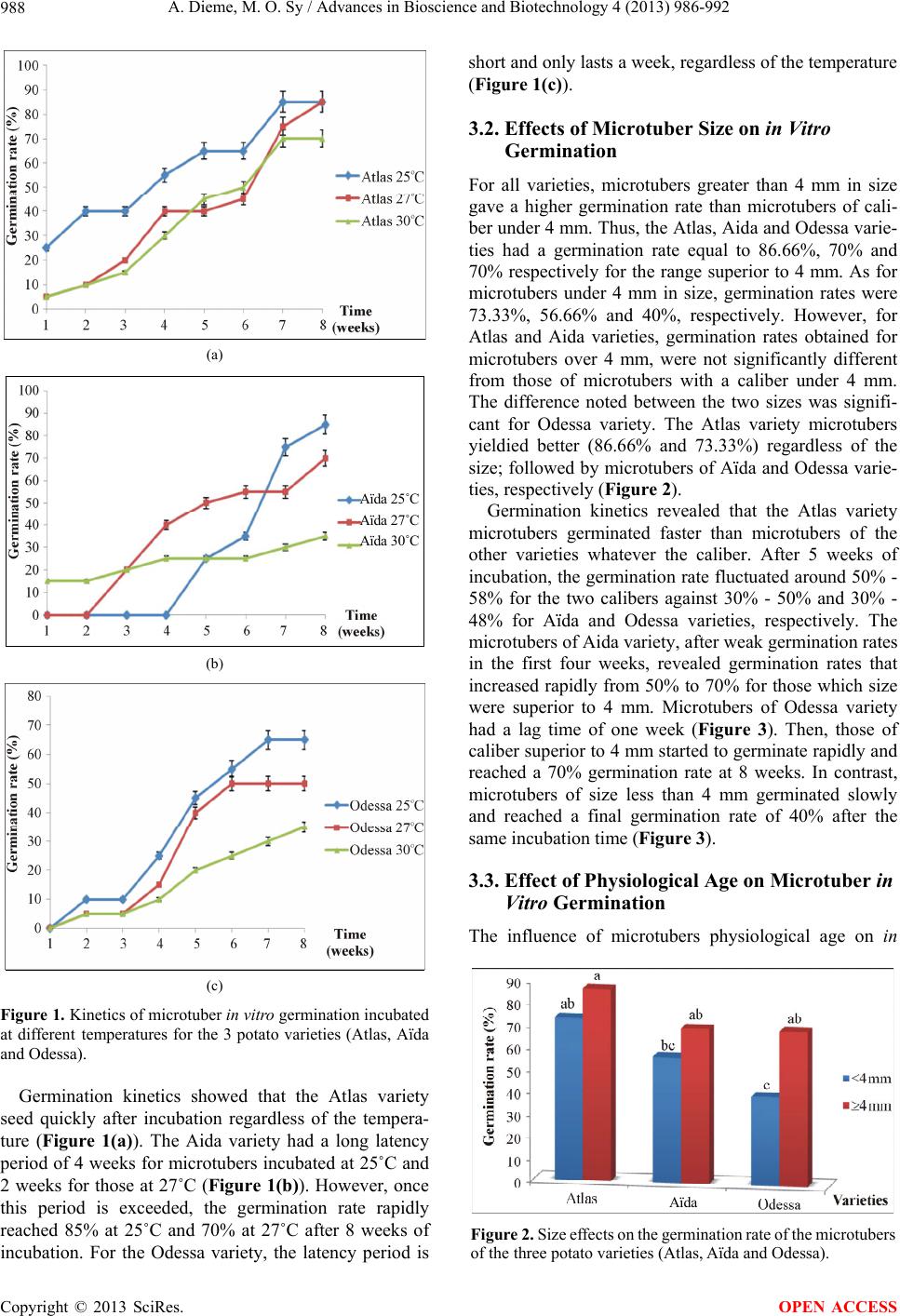
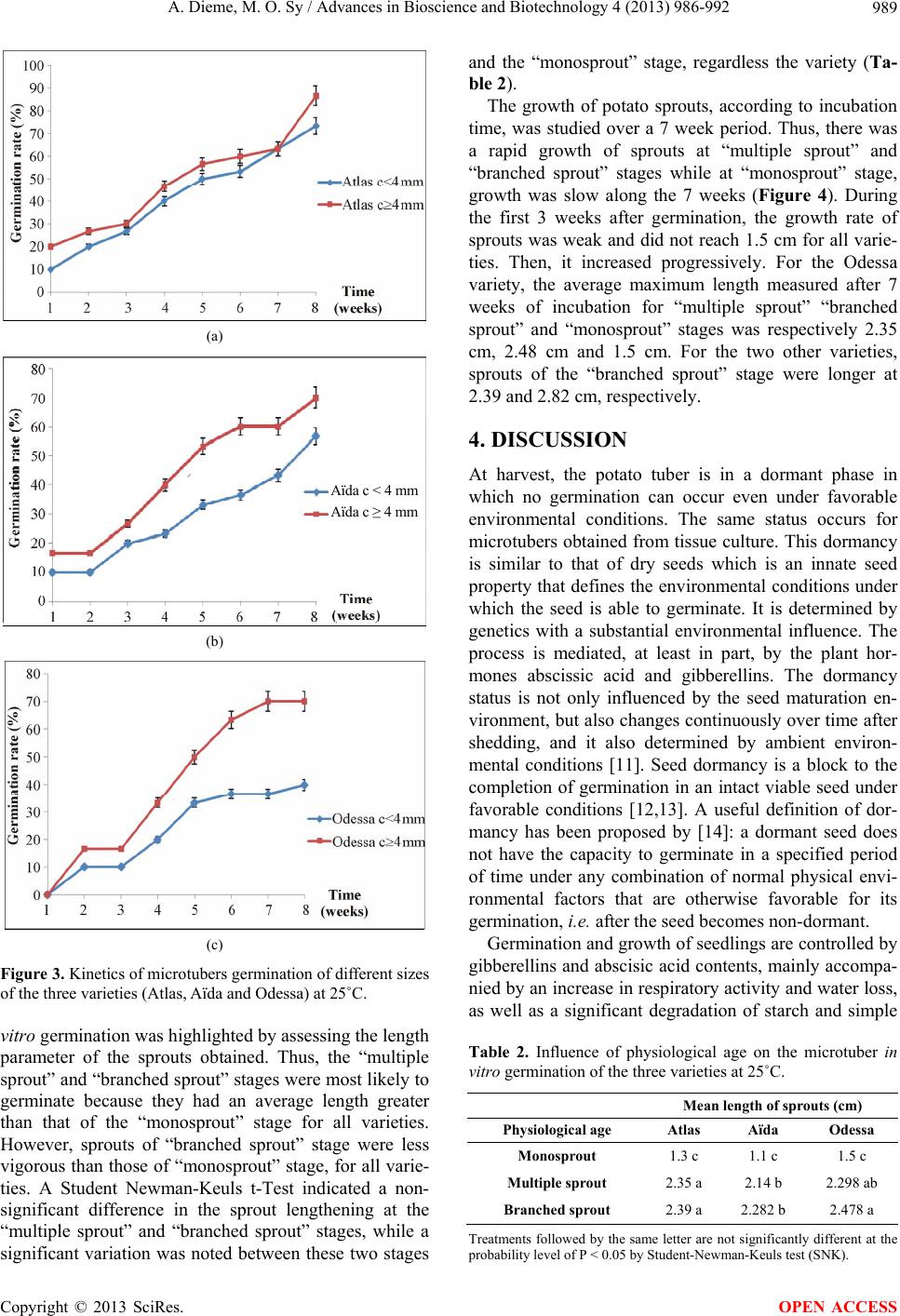
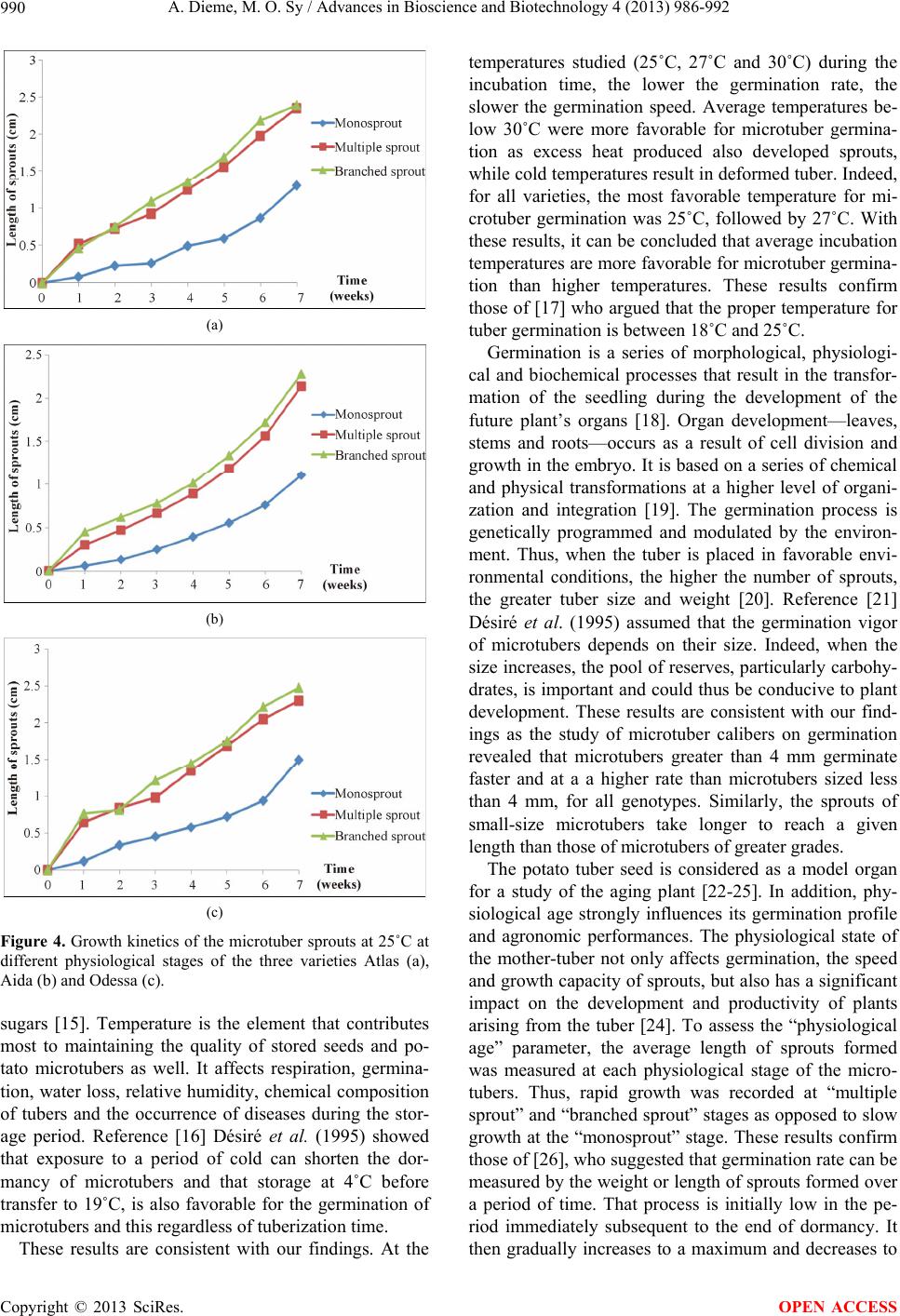
A. Dieme, M. O. Sy / Advances in Bioscience and Biotechnology 4 (2013) 986-992 991
zero when sprouts start forming new tubers again. Ref-
erences [27,28] also demonstrated that as potato tubers
get physiologically older, germination occurs propor-
tionally faster. When physiologically young, microtubers
are either dormant or in a phase of insignificant germina-
tion. Then, as they get older, their germination vigor in-
creases and germination is therefore accelerated. In the
same manner, reference [26] confirmed that during the
dormant phase, the inability to grow is not necessarily a
specific characteristic of buds, but may result in a tem-
porary inability of the tuber to provide some metabolites
necessary for growth. These results are consistent with
our findings because with the Odessa variety, a latency
period occured during a week. This can be explained by
the fact that microtubers were still dormant when put to
germinate. At the end of dormancy, metabolites are re-
leased, making it possible for sprouts to grow from the
tuber. This release is gradual and as, the tuber gets phy-
siologically older, allows a more rapid growth of an in-
creasing number of sprouts.
Microtubers, at “multiple and branched sprout” stages
are more likely to germinate than sprouts of the “mono-
sprout” stage, with an average maximum sprout length of
2.35 cm, 2.48 cm and 1.5 cm, respectively. These results
confirm those of [29] and [24] explaining that the phy-
siological state of a tuber, at a given time, determines
vegetative growth. The “monosprout” stage is character-
ized by an apical dominance, the “multiple sprout” stage
by multiple germinations and rapid growth of sprouts
and the “branched sprouts” stage by stunted sprouts [30].
Experiments on the influence of temperature, size and
physiological age on the in vitro germination of micro-
tubers have enable us to conclude that the average tem-
perature of 25˚C is the most favorable for the germina-
tion of microtubers at a better and faster rate, irrespective
of the variety. Microtubers with a grade greater than 4
mm, germinated more quickly, and at a higher rate, than
those of a smaller caliber for all genotypes. Physiological
age influenced microtuber germination. In fact, the mean
length of sprouts was greater at “multiple sprout” and
“branched sprout” stages than at a “monosprout” stage.
Thus, it is necessary to plant microtubers at the “multiple
sprout” stage to optimize their performance for the vege-
tative growth of plants and minituber production.
5. ACKNOWLEDGEMENTS
Authors are grateful to Mrs. Aminata Sow for correcting the English
version of the manuscript.
REFERENCES
[1] Charles, G., Rossignol, L. and Rossignol, M. (1995) Mise
au point d’un modèle de développement et de tubérisation
contrôlés et synchrones chez lapomme de terre cultivée in
vitro. Acta Botanica Gallica, 142, 289-300.
http://dx.doi.org/10.1080/12538078.1995.10515245
[2] Désiré, S., Couillerot, J.P. and Vasseur J. (1995) Germi-
nation en serre des microtubercules de pomme de terre
(Solanum tuberosum L.) produit in vitro: Influence du
diamètre, de la densité de la plantation et de l’âge des
microtubercules sur le rendement. Acta Botanica Gallica,
142, 379-387.
http://dx.doi.org/10.1080/12538078.1995.10515258
[3] Coleman, W.K., Donnelly, D.J. and Coleman, S.E. (2001)
Potato microtubers as research tools: A review. American
Journal of Potato Research, 78, 47-55.
http://dx.doi.org/10.1007/BF02874824
[4] Estrada, R., Tovar, P. and Dodds, J.H. (1986) Induction
of in vitro tubers in a broad range of potato genotypes.
Plant Cell Tissue and Organ Culture, 3, 3-10.
http://dx.doi.org/10.1007/BF00043915
[5] Slimmon, T., Machado, V.S. and Coffin, R. (1989) The
effect of light on in vitro microtuberization of potato cul-
tivars. American Potato Journal, 66, 843-848.
http://dx.doi.org/10.1007/BF02853965
[6] Wiersema, S.G., Cabello, R., Tovar, P. and Dodds, J.H.
(1987) Rapid seed multiplication by planting into beds
microtubers and in vitro plants. Potato Research, 30, 200-
214. http://dx.doi.org/10.1007/BF02357690
[7] Leclerc, Y., Donnelly, D.J. and Seabrook, J.E.A. (1994)
Microtuberization of layered shoots and cuttings of potato:
The influence of growth regulators and incubation peri-
ods. Plant Cell Tissue and Organ Culture, 37, 113-120.
http://dx.doi.org/10.1007/BF00043604
[8] Dieme, A., Sagna, M. and Sy, M.O. (2011) Influence of
hormonal treatments and of sucrose on the microtuberiza-
tion of three potato varieties (Solanum tuberosum L.)
adapted to agroclimatic conditions in Senegal). Interna-
tional Journal of Plant, Animal and Environmental Sci-
ences, 1, 69-77.
[9] Murashige, T. and Skoog, F. (1962) A revised medium
for rapid growth and bioassays with tobacco tissue cul-
ture. Physiologa Plantarum, 15, 473-497.
http://dx.doi.org/10.1111/j.1399-3054.1962.tb08052.x
[10] Diémé, A., Sambe, M.A.N., Agbangba, E.C. and Sy, M.O.
(2013) Residual effects of sucrose and hormonal treat-
ments of the tuberization medium on in vitro germination
of potato (Solanum tuberosum L.) microtubers. American
Journal of Plant Sciences, 4, 1872-1878.
http://dx.doi.org/10.4236/ajps.2013.49230
[11] Finch-Savage, W.E. and Leubner-Metzger, G. (2006)
Seed dormancy and the control of germination. New Phy-
tologist, 171, 501-523.
http://dx.doi.org/10.1111/j.1469-8137.2006.01787.x
[12] Bewley, J.D. (1997) Seed germination and dormancy.
Plant Cell, 9, 1055-1066.
http://dx.doi.org/10.1105/tpc.9.7.1055
[13] Li, B.L. and Foley, M.E. (1997) Genetic and molecular
control of seed dormancy. Trends in Plant Science, 2,
384-389.
http://dx.doi.org/10.1016/S1360-1385(97)90053-4
[14] Baskin, J.M. and Baskin, C.C. (2004) A classification
Copyright © 2013 SciRes. OPEN ACCESS