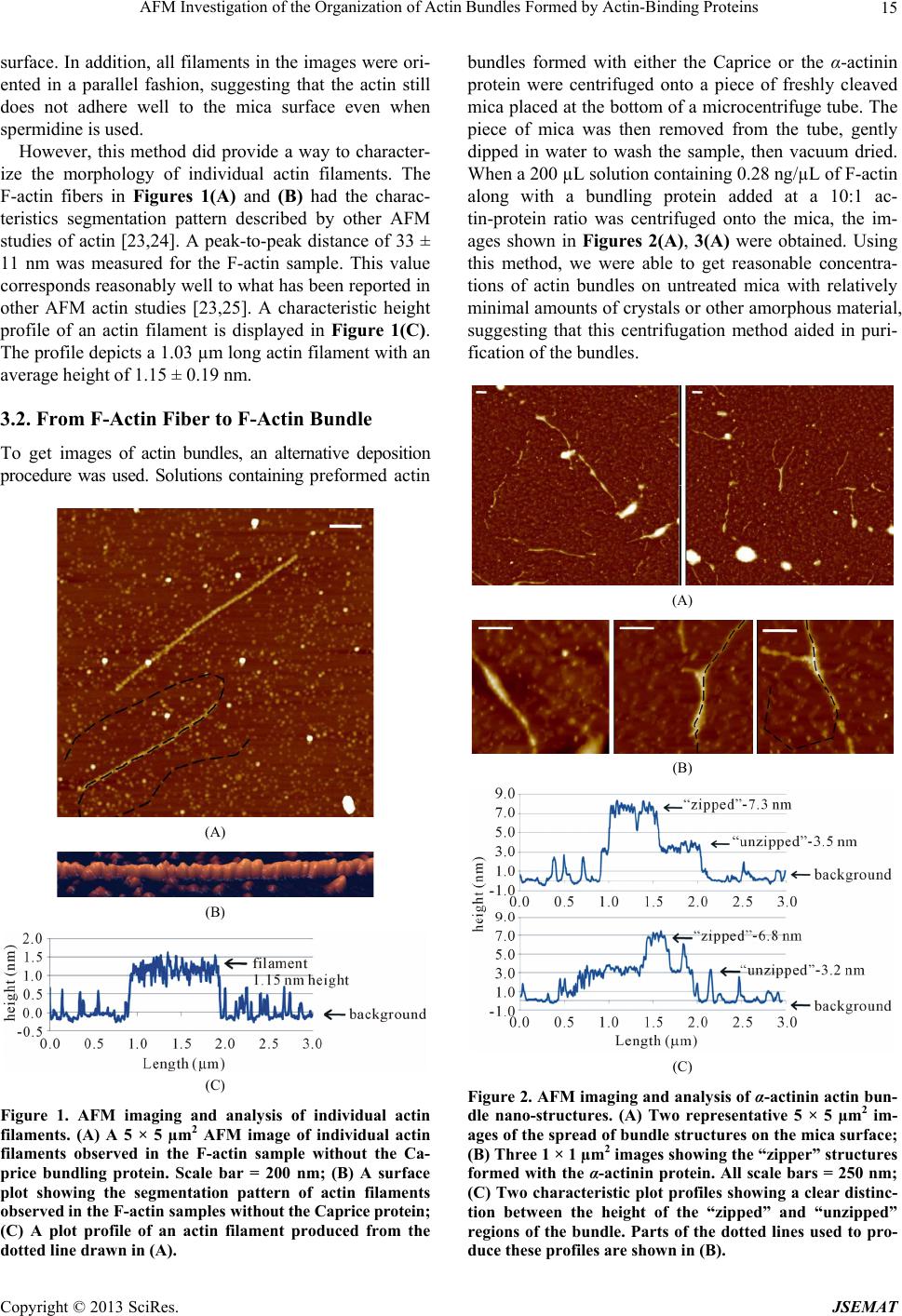
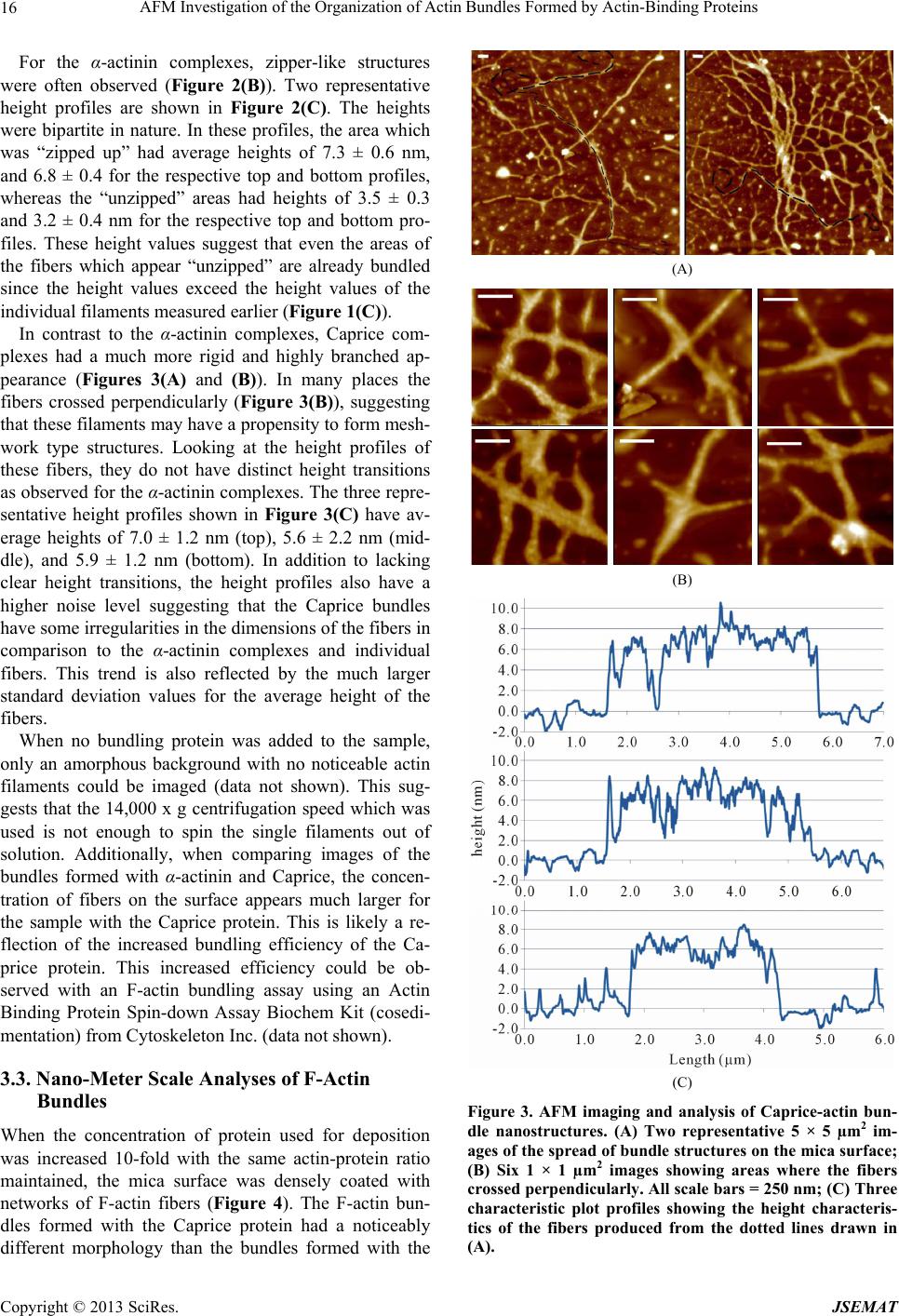
AFM Investigation of the Organization of Actin Bundles Formed by Actin-Binding Proteins
Copyright © 2013 SciRes. JSEMAT
18
Japan. M.K. is supported by a Grant-in-Aid for Scientific
Research on Innovative Areas “Spying minority in bio-
logical phenomen a” (#24115512 ) from MEXT Jap an and
a Cross-Disciplinary Research Promotion Project from
iCeMS, Kyoto University, Japan.
REFERENCES
[1] P. Hansma, V. Elings, O. Marti and C. Bracker, “Scan-
ning Tunneling Microscopy and Atomic Force Micro-
scopy: Application to Biology and Technology,” Science,
Vol. 242, No. 4876, 1988, pp. 209-216.
http://dx.doi.org/10.1126/science.3051380
[2] G. Binnig, C. Quate and C. Gerber, “Atomic Force Mi-
croscope,” Physical Review Letters, Vol. 56, No. 9, 1986,
pp. 930-933.
http://dx.doi.org/10.1103/PhysRevLett.56.930
[3] V. Morris, A. Gunning, and A. Kirby, “Atomic Force
Microscopy for Biologists,” Imperial College Press, Lon-
don, 1999. http://dx.doi.org/10.1142/p173
[4] B. Bhushan, “Scanning Probe Microscopy in Nanosci-
ence and Nanotechnology,” Springer-Verlag Berlin Hei-
delberg, Berlin, 2010.
[5] B. Bhushan, “Scanning Probe Microscopy in Nanosci-
ence and Nanotechnology,” Springer-Verlag Berlin Hei-
delberg, Berlin, 2011.
[6] K. Takeyasu, “Atomic Force Microscopy in Nano-Biol-
ogy,” Pan Stanford Publishing Co., Singapore City, 2013.
[7] Q. Zhong and D. Inniss, “Fractured Polymer/Silica Fiber
Surface Studied by Tapping Mode Atomic Force Mi-
croscopy,” Surface Science Letters, Vol. 290, No. 1-2,
1993, pp. L688-L692.
[8] K. Hohmura, Y. Itokazu, S. Yoshimura, G. Mizuguchi, Y.
Masamura, K. Takeyasu, et al., “Atomic Force Micros-
copy with Carbon Nanotube Probe Resolves the Subunit
Organization of Protein Complexes,” Journal of Electron
Microscopy (Tokyo), Vol. 49, No. 3, 2000, pp. 415-421.
http://dx.doi.org/10.1093/oxfordjournals.jmicro.a023823
[9] J. Gilmore, Y. Suzuki, G. Tamulaitis, V. Siksnys, K. Ta-
keyasu and Y. Lyubchenko, “Single-Molecule Dynamics
of the DNA-EcoRII Protein Complexes Revealed with
High-Speed Atomic Force Microscopy,” Biochemistry,
Vol. 48, No. 44, 2009, pp. 10492-10498.
http://dx.doi.org/10.1021/bi9010368
[10] Y. Suzuki, Y. Higuchi, K. Hizume, M. Yokokawa, S.
Yoshimura, K. Yoshikawa, et al., “Molecular Dynamics
of DNA and Nucleosomes in Solution Studied by Fast-
Scanning Atomic Force Microscopy,” Ultramicroscopy,
Vol. 110, No. 6, 2010, pp. 682-688.
http://dx.doi.org/10.1016/j.ultramic.2010.02.032
[11] Y. Suzuki, J. Gilmore, S. Yoshimura, R. Henderson, Y.
Lyubchenko and K. Takeyasu, “Visual Analysis of Con-
certed Cleavage by Type IIF Restriction Enzyme SfiI in
Subsecond Time Region,” Biophysical Journal, Vol. 101,
No. 12, 2011, pp. 2992-2998.
http://dx.doi.org/10.1016/j.bpj.2011.09.064
[12] H. Sanchez, Y. Suzuki, M. Yokokawa, K. Takeyasu and
C. Wyman, “Protein-DNA Interactions in High Speed
AFM: Single Molecule Diffusion Analysis of Human
RAD54,” Integrative Biology (Camb), Vol. 3, No. 11,
2011, pp. 1127-1134.
http://dx.doi.org/10.1039/c1ib00039j
[13] M. Yokokawa, C. Wada, T. Ando, N. Sakai, A. Yagi, S.
Yoshimura, et al., “Fast-Scanning Atomic Force Micros-
copy Reveals the ATP/ADP-Dependent Conformational
Changes of GroEL,” The EMBO Journal, Vol. 25, No. 19,
2006, pp. 4567-4576.
http://dx.doi.org/10.1038/sj.emboj.7601326
[14] N. Kodera, D. Yamamoto, R. Ishikawa and T. Ando,
“Video Imaging of Walking Myosin V by High-Speed
Atomic Force Microscopy,” Nature, Vol. 468, No. 7320,
2010, pp. 72-76. http://dx.doi.org/10.1038/nature09450
[15] M. Yokokawa and K. Takeyasu, “Motion of the Ca2+
Pump Captured,” FEBS Journal, Vol. 278, No. 17, 2011,
pp. 3025-3031.
http://dx.doi.org/10.1111/j.1742-4658.2011.08222.x
[16] M. Shibata, T. Uchihashi, H. Yamashita, H. Kandori and
T. Ando, “Structural Changes in Bacteriorhodopsin in
Response to Alternate Illumination Observed by High-
Speed Atomic Force Microscopy,” Angewandte Chemie
International Edition, Vol. 50, No. 19, 2011, pp. 4410-
4413. http://dx.doi.org/10.1002/anie.201007544
[17] Y. Suzuki, T. Goetze, D. Stroebel, D. Balasuriya, S. Yo-
shimura, R. Henderson, et al., “Visualization of Structural
Changes Accompanying Activation of N-Methyl-D-As-
partate (NMDA) Receptors Using Fast-Scan Atomic Force
Microscopy Imaging,” The Journal of Biological Chem-
istry, Vol. 288, No. 2, 2013, pp. 778-784.
http://dx.doi.org/10.1074/jbc.M112.422311
[18] N. Crampton, M. Yokokawa, D. Dryden, J. Edwardson, D.
Rao, K. Takeyasu, et al., “Fast-Scan Atomic Force Mi-
croscopy Reveals That the Type III Restriction Enzyme
EcoP15I is Capable of DNA Translocation and Looping,”
Proceedings of the National Academy of Sciences of the
United States of America, Vol. 104, No. 31, 2007, pp.
12755-12760. http://dx.doi.org/10.1073/pnas.0700483104
[19] Y. Hirano, K. Ishii, M. Kumeta, K. Furukawa, K. Take-
yasu and T. Horigome, “Proteomic and Targeted Ana-
lytical Identification of BXDC1 and EBNA1BP2 as Dy-
namic Scaffold Proteins in the Nucleolus,” Genes Cells,
Vol. 14, No. 2, 2009, pp. 155-166.
http://dx.doi.org/10.1111/j.1365-2443.2008.01262.x
[20] M. Kumeta, Y. Hirai, S. Yoshimura, T. Horigome and K.
Takeyasu, “Antibody-Based Analysis Reveals ‘Filamen-
tous vs. Non-Filamentous’ and ‘Cytoplasmic vs. Nuclear’
Crosstalk of Cytoskeletal Proteins,” Experimental Cell Re-
search, in Press.
[21] M. Zhu, F. Settele, S. Kotak, L. Sanchez-Pulido, L. Ehret,
C. Ponting, et al., “MISP Is a Novel Plk1 Substrate Re-
quired for Proper Spindle Orientation and Mitotic Pro-
gression,” The Journal of Cell Biology, Vol. 200, No. 6,
2013, pp. 773-787.
http://dx.doi.org/10.1083/jcb.201207050
[22] K. Zechel and K. Weber, “Actins from Mammals, Bird,
Fish and Slime Mold Characterized by Isoelectric Focus-
ing in Polyacrylamide Gels,” European Journal of Bio-
chemistry, Vol. 89, No. 1, 1978, pp. 105-112.