 Vol.5, No.11, 1154-1164 (2013) Natural Science http://dx.doi.org/10.4236/ns.2013.511142 The most northerly record of the sirenian Protosiren and the possible polyphyletic evolution of manatees and dugongs Cajus G. Diedrich Paleologic, Petra Bezruce 96, Zdice, Czech Republic; cdiedri@gmx.net, www.paleologic.eu Received 6 April 2013; revised 6 May 2013; accepted 14 May 2013 Copyright © 2013 Cajus G. Diedrich. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT Newly discovered remains of the early Middle Eocene (Lutetian) sirenian Protosiren (Protosi- renidae) in shark tooth rich conglomerates from a coastal delta environment northwest of the European Rhenish Massif at Fürstenau (north- western Germany), represent the most northerly occurrence of this genus w hose global distribu- tion was generally restricted to warm waters. Its presence of the remains so far north can be ex- plained by seasonal inflow of warm Tethys sur- face water into the cool, upwelling-influenced, basin. The existence of two discrete centers of sirenian evolution can be explained by the opening of the Atlantic and the upwelling that separated the North American warm water fau- nal province from those of Africa and Eurasia. A slightly modified evolutionary model is pre- sented in which the oldest Early Eocene mana- tee sirenians evol ved in the Caribbea n of Centr al America. Protosiren, however, appears to have developed polyphyletically along the African coastline of the Tethys, and represents the old- est known dugong ancestor. Younger (Oligo- cene) European sirenian skeletons of Halith- erium and Anomotherium are included in the phylostratigraphic model in which sirenians had generally reduced their teeth by 28 Ma as an adaptation for feeding on sea-plants (macroal- gae/seagrass). Teeth from early megatooth sharks, which preyed on sirenians, have been recorded from shallow marine Eocene and Oli- gocene coastlines of the southern proto-North Sea Basin, and shark bite marks have been found on sirenian skeletons. Keyw ords: Sirenian Remains; Early Middle Eocene; Palaeobiogeography; Olde st Sirenians of the Proto-North Sea Basin of Central Europe 1. INTRODUCTION Extant manatees and dugongs, together with the re- cently extinct Steller’s seacow, are three distinct types of modern sirenian [1] that have their earliest origins in the Late Palaeocene to Early Eocene (Figure 1) [2-4]. The most primitive skeletons of quadrupedal sirenians (Ypre- sian, Early Eocene) ever found in the shallow marine palaeoenvironments of the Middle American Caribbean are of the primitive Prorastomus and the slightly more evolved Pezosiren (Figure 1) [5]. The global fossil re- cord for sirenians in the Eocene is much poorer, as it is also in the Oligocene and Miocene [1], and therefore new discoveries, especially well dated material in a bio- diverse and palaeoecological context, are important to our understanding of the evolution and life of the basal sirenians. Early Middle Eocene (Lutetian) sirenian remains from the new sirenian locality at Dalum, near Fürstenau (north-western Germany, Figure 2) are also considered herein. The shallow coastal and slightly carbonatic Eo- cene sands at this locality have been bioturbated by crustaceans and are interrupted by a transgressive con- glomerate bed in which phosphorite nodules and verte- brate remains are concentrated [6]. Those conglomerates within the Fürstenau Formation are extraordinarily rich in shark teeth, and also contain coprolites from several different sharks, including megatooth and white shark forms [6-8]. The rich shark biodiversity, which includes about 15 species with tooth sizes larger than 5 mm, also includes more rare large, serrated teeth up to eight cen- time tres from th e megatooth Otodus and teeth up to four centimetres from the white shark Carcharodon [8]. The relationship between megatooth sharks and their first Copyright © 2013 SciRes. OPEN A CCESS 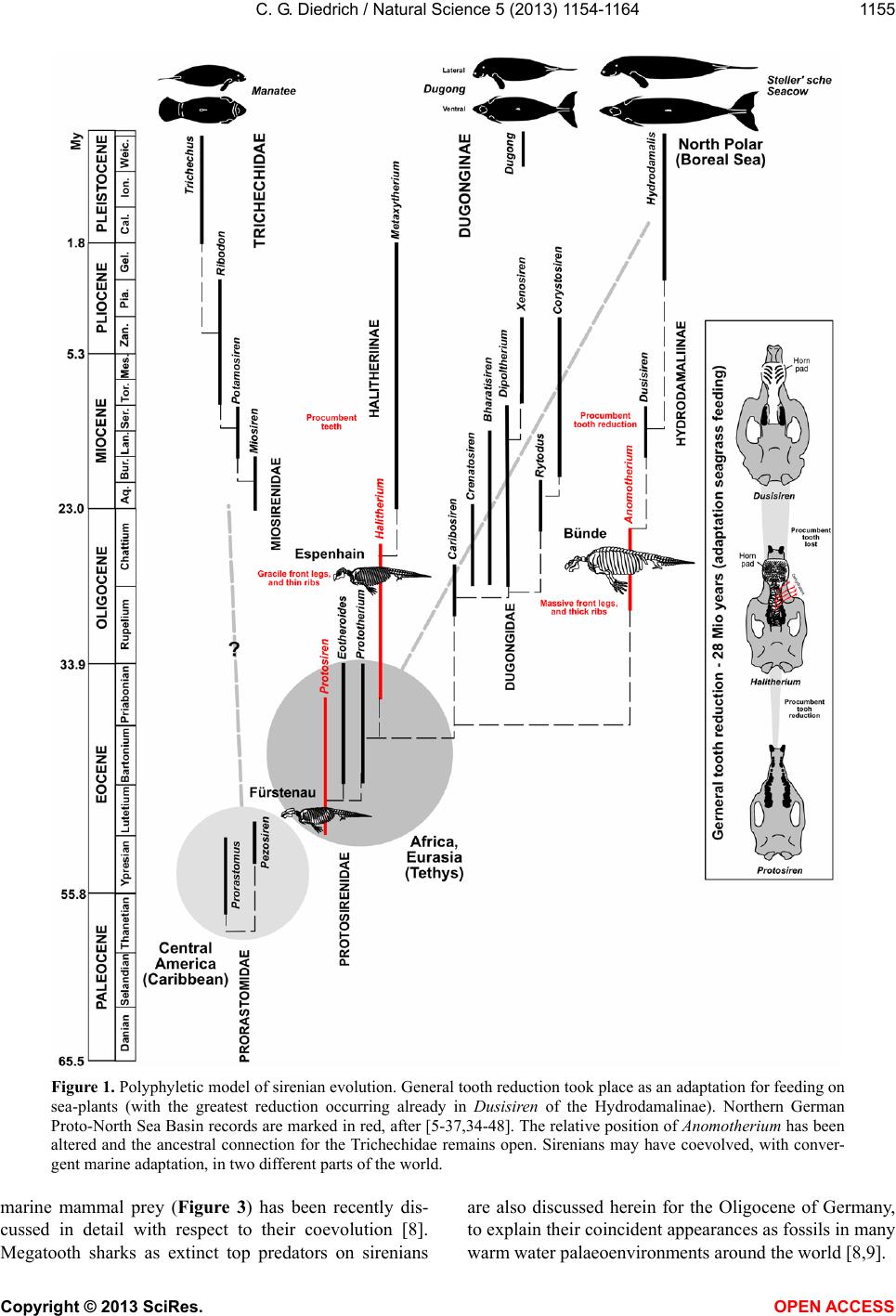 C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1155 Figure 1. Polyphyletic model of sirenian evolution. General tooth reduction took place as an adaptation for feeding on sea-plants (with the greatest reduction occurring already in Dusisiren of the Hydrodamalinae). Northern German Proto-North Sea Basin records are marked in red, after [5-37,34-48]. The relative position of Anomotherium has been altered and the ancestral connection for the Trichechidae remains open. Sirenians may have coevolved, with conver- gent marine adaptation, in two different parts of the world. marine mammal prey (Figure 3) has been recently dis- cussed in detail with respect to their coevolution [8]. Megatooth sharks as extinct top predators on sirenians are also discussed herein for the Oligocene of Germany, to explain their coincident appearances as fossils in many warm water palaeoenvironments around the world [8,9]. Copyright © 2013 SciRes. OPEN A CCESS 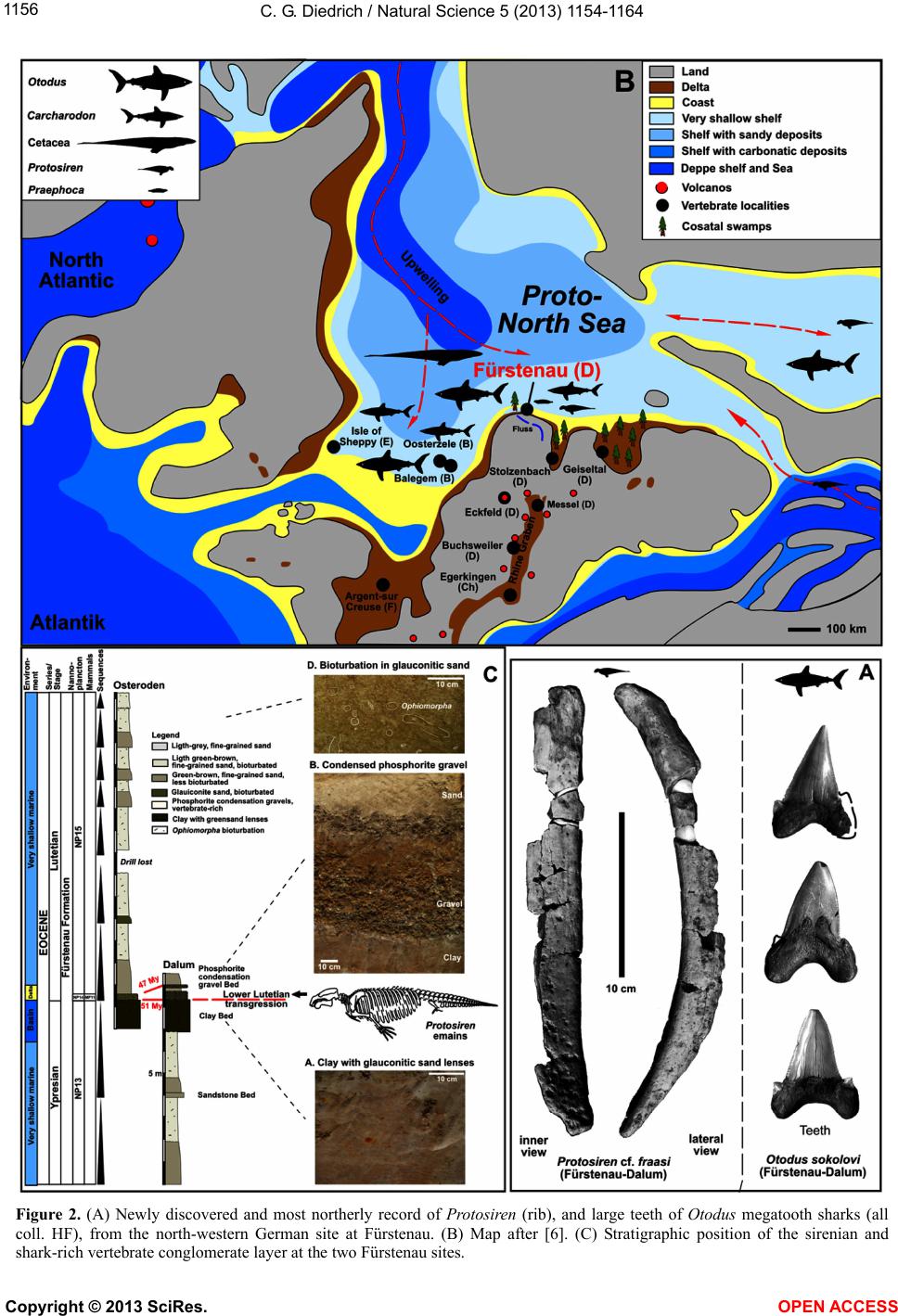 C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1156 Figure 2. (A) Newly discovered and most northerly record of Protosiren (rib), and large teeth of Otodus megatooth sharks (all coll. HF), from the north-western German site at Fürstenau. (B) Map after [6]. (C) Stratigraphic position of the sirenian and shark-rich vertebrate conglomerate layer at the two Fürstenau sites. Copyright © 2013 SciRes. OPEN A CCESS 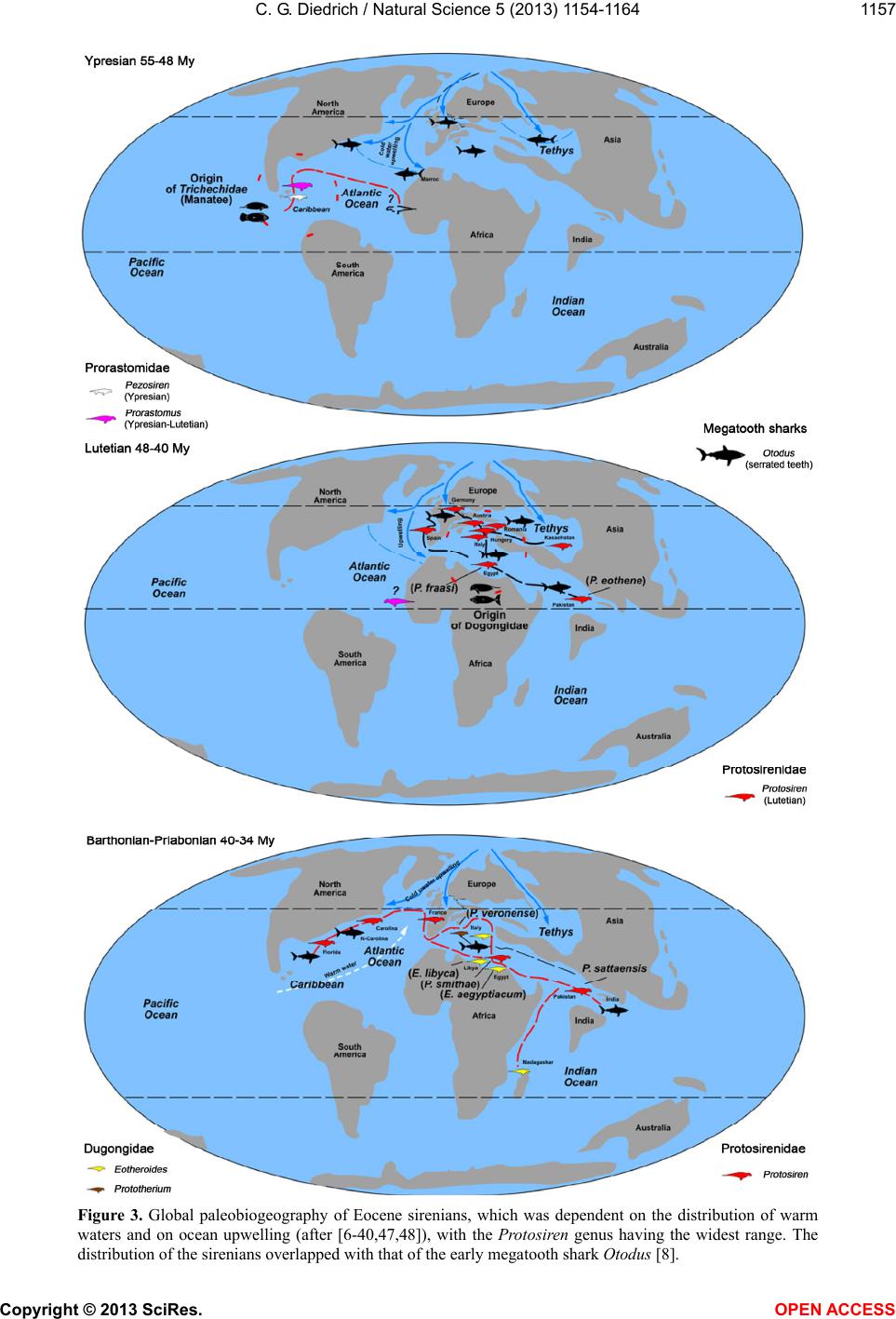 C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1157 Figure 3. Global paleobiogeography of Eocene sirenians, which was dependent on the distribution of warm waters and on ocean upwelling (after [6-40,47,48]), with the Protosiren genus having the widest range. The distribution of the sirenians overlapped with that of the early megatooth shark Otodus [8]. Copyright © 2013 SciRes. OPEN A CCESS  C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 Copyright © 2013 SciRes. OPEN A CCESS 1158 A small number of Tertiary sirenian fossils have been described from central Europe, from the proto-North Sea Basin [10-14]. Three different Paleogene (Eocene to Oligocene) genera from northern Germany are reviewed herein: Protosiren (Middle Eocene, single bones), Hali- therium (lower/basal Upper Oligocene, several skele- tons), and Anomotherium (Upper Oligocene, two skele- tons) from the proto-North Sea Basin are placed within the cladistic models based on former descriptions [1-5]. An extended model of sirenian evolution that includes a phylostratigraphic and possibly polyphyletic evolution of manatees and dugongs is presented, reviewing most of the known important global material and including new Eocene (Lutetian) fossil remains from northern Germany (Figure 2). These most northern global records of the Protosiren genus are also signifycant in the reconstruc- tion of ancient, climate-influenced palaeocurrents (e.g., cold upwellings and warm surface currents), and of global paleob iogeography. 2. MATERIAL AND METHODS About 250,000 shark teeth, 12,000 fish otoliths, some terrestrial mammal teeth, marine seal remains, shark coprolites, teleostean fish remains, and invertebrate fos- sils have been collected by H. Felker (HF) from two Eo- cene fossil sites at Bippen and Dalum, near Fürstenau (Figure 2), over the past 30 years. This collection has now been examined for marine mammal remains. About 60 higher vertebrate bone fragments are present, include- ing some mosasaur remains reworked from Upper Cre- taceous horizons, making small fragments difficult to determine macroscopically using only their osteological morphology. The sirenian material from Dalum is repre- sented by at least one almost complete sirenian rib (Fig- ure 1), a few other rib fragments, and a possible verte- bral neural arch fragment. Possible whale remains are also present but these bone fragments, especially the vertebra fragments, can as yet not be firmly distinguish from those of sirenians. The cross sections of the frac- tured sirenian ribs were examined in order to verify that they were pachyostotic ribs, with concentric growth rings. The material was then compared with a Protosiren fraasi skeleton from the Stuttgart State Museum for Natural History (SMNS), with skeletons or bones (from younger Oligocene horizons) of Halitherium schinzii from the Senckenberg Museum in Frankfurt (SMF), and with Anomotherium langewieschei remains from the Do- bergmuseum Geological Museum of Ostwestfalen-Lippe in Bünde (DMB). Megatooth shark teeth in the private collection of U. Beye (UB) are from Barleben, near Magdeburg. Finally, the teeth of predatorial megatooth sharks from Fürstenau and sirenian skeletons with evi- dence of shark-bites have also been included in these investigations in order to understand the relationships between, and palaeobiogeographical distributions of, these predators and their prey. Order Sirenia Illiger 1811 Family Protosirenidae Sickenberg 1934 Protosiren Abel 1907 Protosiren cf. fraasi Abel 1904 Material: One almost complete rib 27 cm long (Figure 2), some small rib fragments, and a possible vertebral neural arch fragment (HF collection). Comparison: Material from this genus is scarce and no complete skeletons are known. All material is therefore fragmentary, but several “species” of the Eocene Proto- siren have been established in different parts of the world, such as P. e othene Zalmout et al. (2003) [15], P. fraasi Abel (1904) [16,17], P. sattaensis Gingerich et al. (1995) [18], P. smithae Domning and Gingerich (1994) [19], and P. minima Desmarest (1822) [15-17], which are clearly in need of global revision. The almost complete rib from Germany illustrated herein (Figure 2) is very similar in shape and cross section to a middle thoracic rib from P. cf. fraasi (SMNS 11090), which is of similar age, coming from the early Middle Eocene of Egypt [16], but the surface of the German rib has been corroded by ocean waters. The inner side of the pachyostotic rib is flattened, while the outer side is convex. The articular facet on the rib capitulum is relatively flat to concave, which is typical of Protosiren ribs [16-20]. Some other fragments are also from ribs, but the exact number of ribs is difficult to estimate for the freshly fractured material. A neural arch fragment is also possibly preserved, but it is too small to permit positive identification. 3. DISCUSSION 3.1. Sirenian Species during the Eocene The substantial morphological diversity and global distribution of the Eocene sirenians Prorastomus, Pe- zosien, Protosiren, Eotheroides and Prototherium (Fig- ures 1 and 3) [1-5,15-41,47] suggests that they must have already evolved from terrestrial mammals by the late Paleocene [1-5]. Early Eocene (Ypresian): The oldest known sirenians are the Prorastomidae, of Early Eocene age. Prorastomus sirenoides Owen, 1855 and Pezosiren portelli Domning, 2011, in the late Ypresian/early Lutetian, are still primi- tive and quadrupedal, with fully developed legs. Both have been recorded from Jamaica, in the Caribbean [1,5] (Figures 1 and 3). If the newest described remain from Senegal (Prorastomidae indet.) [47] does not belong into this time-frame must remain unclear, because this strong- ly damaged vertebra could also have been reworked ma- rine from older layers. It would then, indicate the ex- change between Carribean and West-African populations (cf. Figure 3). 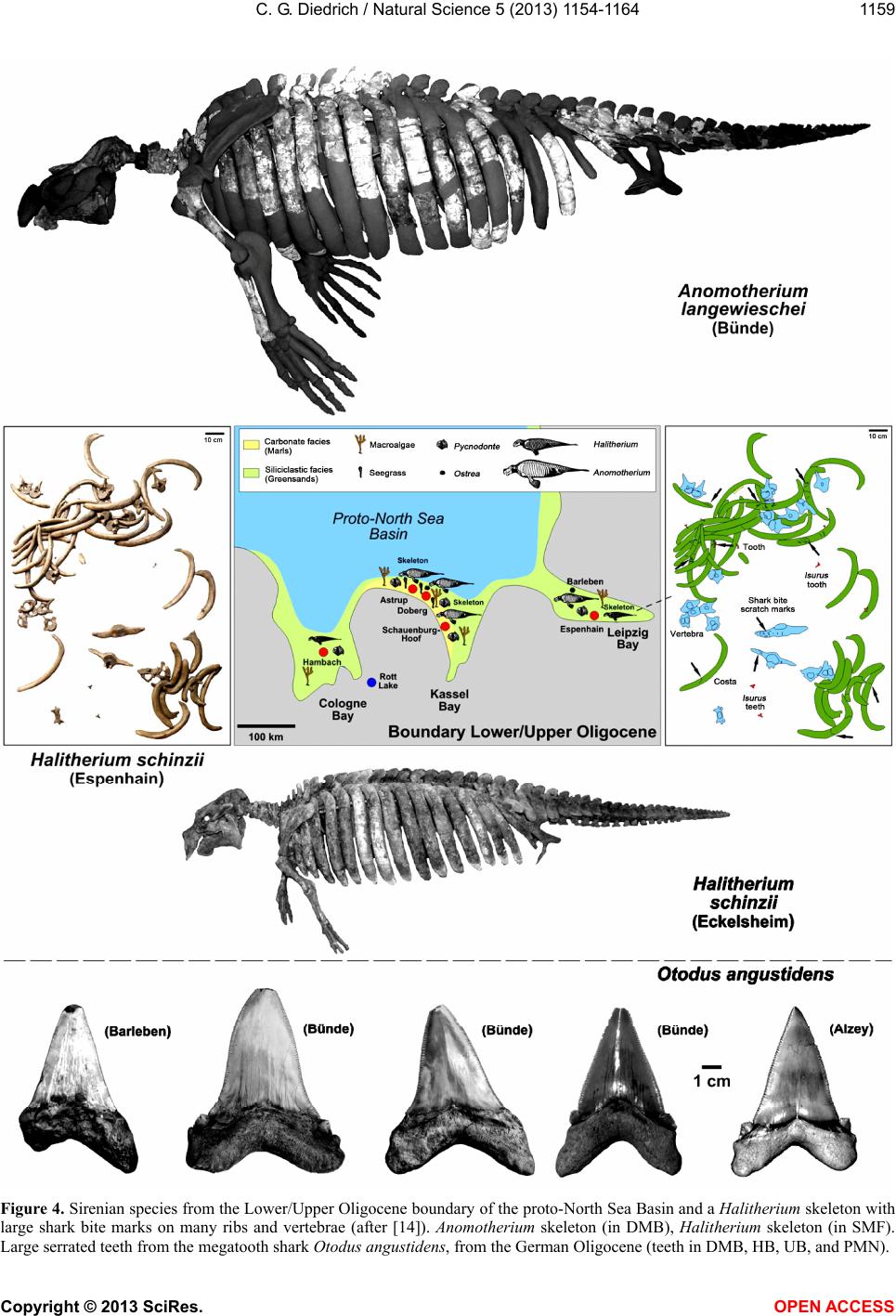 C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1159 Figure 4. Sirenian species from the Lower/Upper Oligocene boundary of the proto-North Sea Basin and a Halitherium skeleton with large shark bite marks on many ribs and vertebrae (after [14]). Anomotherium skeleton (in DMB), Halitherium skeleton (in SMF). arge serrated teeth from the megatooth shark Otodus angustidens, from the German Oligocene (teeth in DMB, HB, UB, and PMN). L Copyright © 2013 SciRes. OPEN A CCESS  C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1160 Early Middle Eocene (Lutetian): The best-preserved and most abundant sirenian specimens with a similar age to the Fürstenau material are the Protosirenidae from Egypt (Protosiren fraasi) [16,18,34], and Pakistan (Pro- tosiren eothene) [15,37] (Figures 1 and 3). Most of the material from Hungary, Germany, and Spain, as well as that from Kazakhstan, should probably be attributed to P. fraasi Abel, 1904 [16-17,19,23]. There may have been only a single species distributed throughout the Tethys (Figure 3), but the fossil record is sparse and fragmen- tary. Upper Middle Eocene to lower Late Eocene (Barto- nian): The protosiren P. sattaensis, which is represented mainly by ribs and a small quantity of other Protosireni- dae material found along the east coast of North America (Carolina, Florida), and which has also been suggested from Pakistan (Figures 1 and 3), has been accurately dated from the North American discoveries [27]. Other protosiren remains from P. smithae have been reported from Egypt [19]. The possible P. m i n ima remains from France are problematic as far as species identification and dating are concerned. Two other Dugongidae genera also developed during the Bartonian, these being Eother- oides [1,21], which has been found along the northern coastline of Africa [37,42], and Eosiren Andrews 1902, which is from the Late Eocene (Bartonian to Priabonian) (Figures 1 and 3) [1]. Late Eocene (Priabonian): The dugongid Prototherium veronense de Zigno, 1875 has been found in Italy [28] and ranged from the upper Middle to the Late Eocene (Figures 1 and 3). The E. libyca Andrews, 1902 has been recorded from the Late Eocene of Egypt [42] (Figures 1 and 3). 3.2. Origin, Palaeobiogeography and Habitat of Early European Sirenians The distribution of early sirenians is related to that of warm ocean waters with seagrass meadows [1,2,4,14]. These aquatic mammals may have had polyphyletic de- velopment from two separate centers (Figure 3). Classi- cal models [1-5] describe an origin in the warm shallow waters of Central America [5], and a subsequent disper- sal to Eurasia. The palaeooceanography of the Early Eo- cene (Ypresian), however, indicates that the opening of the North Atlantic resulted in massive upwellings of cold, arctic bottom currents, which are likely to have inh ibited any such migration. These upwellings extended not only into the proto- North Sea Basin, but also as far south as north-western Morocco, where the absence of any sirenian remains in shark-rich phosphatic b onebeds lends support to the idea that the upwellin gs presented a barrier to sirenian migra- tion (Figure 3). The modern Trichechidae and Dugongi- dae are therefore more likely to have had different ori- gins and ancestors (Figures 1 and 3), one in the Carib- bean, and the other in the very different habitat provided by the shallow, warm, marine waters off northern Africa (Figure 3). The monophyletic model is based on classi- cal monophyly [1-5], but th e “evolution of the sirenians” presented herein is a little different (Figures 1 and 3), bringing together all of the data, and leaving open any gaps in the fossil record or unclear phylogenetic rela- tionships betw een families and subfamili es. The Middle Eocene Protosiren (Protosirenidae) is be- lieved herein to be known after reviewing the descrip- tions and figures of single finds in the Lutetian from Italy, Spain [26], Hungary [16], Romania [32], Egypt [17,19], Kazakhstan, and Pakistan [15-40], and now also from Germany. These early sirenians have recently been re- corded from along the equator and further north in the Tethys, the northern Indian Ocean, the proto-North Sea Basin, and the western coastlines of the already opened North Atlantic (Figure 3) [15-40], but they always ap- pear to have been in shallow marine habitats of warm oceans, or in areas influenced by warm surface waters. The remains from the proto-North Sea Basin, which are the most northerly sirenian record for that time, are from a slightly different environment from other discoveries in that this was primarily a cold shallow marine basin, in- fluenced by up wel l ing ( Figure 2(B)) [6]. Periodic influxes of warm surface waters from the Tethys are, however, clearly documented by the presence of the remains of sirenians, whales, and megatooth sharks (Figure 2) [8], that must have come into the area from the east or southeast, at least during the Lutetian (Figure 3). The early Lutetian Protosiren appears from the re- cently discovered fossil record not to have originated in Central America, but from the coastline of northern Af- rica [1,27], as is also the case for the Early Eocene primitive quadrupedal sirenians (ancestors of Trichechi- dae). Their distribution appears to have been clearly re- stricted by the cold water upwelling, which had already started in the Paleocene. The North Atlantic therefore would have been a barrier to dispersal, which would also explain another polyphyletic development-the emergence of the Dugongidae in Eurasian coastal Tethys habitats (Figures 1 and 3). These did not arrive on the east coast of North America before the Bartonian [1,27], which would again appear to support an origin in Eurasia (Fig- ures 1 and 3). Their original habitat was more likely to have been along the more gently sloping coastlines cov- ered with seagrass (and possibly also macroalgae) that had already become the main food source for sirenians during the Eocene [1-5,14,45]. Adaptation to feeding on seagrass caused a general tooth reduction in sirenians between the Eocene and the Pliocene (Figure 1). Com- Copyright © 2013 SciRes. OPEN A CCESS  C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1161 plete incisor, premolar, and anterior molar tooth reduce- tion occurred within the Trichechidae and the Dugongi- nae, but the greatest reduction (including the anterior procumbent teeth) occurring in the Hydrodamalinae (Figure 1). Further support for a polyphyletic develop- ment is provided by the major sirenian radiation in Eura- sia (Figures 1 and 3). Within the Barthonian, the Du- gongidae Eotheroides and Prototherium developed on the shallow marine shelves of northern Africa, and in the Tethys area in general (Figure 3), as did the Protosireni- dae (a little later) which are believed to have been distant non-ancestors of modern Dugongs [1-5]. 3.3. Younger Sirenian Records from the Proto-North Sea Basin Focusing only on the pro to-North Sea Basin of central Europe, Miosirenidae and Halitherinae are now known from horizons of various ages (Figure 2), which include the Early to lower Late Oligocene Halitherium schinzii Kaup, 1838 [10,11,44] (also distributed in the Cologne and Leipzig bays, in northern Germany at Doberg, in the Molasse Basin, and in the Rhine Graben: Figure 4 [14, 46]), and the large Upper Olig ocene form Anomotherium langewieschei Siegfried, 1965 [12,14] (only recorded in the southern proto-North Sea Basin, Figure 4, with two skeletons found near Bünde [14] and Schaumburg-Hof [13,14]). The Anomotherium discoveries coincide with the dis- tribution of a carbonate facies in which indirect evidence for macroalgae and seagrass is provided by the xeno- morphic attachment surfaces of various oysters [14,45] (Figure 4). The new phylogenetic position suggested herein for Anomotherium within the evolutionary model is a pre- liminary step based on characteristics such as the ex- tremely massive thickened ribs, which are 6 - 8 cm in width (as typically seen in hydrodamalines, and similar to those in Hydrodamalis). The much larger-proportioned and more massive forelegs in Anomotherium and Hy- drodamalis (in contrast to the reduced forelegs of Halitherum; Figure 4) are also important. These postcra- nial skeleton characteristics allow the parallel-living Anomotherium/Halitherium genera to be separated from each other, especially in the proto-North Sea Basin [14]. This distinction is thus based on the large robust general habitus and body size, which in the case of Anomoth- erium is close to that of the Steller’s seacow (Hydro- damalis). Cranial characteristics can not be used due to the high degree of damage to the skull: the presence or absence of procumbent teeth would help to position Anomotherium more accurately within the systematics, which are mainly based on cranial characteristics. How- ever, the systematic position will need to be revised in the future using the two known Anomotherium skeletons from Germany The holotype is currently spread over three collections and many bone fragments have not yet been included in the skeleton but are still in boxes: the reassembled skeleton presented herein in Figure 4 is therefore not fully rep resentative of the holotype skeleton. Finally, the palaeobiogeography of Anomotherium, which is only present globally in the northern proto- North Sea Basin, seems to suggest a large sirenian al- ready adapted to seagrass feeding and temperate waters- a habitat that was already occupied by the hydrodama- lines. 3.4. The Sirenians’ Predators The megatooth shark ancestor Otodus sokolovi was already present in the Middle Eocene as a large form (with teeth up to 8 cm long, Figure 2(A)), indicating the parallel evolution of these large sharks with their main prey, the whales and sirenians [8]. Several large teeth have also been found at Fürstenau, suggesting a spatial correlation with their prey, the sirenians (and probably whales) [8], and also reflecting their presence in warm waters only. The same spatial correlation can also be reported from the Upper Oligocene horizons in the proto-North Sea Basin, where toothed whales (Eosqualodon) and sireni- ans (Anomotherium) again appear together with mega- tooth shark remains, but in this case from the more highly evolved Otodus angustidens (Figure 4) [8,14]. The bite marks that are commonly found on the ribs and vertebrae of Halitherium (Figure 4) and other sirenians [14,46] indicate that they were scavenged, and possibly preyed upon, by large sharks. In contrast, the smaller white shark ancestor Carcharodon auriculatus, from which some thousands of teeth have been found in the Middle Eocene gravels at Fürstenau, was adapted to cold water, as were its main prey, the seals [8]. 4. RESULTS Early Eocene (Ypresian) sirenians (the world’s oldest sirenians) appear to have already developed in the Car- ibbean by the Late Paleocene (Thanetian), in tropical shallow coastal waters and freshwater river environments. Two different sirenian genera, Pezosiren and Prorasto- mus, which are known from “New World” skeletons, did possibly not spread over the Atlan tic to the “Old World”, because the continents were separated by cold upwelling currents in the northern Atlantic. These quadrupedal basal sirenian forms are likely to have been the ancestors of the modern manatees (and their predecessors), which have a similar geographical distribution and also live in coastal brackish and freshwater environments. The rela- tionship between the oldest sirenians and the manatees remains unresolved and the phylostratigraphic model is Copyright © 2013 SciRes. OPEN A CCESS  C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1162 therefore not yet complete. The newly discovered marine Middle Eocene (Lu- tetian) sirenians (including Protosiren), which may have had a polyphyletic development, appear to have been evolved on the shallow marine coastlines of the Tethys, such as along northern Africa, or between Eurasia and India. The most northern discoveries (from Germany, in the proto-North Sea Basin) resulted from seasonal varia- tions between warm surface waters of the Tethys and cold upwelling currents from the North Atlantic that were responsible for a rich fish fauna and the presence of three types of mammal: seals, sirenians, and whales. The Protosirenidae are seen herein to appear to be possible polyphyletic or even “ancestors” of the Dugongidae (Figure 1), which was seen different by other authors. Within this lineage, it is placed herein as the massive European Oligocene form Anomotherium from the proto- North Sea Basin, which has only been recorded from temperate carbonate seagrass-meadow environments. This little-reported form, which is only recorded from the northern hemisphere, is best placed on the basis of anatomy (from postcranial characteristics of massive, thickened ribs and large massive forelimbs) and palaeo- geography within the Hydrodamalinae lineage, with its adaptation to temperate and cold waters. The tooth re- duction and horny pad development started in the Eocene as an adaptation to feeding on seagrass/macroalgae, con- tinued in tooth reduction by 28 Ma in the Oligocene, and is best evolved in the Hydrodamalinae; in contrast to the other forms, that the Hydrodamalinae-Dusosiren already had no procumbent teeth by the early Miocene (23 Ma). The first indirect evidence of sirenians having been hunted by megatooth sharks is found in the Middle Eo- cene of north-western Germany, where they are both (occasionally) present in the same horizons. The coinci- dental occurrence of both sirenians and megatooth sharks at several different locations around the globe in Paleo- gene to Neogene sediments also provides supporting evidence for early development of a predator-prey co- evolutionary relationship. The best examples of bite damage on ribs and vertebrae are found on skeletons of Halitherium, the most common Oligocene sirenian in Europe. Sirenians in general appear to have been hunted and killed (in a similar way to whales) by megatooth sharks (Otodus), but were also scavenged by other sharks (Isurus, and others). 5. ACKNOWLEDGEMENTS The sirenian remains described herein were discovered by the hobby palaeontologist, Mr. H. Felker, who kindly allowed the specimens to be studied. A cast of the original rib (which not a holotype) is in the public collection of the Seehundstation Nationalpark-Haus and Waloseum Museum, Vogelpflegestation Norden-Norddeich. The private company PaleoLogic was responsible for the scientific research and funding. M. Strauss from the Dobergmuseum Bünde (DMB) allowed the study of the sirenian skeleton and shark tooth material. Dr. B. H erkner from the Senkenbergmuseum Frankfurt (SMF) made the sirenian skeleton in the exhibition available. I thank H. Stapf of the Paläontologisches Museum Nierstein (PMN) for providing a photo of a shark tooth from Alzey. H. Breitkreutz (HB) permitted the study of his private tooth collection from the Doberg. U. Beye supported with a photo of a tooth in his collection from Barleben. Finally E. Manning supported much with the spell-check. REFERENCES [1] Domning, D.P. (2001) Evolution of the Sirania and Dsmostylia. In: Mazin, J.M. and de Buffrénil, V., Eds., Secondary Adaptation of Tetrapods to Life in Water, Pfeil-Verlag, Munich, 151-167. [2] Domning, D.P. (1982) Evolution of manatees. Journal of Vertebrate Paleontology, 56, 599-619. [3] Domning, D.P. and Gingerich, P.D. (1994) Protosiren smithae, new species (Mammalian, Sirenia) from the Late Middle Eocene of Wa di Hitan, Egypt. Contributions from the Museum of Paleontology of the University of Michi- gan, 29, 69-87. [4] Ripple, J. (2002) Manatees and Dugongs of the World. Voyageur Press, Hong-Kong, 144. [5] Domning, D.P. (2001) The earliest known fully quadru- pedal sirenian. Nature, 413, 625-627. http://dx.doi.org/10.1038/35098072 [6] Diedrich, C. (2012) Eocene (Lutetian) shark-rich coastal paleoenvironments of the Southern North Sea basin in Europe: Biodiversity of the marine Fürstenau formation including the earliest ancestors of the White Sharks. In- ternational Journal of Oceanography, 2012, 565326. http://dx.doi.org/10.1155/2012/565326 [7] Diedrich, C. and Felker, H. (2012) Middle Eocene shark coprolites from shallow marine and Deltaic coasts of the pre-North Sea Basin in Central Europe. New Mexico Mu- seum of Natural History and Science, Bulletin, 57, 311- 318. [8] Diedrich, C. (2013) White and megatooth shark evolution and predation origin onto seals, sirenians and whales. Natural Science, accepted. [9] Purdy, R. (1996) Paleoecology of fossil White Sharks. In: Kimley, A.P. and Ainley, D.G., Eds., Great White Sharks: The Biology of Carcharodon carcharias, Academic Press, San Diego, 67-78. http://dx.doi.org/10.1016/B978-012415031-7/50009-4 [10] Voss, M. (2006) Funde von Halitherium (Sirenia) aus dem Unteroligozän des Ruhr-Gebietes. Abtract, Arbeits- kreis Wirbeltierpaläontologie der Paläontologischen Ge- sellschaft, 2006, 33. [11] Müller, A. (1983) Fauna und Paläoökologie des marinen Miteloligozäns der Leipziger Tieflandsbucht (Böhlener Schichten). Altenburger naturwissenschaftliche Forschun- gen, 2, 1-152. [12] Siegfried, P. (1965) Anomotherium langewieschei n.g. n.sp. (Sirenia) aus dem Ober-Oligozän des Dobergs bei Copyright © 2013 SciRes. OPEN A CCESS  C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 1163 Bünde/Westfalen. Palaeo ntographica A, 124, 116-150. [13] Fichter, J. (1999) Die fossile Seekuh von Schauenberg- Hoof, Kreis Kassel. Paläontologische Denkmäler in Hes- sen, 8, 1-14. [14] Diedrich, C. (2008) The food of the miosiren Anomo- therium langenwieschei (Siegfried)—indirect proof of seaweed or seagrass by xenomorphic oyster fixation structures in the Upper Oligocene (Neogene) of the Do- berg, Bünde (NW Germany) and comparisons to modern Dugong dugon (Müller) feeding strategies. Senckenber- giana maritima, 38, 59-73. http://dx.doi.org/10.1007/BF03043869 [15] Zalmout, I.S., Ul-Haq, M. and Gingerich, P.D. (2003) New species of Protosiren (Mammalia, Sirenia) from the early Middle Eocene of Balochistan (Pakistan). Contribu- tions from the Museum of Paleontology, University of Michigan, 31, 79-87. [16] Abel, O. (1904) Die Sirenen der mediterranen Tertiärbil- dungen Östereichs. Abhandlungen der Geologischen Rei- chsanstalt Wien, 19, 1-223. [17] Gingerich, P.D., Domning, D.P., Blane, C.E. and Uhen, M.D. (1994) Cranial morphology of Protosiren fraasi (Mammalia, Sirenia) from the middle Eocene of Egypt. Contributions from the Museum of Paleontology, Univer- sity of Michigan, 29, 41-67. [18] Gingerich, P.D., Arif, M., Bhatti, M.A., Raza, H.A. and Raza, S.M. (1995) Protosiren and Babiacetus (Mammalia, Sirenia and Cetacea) from the middle Eocene Drazinda Formation, Sulaiman Range, Punjab (Pakistan). Con- tributions from the Museum of Paleontology, University of Michigan, 29, 331-357. [19] Domning, D.P. and Gingerich, P.D. (1994) Protosiren smithae, new species (Mammalian, Sirenia) from the Late Middle Eocene of Wa di Hitan, Egypt. Contributions from the Museum of Paleontology of the University of Michi- gan, 29, 69-87. [20] Bajpai, S., Thewissen, J.G.M., Kapur, V.V., Tiwari, B.N. and Sahni, A. (2006) Eocene and Oligocene sirenians (Mammalia) from Kachchh, India. Journal of Vertebrate Paleontology, 26, 400-410. http://dx.doi.org/10.1671/0272-4634(2006)26[400:EAOS MF]2.0.CO;2 [21] Abel, O. (1912) Die Eozänen Sirenen der Mittelmeer- region, Erster Teil: Der Schädel von Eotherium aegy- ptiacum. Palaeontographica, 59, 289-360. [22] Richard, M. (1946) Contribution à l’étude du Bassin d’A- quitaine. Les gisements de mammif`eres tertiaires. Mé- moires de la Société géologique de France N.S., 52, 1- 380. [23] Kordos, L. (1981) Some complements to the knowledge of a Middle Eocene Sirenia, Sirenavus hungaricus Kret- zoi, 1941. Fragmenta Mineralogica et Palaeontologica, 10, 75-78. [24] Kordos, L. (2002) Eocene seacows (Sirenia, Mammalia) from Hungary. Fragmenta Palaeontologica Hungarica, 20, 43-48. [25] Kretzoi, M. (1941) Sirenavus hungaricus nov. g., nov. sp., ein neuer Prorastomide aus dem Mittleleozän (Lutetium) von Felsögalla in Ungarn. Annales Musei Nationalis Hungarici, Pars Mineralogica, Geologica et Palaeon- tologica, 34, 146-156. [26] Astibia, H. , Ba rdet, N., Pereda-Suberbiola, X., Payros, A., De Buffr, V., Elorza, J., Tesquella, J., Berreteaga, A. and Badiola, A. (2010) New fossils of Sirenia from the Mid- dle Eocene of Navarre (Western Pyrenees): The oldest West European seacow record. Geological Magazine, 147, 665-673. http://dx.doi.org/10.1017/S0016756810000130 [27] Bizzotto, B. (2005) La struttura cranica di Prototherium intermedium (Mammalia: Sirenia) dell ’Eocene superiore Veneto.Nuovi contributi alla sua anatomia e sistematica. Lavori della Societ`a Veneziana di Scienze Naturali, 30, 107-125. [28] Bizzotto, B. (2005) La struttura cranica di Prototherium intermedium (Mammalia: Sirenia) dell ’Eocene superiore Veneto.Nuovi contributi alla sua anatomia e sistematica. Lavori della Societ`a Veneziana di Scienze Naturali, 30, 107-125. [29] Kordos, L. (1977) A New Upper Eocene Sirenian Paralitherium tarkanyense n,g,n,sp, from Felsotarkany, NE Hungary. Magyar Allami Foldtani Intezet Evi Jelen- tese az Evrol, 1977, 349-367. [30] Sahni, A., Kumar, K. and Tiwari, B.N. (1980) Lower Eocene Marine Mammal (Sirenia) from Dharampur, Simla Himalayas, H. P. Current Science, 49, 270-271. [31] Sahni, A. and Kumar, K. (1980) Lower Eocene Sirenia, Ishatherium subathuensis, gen, et sp. nov. from the Type Area, Subathu Formation, Subathu, Simla Himalayas. Journal of the Palaeontological Society of India, 23-24, 132-135. [32] Fuchs, H. (1970) Schädelfragment einer Sirene aus dem Eozän von Cluj, SR Rumänien. Geologica, 19, 1185- 1191. [33] Gingerich, P.D. (1992) Marine mammals (Cetacea and Sirenia) from the Eocene of Gebel Mokattam and Fayum, Egypt: Stratigraphy, age, and paleoenvironments. Univer- sity of Michigan Papers on Paleontology, 30, 1-84. [34] Zdansky, O. (1938) Eotherium majus sp. n, eine neue Sirene aus dem Mitteleozän von Ägypten. Palaeobio- logica, 6, 429-434. [35] Domning, D.P., Gary, S., Morgan and Ray, C.E. (1982) North American Eocene seacows (Mammalia: Sirenia). Smithsonian contributions to Paleobiology, 52, 1-69. [36] Zalmout, I. S., Haq, M. and Gingerich, P.D. (2001) Earli- est Tethyan seacow from the Lutetian Eocene of Pakistan. Journal of Ve rtebrate Paleontology, 2, 117. [37] Sickenberg, O. (1934) Beträge zur Kenntnis tertiärer Sirenen. I. Die Eozänen Sirenen des Mittelmeergebietes. II. Die Sirenen des Belgischen Tertiärs. Mémoires de la Musée Royal d’Histoire Naturelle de Belgique, 63, 1-352. [38] Siler, W.L. (1964) A middle Eocene Sirenian in Alabama. Journal of Paleontology, 38, 1108-1109. [39] Bufferénil, V. De, Astiba, H., Pereda-Suberbiola, X., Berreteagea, A. and Bardet, N. (2008) Variation in bone histology of middle Eocene sirenians from western Europe. Geodiversitas, 30, 425-532. [40] Bajpai, S., Domning, D.P. and Mishra, V.P. (2009) A new Copyright © 2013 SciRes. OPEN A CCESS  C. G. Diedrich / Natural Scienc e 5 (2013) 1154-1164 Copyright © 2013 SciRes. OPEN A CCESS 1164 middle Eocene sirenian (Mammalia, Protosirenidae) from India. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 252, 257-67. http://dx.doi.org/10.1127/0077-7749/2009/0252-0257 [41] Sagne, C. (2001) Halitherium taulannense, nouveau siré- nien (Sirenia, Mammalia) de l’Éocene supérieur prove- nant du domaine Nord-Téthysien (Alpes-de-Haute-Pro- vence, France). Comptes Rendus de l’Académie des Sci- ences Paris, Sciences de la Terre et des planets, 333, 471- 476. [42] Sauvage, R.J.G. (1971) Review of fossil mammals of Libya. In: Gray, C., Ed., Symposium on the Geology of Libya, University of Libya, 215-225. [43] Samonds, K., Zalmout, I.S. and Krause, D.W. (2005) New Sirenian fossils from the late Eocene of Madagascar. Journal of Ve rtebrate Paleontology, 25, 108A. [44] Fischer, K. and Krumbiegel, G. (1982) Halitherium schin- zi Kaup 1838 (Sirenia, Mammalia) aus dem marinen Mit- teloligozän des Weißelsterbeckens (Bezirk Leipzig, DDR). Halleshes Jahrbuch für Geowissenschaften, 7, 73-95. [45] Diedrich, C. (2011) Palaeoecology of the macrofauna of the Upper Oligocene (Palaeogene) littoral to shallow sub- tidal of Astrup near Osnabrück (NW Germany). Central European Journal of Geosciences, 4, 163-187. http://dx.doi.org/10.2478/s13533-011-0051-1 [46] Pervesler, P., Roetzel, R. and Steininger, F. (1995) Tapho- nomie der Sirenen in den marinen Flachwasserablagerun- gen (Burgschleinitz-Formation, Eggenburgium, Unter- miozän) der Gemeindesandgrube Kühnring (Niederöster- reich). Jahrbuch der Geologischen Bundesandesanstalt, 138, 89-121. [47] Hautier, L., Sarr, R., Tabuce, F., Lihoreau, S., Adnet, S., Domning, D.-P., Samb, M. and Hameh, P.M. (2012) First prorastomid sirenian from Senegal (western Africa) and the Old World origin of sea cows. Journal of Vertebrate Paleontology, 32, 1218-1222. http://dx.doi.org/10.1080/02724634.2012.687421 [48] Gheerbrant, E, Domning, D.P. and Tassy, P. (2005) Pae- nungulata (Sirenia, Proboscidea, Hyracoidea, and rela- tives). In: Rose, K.D. and Archibald, J.D., Eds., The Rise of Placental Mammals: Origin and Relationships of the Major Extant Clades, Baltimore John Hopkins University Press, 84-105.
|