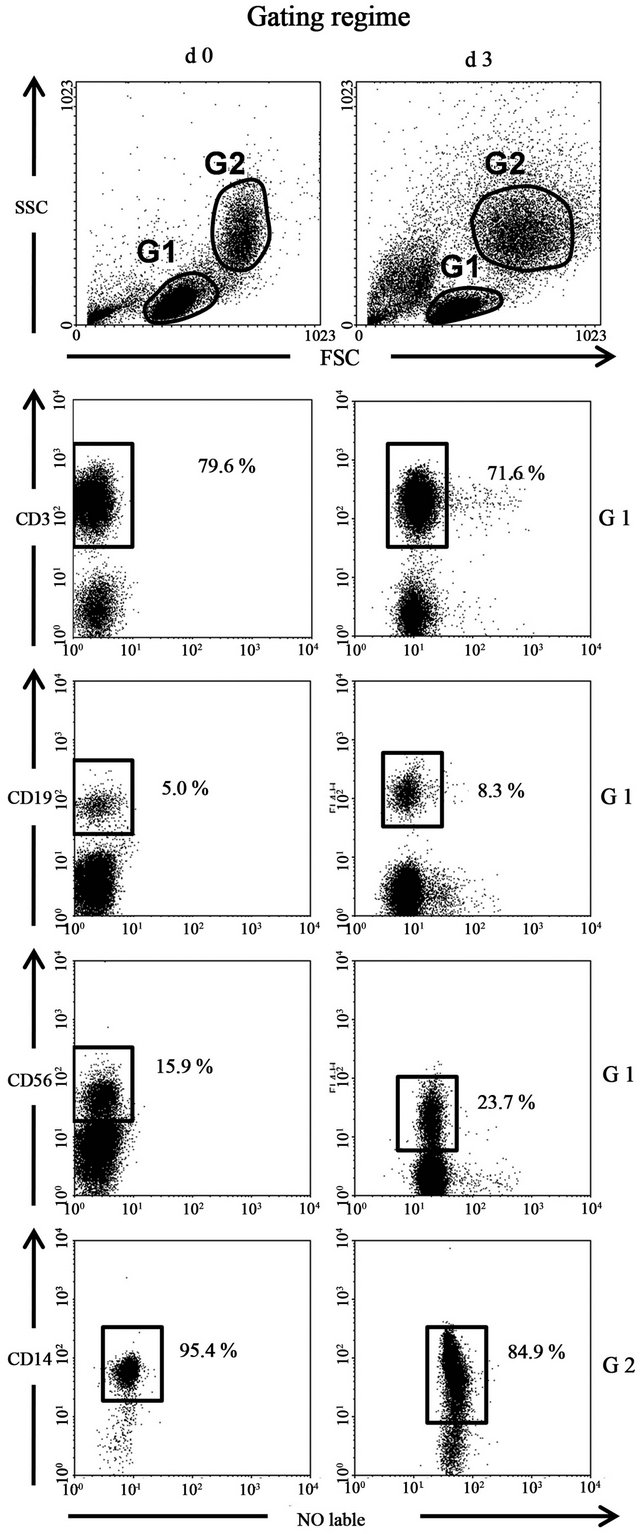
Figure 1. The frequencies of T cells (CD3+), B cells (CD19+), NK cells (CD56+) and monocytes (CD14+) in the responder cell populations day 0 and day 3 in an allogeneic two-way MLC.
3. RESULTS
3.1. FACS Gating Strategy
Figure 1 shows the FACS gating strategy for PBMC. The frequencies of T cells (CD3+), B cells (CD19+), NK cells (CD56+) and monocytes (CD14+) are shown in the FACS plot of normal PBMC and day 3 responder cells in an allogeneic two-way MLC stained with cell type specific antibodies. As shown, the frequency of the four major cell types changes only slightly during the three days of MLC. Numbers are the percent of labeled cells in the respective gates.
3.2. B7-H4-Ig Reacts with the Majority of Monocytes and NK Cells in Normal and Alloantigen-Activated PBMC
PBMCs from two donors were set up in a standard two-way MLC. On day 0 and day 3 the cells were harvested and treated with Synagis for 15 minutes (to block potential B7-H4-Ig binding to Fc receptors). Then the cells were exposed to B7-H4-Ig or PBS for 1 hr on ice. After washing the cells were stained for B7-H4 and CD3, CD19, CD56 or CD14, as indicated in Figure 2. Staining with isotype-matched mAbs was used to set cut-off limits. PBMCs were first incubated with B7 H4-Ig and subsequently with a B7-H4 antibody (Figure 2). B7-H4 antibody positivity on subpopulations of monocytes, NK cells and B cells indicate the expression of a putative receptor forB7-H4-Ig on these cell types. The numbers in the figure represent the percentages of putative B7-H4 receptor expressing cells of the indicated cell subset. Virtually all resting monocytes and around 75% of the CD56+ NK cells bind B7-H4-Ig, whereas 2-10% of B cells do so (in three comparable binding experiments). During the course of the MLC one third of the monocytes lose their putative B7-H4 receptors and so do all of the B cells. NK cells, in contrast, express this receptor at a relatively stable frequency of about 75%. Neither normal, nor allo-activated T cells appear to bind B7-H4-Ig.
3.3. B7-H4-Ig Does Not React with Activated T Cells
Previous reports have shown that anti-CD3 activated human T cells react with B7-H4-Ig constructs [4]. To test this hypothesis in our hands and under our assay formats, PBMC were subjected to two consecutive rounds of adherence to plastic and the non-adherent T cells were activated for three days with immobilized anti-CD3 antibody. Then they were exposed to B7-H4-Ig or PBS, washed and stained with PE conjugated anti-B7H4 antibody. Figure 3 shows one of two independent FACS analyses. The activated T cells appear not to express re ceptors for B7-H4-Ig. In contrast, the leftover non-CD3+ cells from the same culture, i.e. mostly monocytes and NK cells, bind B7-H4-Ig. This result is consistent with data from the flow cytometric analysis of cells in MLC.

Figure 2. B7-H4-Ig constitutively binds to human NK-cells and monocytes. PBMCs from two donors were prepared on day 0. A standard two-way MLC were prepared. On day 0 and day 3 the cells were exposed to 100 μg/ml B7-H4-Ig (gray curve) or PBS (black curve) for 1 h on ice. After washing the cells were co-stained with anti-B7-H4 and antiCD3, anti-CD19, anti-CD56 or anti-CD 14, as indicated; staining with isotype-matched mAbs was used to set cut-off limits. The number in the plots represents percentage B7-H4 positive cells of the indicated cell subset (gated). One of at least three independent experiments with similar results is depictured.
3.4. B7-H4-Ig in Allogeneic MLC
To assess the general effect of soluble B7-H4-Ig on the adaptive immune system in vitro, cell proliferation in oneand two-way allogeneic MLCs was studied. As shown in Figures 4 and 5, the presence of B7-H4-Ig in MLC apparently results in an increase in proliferation in a dose-dependent manner when compared with the medium control. This effect was only significant in one-way MLC before correcting for multiple testing (Benjamini-Hochberg correction). The non-significant stimulatory tendency holds also when the two-way MLC is depleted for CD8 T cells (Figure 5). KS, a non-functional mutant of B7-DC-Ig fusion protein similar to B7-H4-Ig, did not have any effect in MLC experiments.
Figure 6 shows that the stimulatory pattern of soluble B7-H4-Ig in MLC illustrated in Figure 4 was also noticed
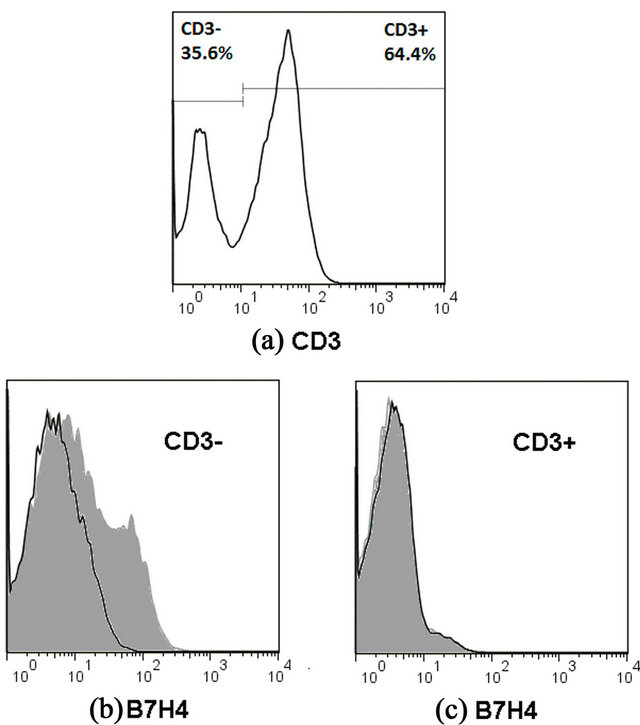
Figure 3. T cells were enriched two-fold from PBMC by non-adherence (a) and stimulated for-three days with immobilized anti-CD3 antibody. Cells were then incubated for 1 hour with PBS (black curve) or B7-H4-Ig (grey shade), washed and stained with PE conjugated anti-B7-H4 and APC conjugated anti-CD3 antibodies. B7-H4-Ig binds to CD3-cells (b) but not to CD3+ T cells (c). One of two independent experiments is shown.
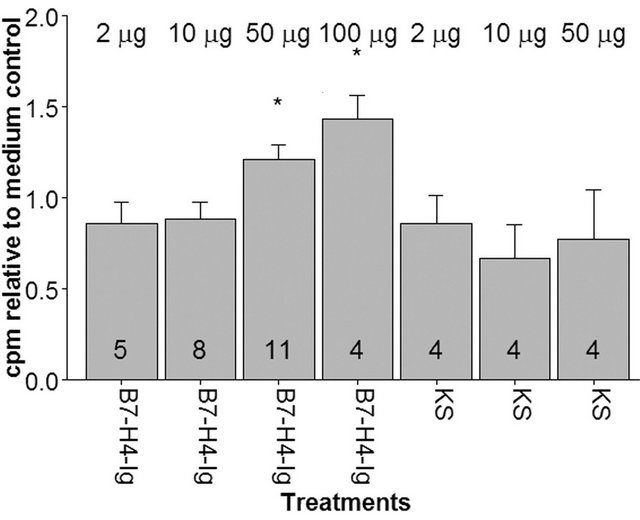
Figure 4. The presence of soluble B7-H4-Ig in one-way MLC has a non-significant tendency to enhance proliferation. Numbers in the columns represent the number of individual experiments. The same responder and stimulator cell donors were used in all experiments. Treatments are normalized in relation to their respective medium controls (mean value around 5000 cpm). Bars represent SEM values. Asterix denotes significant differences between medium control and treatments in a one-sample t-test analysis (p < 0.05). This difference is not significant after the correction for multiple testing.
in individual MLC with four different responders. Thus, the results from the MLC experiments show that B7-H4-

Figure 5. The presence of soluble B7-H4-Ig in a regular two-way MLC (left) and a two-way MLC depleted for CD8 T cells (right) has a nonsignificant tendency to enhance proliferation. Numbers in the columns represent the number of individual experiments. The same responder and stimulator cell donors were used in all experiments. Treatments are normalized in relation to their respective medium controls (mean value around 14,000 cpm). Bars represent SEM values.

Figure 6. The presence of medium, KS or B7- H4-Ig (50 μg/ml) in one-way MLC with five different responder donors against the same stimulator donor. Bars represent SD values.
Ig, in spite of its binding to monocytes, does not interfere with alloantigen-induced T cell proliferation.
3.5. Effect of Soluble B7-H4-Igon Anti-CD3 Antibody Activated T Cells
Since it was reported that B7-H4-Ig fusion proteins inhibit anti-CD3 antibody-driven T cell activation [1-4], we studied the effect of soluble and plastic immobilized B7-H4-Ig on T cells exposed to immobilized anti-CD3 antibody. Figure 7 shows the effect of soluble B7-H4-IgKS mutant and Synagis on T cell proliferation. As shown in the figure B7-H4-Ig added to the cultures at 2 μg/ml - 50 μg/ml did not influence proliferation.
3.6. Effect of Immobilized B7-H4-Ig on Anti-CD3 Antibody Activated T Cells
To reproduce previous in vitro experiments demonstrating the inhibitory effect of B7-H4-Ig onT cells [1-4], immobilized, instead of soluble B7-H4-Ig was investigated. As shown in Figure 8, a strong, significant, dose dependent inhibitory effect was found both for B7-H4-Ig but also for the control agents, KS mutant and, to a lesser

Figure 7. Non-adherence enriched T cells were exposed to immobilized anti-CD3 antibody in the presence of 2,10 and 50 μg/ml of soluble B7-H4-Ig, KS mutant or Synagis for three days). Treatments are normalized in relation to their respective medium controls (mean value around 14,000 cpm). Numbers in columns represent the number of individual experiments and bars represent SEM values.

Figure 8. Non-adherence enriched T cells were exposed to immobilized anti-CD3 antibody for three days. Immobilized B7-H4-Ig, KS mutant and Synagis were added in doses shown in the figure. Treatments are normalized in relation to their respective medium controls (mean around value 14,000 cpm). Numbers in and above columns represent number of individual experiments and bars represent SEM values. Asterix denotes significant differences between medium control and treatments in a one-sample t-test analysis (p<0.05) before application of multiple testing correction. This difference is still significant after the correction.

Figure 9. Immobilized anti-CD3 antibody-induced T cell proliferation in the absence (black curve) or presence (light gray curve) of immobilized B7-H4-Ig.
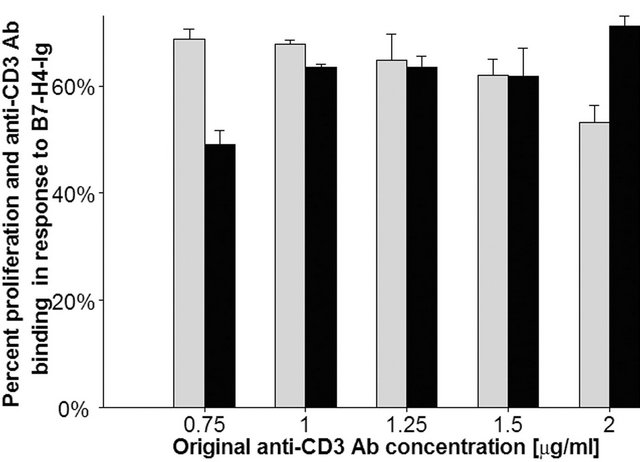
Figure 10. Amounts of immobilized anti-CD3 antibody is reduced by the presence of 50 μg/ml immobilized B7-H4-Ig to approximately 60% of its original concentration, which correlates with a reduction in T cell proliferation to about the same level. Data represent the mean of two independent experiments, bars represent SD.
degree, for Synagis.
3.7. Immobilized B7-H4-Ig Competes with Anti-CD3 Antibody for Binding to the Culture Well
T cell activation experiments using titrated doses of overnight immobilized anti-CD3 antibody were carried out. Figure 9 shows a typical stimulation experiment in the absence or presence of 2 hrs immobilized 50 μg/ml B7-H4-Ig in the culture essentially as described by Sica et al. [2]. Relationship between anti-CD3 concentration and proliferation, both with and without B7-H4-Ig, fits a logistic model, which allows one to calculate what reduction of anti-CD3 antibody concentration would result in a similar reduction in proliferation as the reduction seen after addition of B7-H4-Ig. Simultaneously, the amount of anti-CD3 antibody bound to the culture wells in the absence and presence of B7-H4-Ig was determined by ELISA assay.
As seen in Figure 10, in the presence of B7-H4-Ig the binding of anti-CD3 antibody at 1 - 1.5 μg/ml is reduced to about 60% of its original concentration, which correlates with a reduction in T cell proliferation to about the same level. Thus, the reason for the apparent B7-H4-Iginduced inhibition (as well as that of KS mutant and Synagis) of anti-CD3 antibody-induced T cell proliferation may be caused by its displacement of plastic bound anti-CD3 antibody.
3.8. Effect of B7-H4-Ig on NK and LAK Cell Activity
Freshly harvested and 24 hrs cytokine exposed PBMC were incubated for 4 hrs with 51Cr labeled K562 and Daudi cells to assay for NK cell and LAK cell activity, respectively. Although B7-H4-Ig binds to the majority of normal and cultures NK cells (Figure 2) it does not influence the killing activity (Figure 11).
4. DISCUSSION
B7-H4 is a member of the B7 superfamily of co-signaling molecules. The mature B7-H4 molecule is a 50 kDa - 80 kDa in size, glycosylated and may in murine cells be GPI-linked [1,6]. The V region domains of mouse and human B7-H4 share more than 90% amino acid sequence identity [2]. When cells were preexposed to B7-H4-Ig and then reacted with anti-human B7-H4 antibody, all monocytes (CD14+), almost 80% of the NK cells (CD56+) and less than 10% of B cells (CD19+) stained positive in normal PBMC, whereas neither nor-
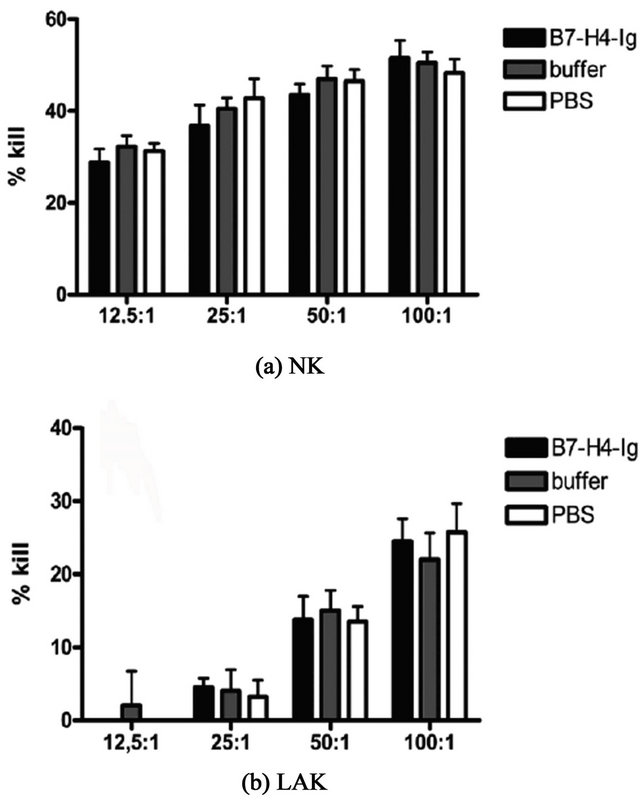
Figure 11. Four hours NK and LAK cell killing activity in freshly harvested blood mononuclear cells (MNC) in the presence of B7- H4-Ig 100 μg/ml, formulation buffer, and PBS. X-axis shows the effector: target cell ratio. Bars indicate SD values. The experiment was repeated twice with similar results from another blood donor.
mal, nor activated T cells did. B7-H4-Ig binding to day 3 alloantigen activated cells was reduced by 33% and 100% in monocytes and B cells respectively. Previous studies showing binding of various B7-H4-Ig fusion proteins to activated T cells in mouse and man [1,2,4] could not be confirmed for B7-H4-Ig binding in the present work. Thus, human T cells activated for 72 hrs by immobilized anti-CD3 antibody did not bind B7-H4-Ig.
This discrepancy may be to due subtle differences in the T cell activation procedures, i.e. the absence or presence of co-stimulatory anti-CD28 antibody as well as differences in the generation process of the B7-H4-Ig fusion protein reported in this study. And the ones used in previous studies.
Previous studies suggest that the unidentified cell membrane receptor which ligates with B7-H4-Ig fusion protein is different from co-regulatory CD28 superfamily members such as BTLA, CTLA4, and PD-1 [6,14] probably reflecting the fact that B7-H4 shares less than 25% amino acid homology in its extracellular portion with other members of the B7 family [2].
In vitro exposure to B7-H4-Ig fusion protein has been shown to inhibit anti-CD3 antibody-dependent induction of T cellactivation and proliferation [1-3] by interference with co-stimulatory pathways downstream for TCR signaling such as ERK, JNK and AKT thereby inhibiting secretion of IL-2 [16]. Using T cell proliferation in a one-way allogeneic MLC as a read-out system for B7-H4 Ig-mediated immune regulatory activity, we were unable to show any inhibitory effect of B7-H4-Ig on T cell proliferation. On the contrary, there was a moderate, but non-significant (when corrected for multiple testing) tendency for soluble B7-H4-Ig to stimulate alloantigeninduced proliferation. KS, a non-functional mutant of B7-DC-Ig fusion protein did not have any effect in the MLC experiments, suggesting that the observed stimulatory tendency is specific for soluble B7-H4-Ig. In support of our MLC data, B7-H4 expressed by renal tubular epithelial cells promotes T cell proliferation and IL-2 and IFN secretion in vitro [8]. Furthermore, a soluble form of B7-H4 was shown exacerbating RA disease development via in vivo gene transfer [11]. Although the majority of in vitro experiments conclude that B7-H4 primarily function as an immune co-inhibitor [1-5], our data from the MLC experiments and those from Chen et al. [8] may point in an opposite direction when assaying for reactivity in more complex in vitro systems such as MLC. In this context, it is also worth to consider that B7-H4 gene deleted mice, do not (as expected) show enhanced T cell responses to activation stimuli in vitro and a just slightly lowered parasite burden upon an experimental parasite infection [17] observations, which suggest that the B7 - H4 molecule, more than being a negative regulator by itself, acts together with other co-regulatory molecules in the fine tuning of immune response.
Previous data have clearly demonstrated that B7-H4- Ig mediates strong inhibition of immobilized anti-CD3 antibody-induced mouse and human [1-4] and human T cell proliferation. In contrast to these studies, we were unable to show inhibitory effects of soluble B7-H4-Ig added to human T cells exposed to immobilized antiCD3 antibody under the specific conditions tested as noted above. For the human study [4] the discrepancy might reflect different spatial conformations and receptor binding avidity between the GST/hB7-H4 fusion protein and B7-H4-Ig, respectively. When both anti-CD3 antibody and B7-H4-Ig were immobilized, B7-H4-Ig exerted a strong, dose-dependent inhibitory effect on T cell proliferation in accordance to what has been suggested for murine T cells [1-3,16], but also immobilized the KS control protein showed nearly a similar degree of inhibition. When the amount of immobilized anti-CD3 antibody was monitored simultaneously by ELISA and T cell proliferation in the absence or presence of B7-H4-Ig, most, if not all, of the inhibition was found to result from B7-H4-Ig mediated competitive displacement of the immobilized anti-CD3 antibody. Thus these data suggest that soluble or immobilized B7-H4-Ig per se does not interfere with anti-CD3 antibody-mediated activation of human T cells. Although human and mouse T cells might show different levels of sensitivity to the effects of B7-H4-Ig fusion protein in vitro, previous studies in mice [1-3,16] used immobilized anti-CD3 antibody and B7- H4-Ig fusion protein without controlling for fusion protein-mediated displacement of anti-CD3 antibody.
B7-H4-Ig was shown to bind to the majority of NK cells present in PBMC but we found no significant effect of B7-H4-Ig on the cytotoxic activity of PBMC against the NK target K562 or the LAK target Daudi cells.
In conclusion, the B7-H4-Ig fusion protein binds to a number of cell populations present in PBMC, under the conditions tested. In these particular studies we were unable to show any significant effects of the presence of soluble B7-H4-Ig in oneand two-ways MLCs, or on NK cell and LAK cell activity present in PBMC. The apparent inhibitory effect of immobilized B7-H4-Ig on the activation of T cells with immobilized anti-CD3 antibody is caused by B7-H4-Ig-mediated displacement of anti-CD3 antibody from the culture well.
5. ACKNOWLEDGEMENTS
We thank the staff at Amplimmune Inc. for critical reading and comments to this manuscript and for providing B7-H4-Ig and control reagents. This work was supported by grants from Else og Mogens Wedell Wedelsborg Foundation. Aase og Ejnar Danielsens Foundation, Aage and Johanne Louis-Hansens Foundation, Dagmar Marshalls fond, Brødrende Hartmans fond, Lægeviden-skabens fremme Foundation.
REFERENCES
- Prasad, D.V.R., Richards, S., Mai, X.M. and Dong, C. (2003) B7S1, a novel B7 family member that negatively regulates T cell activation. Immunity, 18, 863-873. doi:10.1016/S1074-7613(03)00147-X
- Sica, G.L., Choi, I.H., Zhu, G.F., Tamada, et al. (2003) B7-H4, a molecule of the B7 family, negatively regulates T cell immunity. Immunity, 18, 849-861. doi:10.1016/S1074-7613(03)00152-3
- Zang, X., Loke, P., Kim, J., Murphy, K., Waitz, R. and Allison, J.P. (2003) B7x: A widely expressed B7 family member that inhibits T cell activation. Proceedings of the National Academy of Sciences, 100, 10388-10392. doi:10.1073/pnas.1434299100
- Mao, Y.X., Chen, Y.J., Ge, Y., Ma, H.B., et al. (2006) Recombinant human B7-H4 expressed in Escherichia coli inhibits T lymphocyte proliferation and IL-2 secretion in vitro. Acta pharmacologica Sinica, 27, 741-746. doi:10.1111/j.1745-7254.2006.00338.x
- Kryczek, I., Wei, S., Zou, L., Zhu, G., et al. (2006) Cutting edge: Induction of B7-H4 on APCs through IL-10: Novel suppressive mode for regulatory T cells. Journal of Immunology, 177, 40-44.
- Choi, I.H., Zhu, G., Sica, G.L., Strome, S.E., Cheville, J.C., Lau, J.S., Zhu, Y., Flies, D. B., Tamada, K. and Chen, L. (2003) Genomic organization and expression analysis of B7-H4, an immune inhibitory molecule of the B7 family. Journal of Immunology, 171, 4650-4654.
- Chinnadurai, R. and Grakoui, A. (2010) B7-H4 mediates inhibition of T cell responses by activated murine hepatic stellate cells. Hepatology, 52, 2177-2185. doi:10.1002/hep.23953
- Chen, Y., Yang, C., Xie, Z., Zou, L., Ruan, Z., Zhang, X., Tang, Y., Fei, L., Jia, Z. and Wu, Y. (2006) Expression of the novel co-stimulatory molecule B7-H4 by renal tubular epithelial cells. Kidney International, 70, 2092-2099.
- Zou, W. and Chen, L. (2008) Inhibitory B7-family molecules in the tumour microenvironment. Nature Reviews, Immunology, 8, 467-477. doi:10.1038/nri2326
- Kryczek I., Zou L., Rodriguez P., Zhu, G. et al. (2006) B7-H4 expression identifies a novel suppressive macrophage population in human ovarian carcinoma. Journal Experimental Medicine, 203, 871-881. doi:10.1084/jem.20050930
- Azuma T., Zhu G.F., Xu H.Y., Rietz A.C., et al. (2009) Potential Role of Decoy B7-H4 in the Pathogenesis of Rheumatoid Arthritis: A Mouse Model Informed by Clinical Data. Plos Medicine, 6, Article ID: e1000166. doi:10.1371/journal.pmed.1000166
- Wei J., Loke P., Zang X. and Allison J.P. (2011) Tissue-specific expression of B7x protects from CD4 T cellmediated autoimmunity. Journal Experimental Medicine, 208, 1683-1694. doi:10.1084/jem.20100639
- Wang X. Hao J., Metzger D.L., Mui A., et al. (2011) Early treatment of NOD mice with B7-H4 reduces the incidence of autoimmune diabetes. Diabetes, 60, 3246-3255. doi:10.2337/db11-0375
- Chen L. (2004) Co-inhibitory molecules of the B7-CD28 family in the control of T-cell immunity. Nature Reviews Immunology, 4, 336-347. doi:10.1038/nri1349
- Geisler C., Plesner T., Pallesen G., Skjødt et al. (1988) Characterization and expression of the human T cell receptor-T3 complex by monoclonal antibody F101.01. Scandinavian Journal of Immunology, 27, 685-696. doi:10.1111/j.1365-3083.1988.tb02402.x
- Wang X., Hao J., Metzger D.L., Ao Z., et al. (2012) B7-H4 treatment of T cells inhibits ERK, JNK, p38, and AKT Activation. PloS One, 7, Article ID: e28232. doi:10.1371/journal.pone.0028232
- Suh W.K., Wang S., Duncan G.S., Miyazaki Y., Mak, T.W., et al. (2006) Generation and characterization of B7-H4/ B7S1/ B7x-deficient mice. Molecular and Cellular Biology, 26, 6403-6411. doi:10.1128/MCB.00755-06
NOTES
*Shared first author position.