A. Papadopoulos / Open Journal of Animal Sciences 3 (2013) 314-320 319
that these optimal efficiencies are homogenous functions
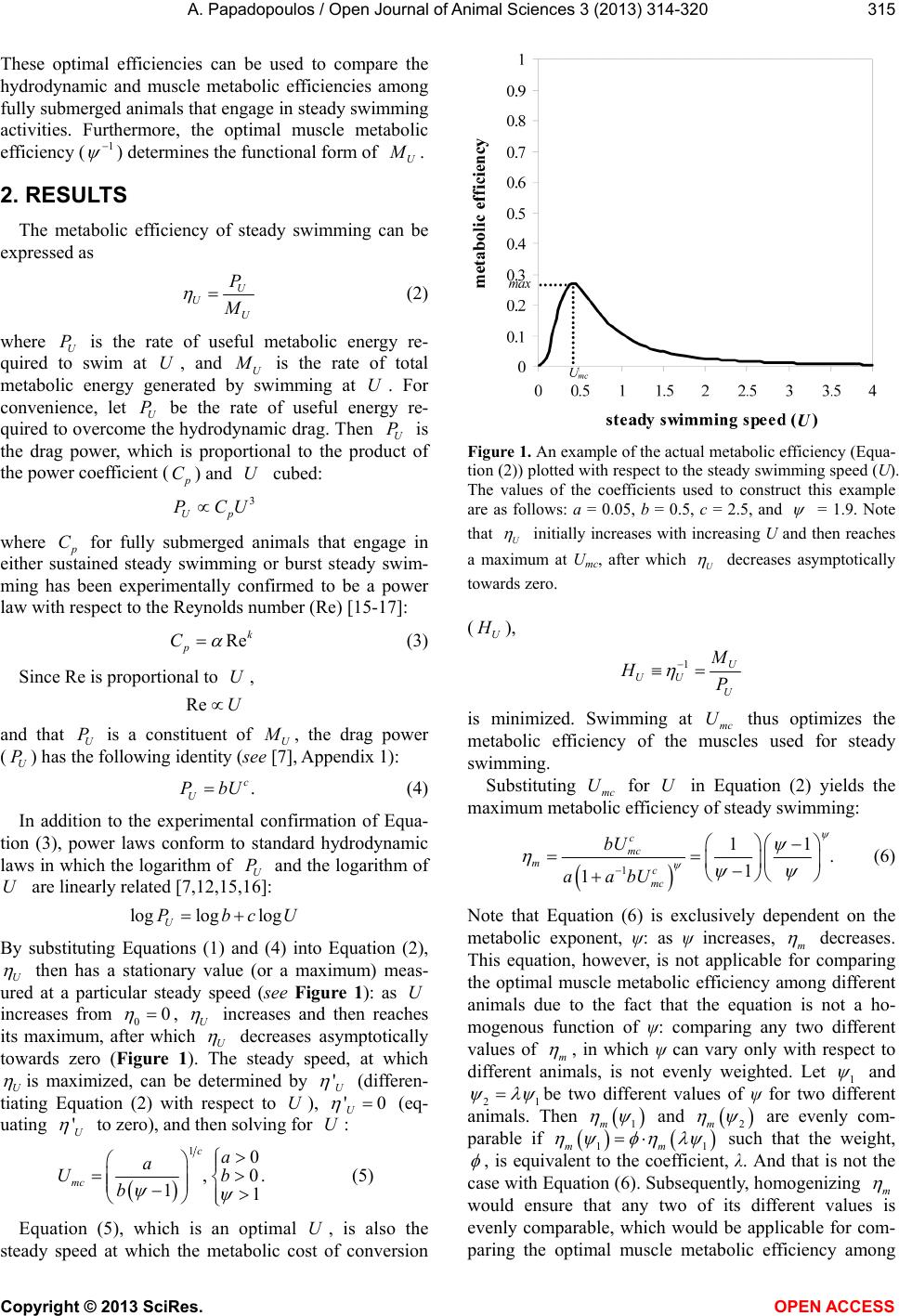
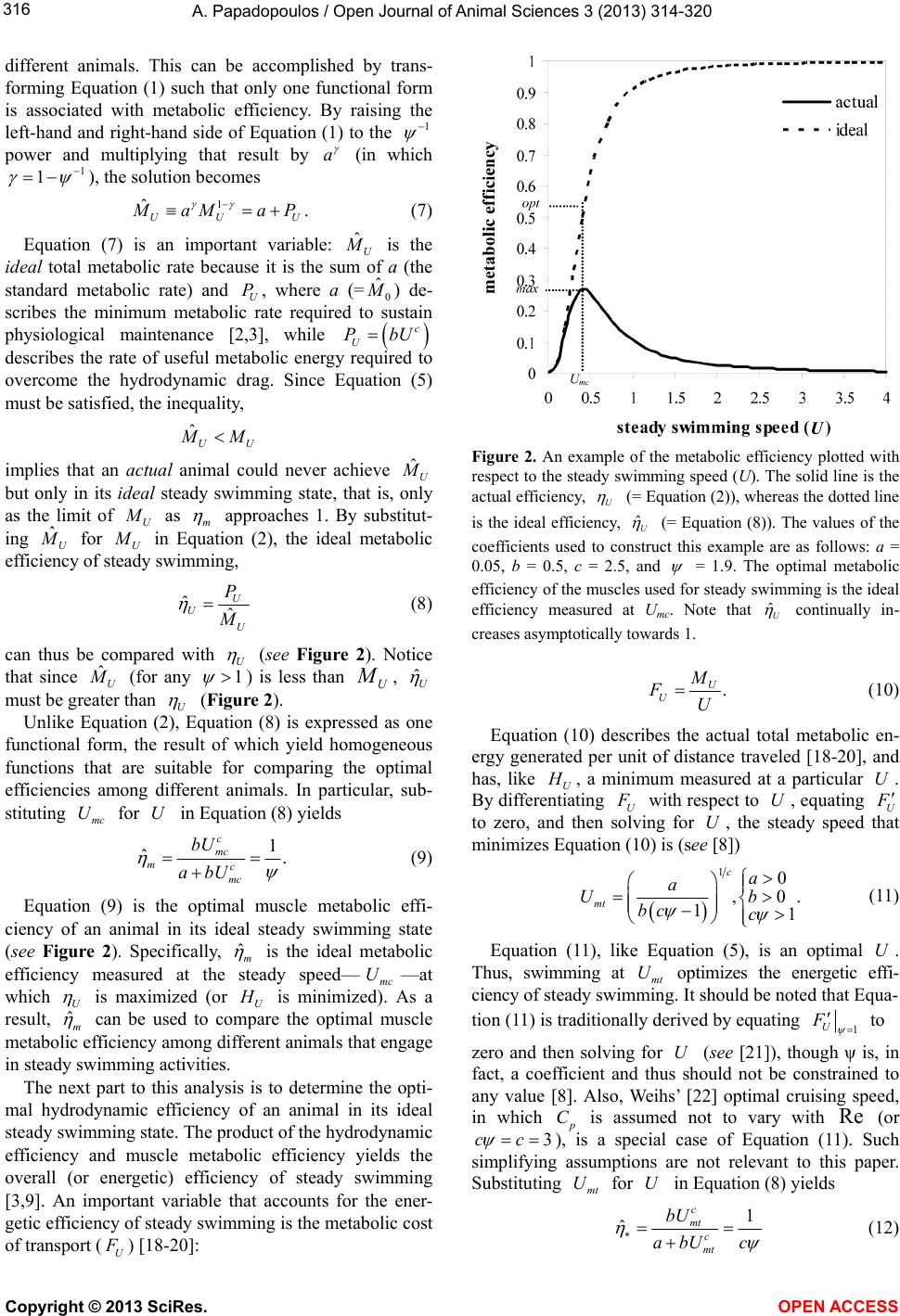
of c and
: ˆh
and ˆm
are, in fact, homogenous due
to the transformation of Equation (2) into Equation (8),
which is the ideal form of U
. Furthermore, since ˆh
and ˆm
are inverses of the exponents in Equation (1),
the different models of U
exclusively arise from the
different values of ˆh
and ˆm
.
5. ACKNOWLEDGEMENTS
I thank the reviewers for providing comments that improved this
manuscript. I was employed at Texas Tech University, where I inde-
pendently wrote this manuscript and derived all of the results. Support
for this work was provided in part by National Science Foundation
award DEB-0616942 to Sean H. Rice.
REFERENCES
[1] Ivlev, V.S. (1960) Active metabolic intensity in salmon
fry (Salmo salar L.) at various rates of activity. Salmon
and Trout Comm, Int Counc Explor Sea, Copenhagen,
213, 1-16.
[2] Brett, J.R. (1964) The respiratory metabolism and swim-
ming performance of young sockeye salmon. Journal of
the Fisheries Research Board of Canada, 21, 1183-1226.
http://dx.doi.org/10.1139/f64-103
[3] Webb, P.W. (1974) Hydrodynamics and energetics of fish
propulsion. Bulletin of the Fisheries Research Board of
Canada, 190, 109-119.
[4] O’Dor, R.K. and Webber, D.M. (1991) Invertebrate ath-
letes: Trade-offs between transport efficiency and power
density in cephalopod evolution. The Journal of Experi-
mental Biology, 160, 93-112.
[5] Hind, A.T. and Gurney, W.S.C. (1997) The metabolic cost
of swimming in marine homeotherms. The Journal of
Experimental Biology, 200, 531-542.
[6] Fish, F.E. (2000) Biomechanics and energetics in aquatic
and semiaquatic mammals: Platypus to whale. Physiolo-
gical and Biochemical Zoology, 73, 683-698.
http://dx.doi.org/10.1086/318108
[7] Papadopoulos, A. (2008) On the hydrodynamics-based
power-law function and its application in fish swimming
energetics. Transactions of the American Fisheries Soci-
ety, 137, 997-1006. http://dx.doi.org/10.1577/T07-116.1
[8] Papadopoulos, A. (2009) Hydrodynamics-based functional
forms of activity metabolism: A case for the power-law
polynomial function in animal swimming energetics. PLoS
ONE, 4, e4852.
http://dx.doi.org/10.1371/journal.pone.0004852
[9] Webb, P.W. (1993) Swimming. In: Evens, D.H., Ed., The
Physiology of Fishes, CRC Press, Boca Raton, 47-73.
[10] Wardle, C.S., Soofiani, N.M., O’Neill, F.G., Glass, C.W.
and Johnstone, A.D.F. (1996) Measurements of aerobic
metabolism of a school of horse mackerel at different
swimming speeds. Journal of Fish Biology, 49, 854-862.
h ttp :// d x. doi . org/1 0 .1111 / j. 1095-8649.1996.tb00084.x
[11] Pettersson, L.B. and Hedenström, A. (2000) Energetics,
cost reduction and functional consequences of fish mor-
phology. Proceedings of the Royal Society B, 267, 759-
764. http://dx.doi.org/10.1098/rspb.2000.1068
[12] Korsmeyer, K.E., Steffensen, J.F. and Herskin, J. (2002)
Energetics of median and paired fin swimming, body and
caudal fin swimming, and gait transition in parrotfish
(Scarus schlegeli) and triggerfish (Rhinecanthus aculea-
tus). The Journal of Experimental Biology, 20 5, 1253-
1263.
[13] Behrens, J.W., Praebel, K. and Steffensen, J.F. (2006)
Swimming energetics of the Barents Sea capelin (Mallo-
tus villosus) during the spawning migration period. Jour-
nal of Experimental Marine Biology and Ecology, 331,
208-216. http://dx.doi.org/10.1016/j.jembe.2005.10.012
[14] Ohlberger, J., Staaks, G. and Holker, F. (2006) Swimming
efficiency and the influence of morphology on swimming
costs in fishes. Journal of Comparative Physiology B,
176, 17-25.
http://dx.doi.org/10.1007/s00360-005-0024-0
[15] Wu, T.Y. (1977) Introduction to the scaling of aquatic
animal locomotion. In: Pedley, T.J., Ed., Scale Effects in
Animal Locomotion, Academic Press, New York.
[16] Wu, T.Y. and Yates, G.T. (1978) A comparative mech-
anophysiological study of fish locomotion with implica-
tions for tuna-like swimming mode. In: Sharp, G.D. and
Dizon, A.E., Eds., Physiological Ecology of Tuna, Aca-
demic Press, New York.
[17] Fung, Y.C. (1990) Biomechanics: Motion, flow, stress,
and growth. Springer-Verlag, New York.
[18] Tucker, V.A. (1970) Energetic cost of locomotion in ani-
mals. Comparative Biochemistry and Physiology, 34, 841-
846. http://dx.doi.org/10.1016/0010-406X(70)91006-6
[19] Tucker, V.A. (1975) The energetic cost of moving about.
American Scientist, 63, 413-419.
[20] van Ginneken, V., Antonissen, E., Müller, U.K., Booms,
R., Eding, E., Verreth, J. and van den Thillart, G. (2005)
Eel migration to the Sargasso: Remarkably high swim-
ming efficiency and low energy costs. The Journal of Ex-
perimental Biology, 208, 1329-1335.
http://dx.doi.org/10.1242/jeb.01524
[21] Videler, J.J. and Nolet, B.A. (1990) Costs of swimming
measured at optimum speed: Scale effects, differences
between swimming styles, taxonomic groups, and sub-
merged and surface swimming. Comparative Biochemis-
try and Physiology, 97A, 91-99.
http://dx.doi.org/10.1016/0300-9629(90)90155-L
[22] Weihs, D. (1973) Optimal fish cruising speed. Nature,
245, 48-50. http://dx.doi.org/10.1038/245048a0
[23] Plaut, I. (2001) Critical swimming speed: Its ecological
relevance. Comparative Biochemistry and Physiology,
131A, 41-50.
[24] Blake, R.W. (2004) Fish functional design and swimming
performance. Journal of Fish Biology, 65, 1193-1222.
h ttp :// d x. doi . org/1 0 .1111 / j. 0022-1112.2004.00568.x
[25] Langerhans, R.B. (2009) Trade-off between steady and
unsteady swimming underlies predator-driven divergence
in Gambusia affinis. Journal of Evolutionary Biology, 22,
Copyright © 2013 SciRes. OPEN ACCESS