Protective Effect of Catalpol on Myocardium in Rats with Isoprenaline-Induced Myocardial Infarcts via
Angiogenesis through Endothelial Progenitor Cells and Notch1 Signaling Pathway
626
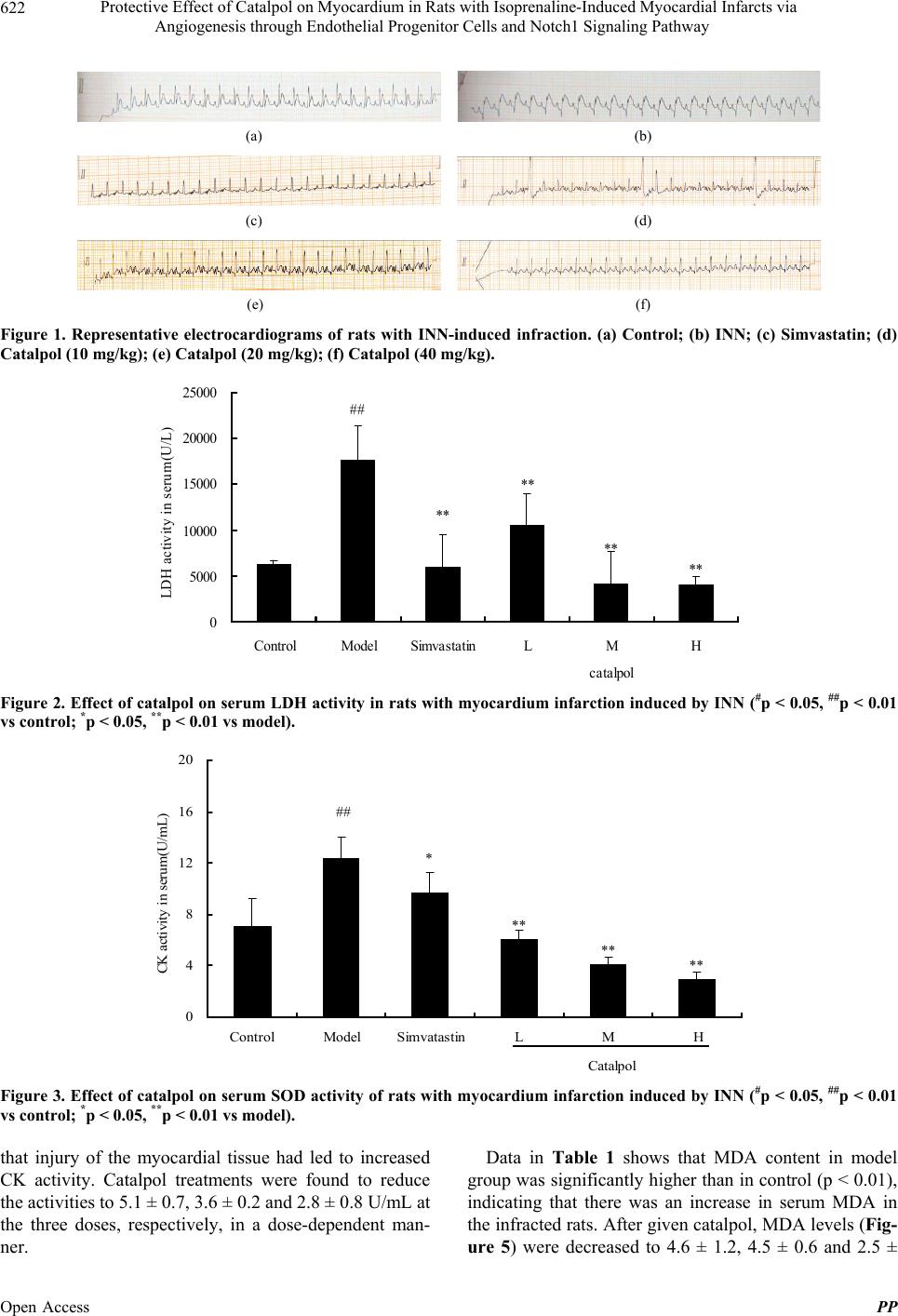
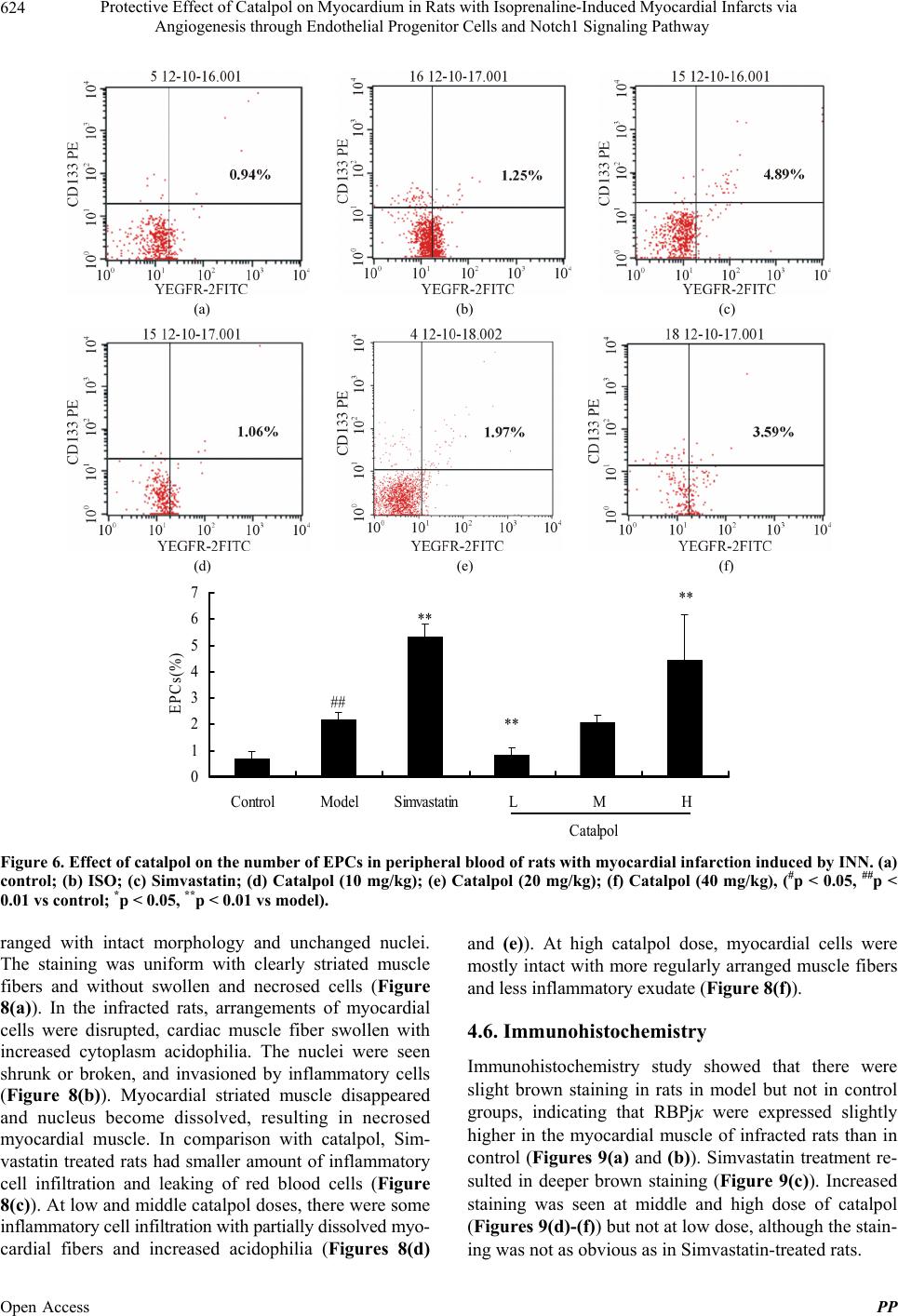
showed that the size of infracting areas observed by TTC
was in line with the data from the enzyme and biomarker
assays. These results all showed that catalpol is an effec-
tive protective agent to myocardial ischemia.
EPCs are the endothelial progenitor cells. They are
involved in embryonic vasculogenesis, angiogenesis after
birth and repair of endothelial injury in blood vessels
[13,14]. Studies have shown that EPCs in bone morrow
trend to home to ischemic tissues. Once arriving in the
ischemic tissues, they differentiate into myocardial cells
and endothelial cells, to repair the impaired tissues. Nor-
mally, EPCs account for 0.1% of the peripheral blood.
When ischemia occurs, bone morrow-derived EPCs are
mobilized to enter peripheral blood at an amount that is
not sufficient for the repair. Therefore, promotion of an-
giogenesis in myocardium tissue through clinical treat-
ments is currently the hotspots of research and practice in
treatment of myocardial ischemia. How to increase the
proliferation and differentiation of EPCs is a new direc-
tion in treatment of coronary heart diseases. Clinically,
ischemic diseases, particularly coronary heart diseases,
are always associated with one or more risk factors for
cardiovascular system. The risks are negatively related to
the number of EPCs, which are considered as prognosis
indicator for coronary heart diseases [15]. There are a
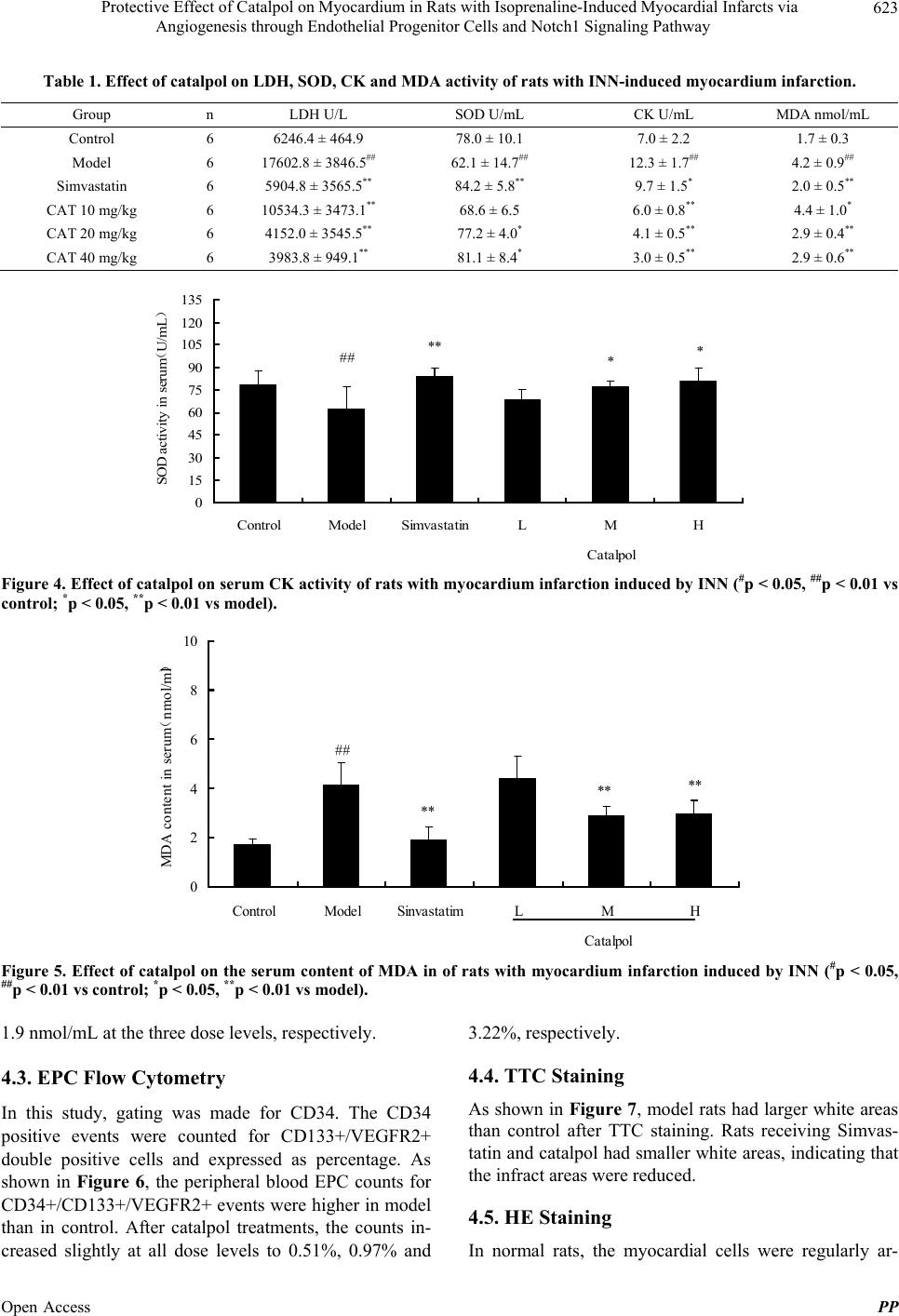
number of methods to determine the amounts of EPCs. In
most studies, CD34+, VEGFR-2+, and CD133+ are three
mostly frequently used indicators. In this study, we in-
vestigated the CD34+, VEGFR-2+, and CD133+ events
in peripheral blood, and found that EPCs were mobilized
to peripheral blood in the infracted rats with or without
drug treatment. These findings confirmed that EPCs are
released from bone marrow to peripheral blood when the
rats are stressed with dramatic shocks such as ischemia.
Notch signal pathway is first discovered in Drosophila,
and made up of receptors (Notch1, Notch 2, Notch 3, and
Notch 4), ligands (Jagged1, Jagged 2, Dll-1, Dll-3 and
Dll-4) and DNA binding protein CSL. It has been shown
that the differentiation of endothelial cells is mainly
regulated via Notch signal pathway, and the endothelial
cells have shown the potential to differentiate into artery
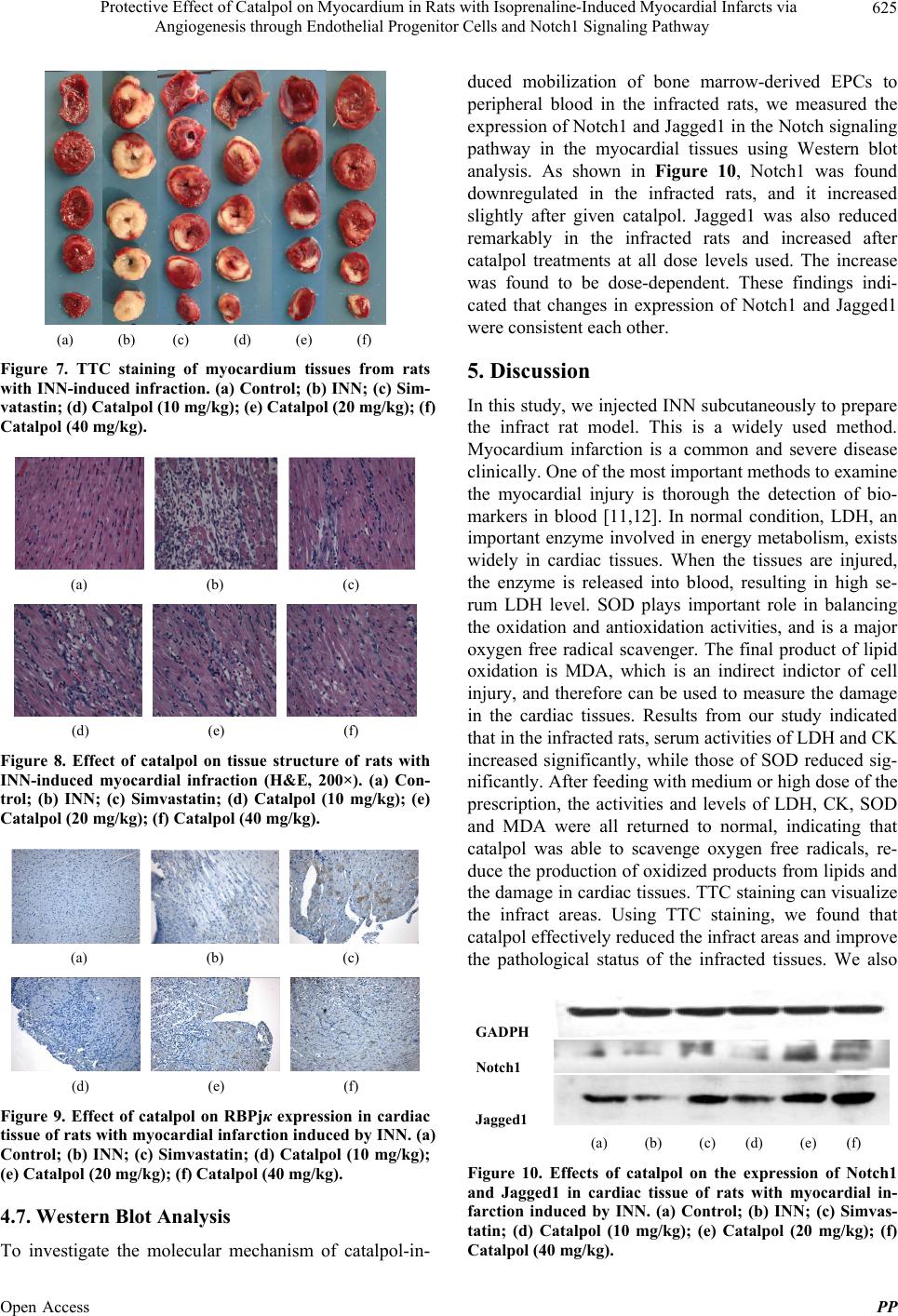
and vein before blood perfusion [16]. Notch/Jagged1 is a
newly discovered important angiogenesis factor. Western
blot analysis indicated that the expression of Notch1 re-
ceptor in the myocardium was reduced remarkably in the
infracted rats and in the normal rats [17]. Feeding of the
rats with the drugs increased the expression, in a dose
dependent way in case of catalpol, where high and me-
dium doses were better than low dose. Therefore, we
speculate that catalpol may activate Notch signal path-
way to promote the differentiation and proliferation of
endothelial cells in the infracted rats, and to improve the
oxygen supply to ischemic and injured myocardial tis-
sues, resulting in protection to myocardium and reduced
infracted area.
6. Acknowledgements
This research was supported by National Natural Science
Foundation of China (No. 81060295).
REFERENCES
[1] T. Asahara, T. Murohara, A. Sullivan, et al., “Isolation of
Putative Progenitor Endothelial Cells for Angiogenesis,”
Science, Vol. 275, No. 5302, 1997, pp. 964-966.
http://dx.doi.org/10.1126/science.275.5302.964
[2] T. Asahara, T. Takahashi, H. Masuda, et al., “VEGF Con-
tributes to Postnatal Neovascularization by Mobilizing
Bone Marrow-Derived Endothelial Progenitor Cells,” The
EMBO Journal, Vol. 18, No. 14, 1999, pp. 3964-3972.
http://dx.doi.org/10.1093/emboj/18.14.3964
[3] T. Takahashi, C. Kalka, H. Masuda, et al., “Ischemia-and
Cytokine-Induced Mobilization of Bone Marrow-Derived
Endothelial Progenitor Cells for Neovascularization,” Na-
ture Medicine, Vol. 5, No. 4, 1999, pp. 434-438.
http://dx.doi.org/10.1038/7434
[4] T. Gridley, “Notch Signaling in Vertebrate Development
and Disease,” Molecular and Cellular Neuroscience, Vol.
9, No. 2, 1997, pp. 103-108.
http://dx.doi.org/10.1006/mcne.1997.0610
[5] L. T. Krebs, Y. Xue, C. R. Norton, et al., “Notch Signal-
ing Is Essential for Vascular Morphogenesis in Mice,”
Genes & Development, Vol. 14, No. 11, 2000, pp. 1343-
1352.
[6] Y. Xue, X. Gao, C. E. Lindsell, et al., “Embryonic Le-
thality and Vascular Defects in Mice Lacking the Notch
Ligand Jagged1,” Human Molecular Genetics, Vol. 8, No.
5, 1999, pp. 723-730.
http://dx.doi.org/10.1093/hmg/8.5.723
[7] S.-M. Kwon, M. Eguchi, M. Wada, et al., “Specific Jag-
ged-1 Signal from Bone Marrow Microenvironment Is
Required for Endothelial Progenitor Cell Development
for Neovascularization,” Circulation, Vol. 118, No. 2,
2008, pp. 157-165.
http://dx.doi.org/10.1161/CIRCULATIONAHA.107.7549
78
[8] X. Tong, Z. Yang, S. Xian, et al., “Clinical Observation
on Mobilization Effect of Bone Marrow Stem Cells with
Bushen Huoxue Fonmula in Acute Myocardial Infrac-
tion,” Liaoning Journal of Traditional Chinese Medicine,
Vol. 37, No. 10, 2010, pp. 1963-1965.
[9] X. Tong, Z. Yang, S. Xian, et al., “Study on Bushen
Huoxue Recipe Mobilizing Marrow Stem Cells of Acute
Myocadial Infraction in Rats,” Chinese Journal of Basic
Medicine in Traditional Chinese Medicine, Vol. 14, No. 8,
2008, pp. 588-591.
[10] D.-Q. Li, Y.-L. Duan, Y.-M. Bao, et al., “Neuroprotection
of Catalpol in Transient Global Ischemia in Gerbils,”
Neuroscience Research, Vol. 50, No. 2, 2004, pp. 169-
177. http://dx.doi.org/10.1016/j.neures.2004.06.009
Open Access PP