Paper Menu >>
Journal Menu >>
 Engineering, 2012, 5, 184-188 doi:10.4236/eng.2012.410B048 Published Online October 2012 (http://www.SciRP.org/journal/eng) Copyright © 2012 SciRes. ENG Preparation of Water Soluble Yeast Glucan by Four Kinds of Solubili zing Processes Liping Du, Xuekua ng Zhan g, Chao Wang, Dongguang Xiao Key Laboratory of Industrial Microbiology, Ministry of Education, Tianjin Industrial Microbiology Key Laboratory, College of Bi otechnology Tianjin University of Science and Technology, Tianjin University of Science and Technology, Tianjin 300457, P. R. China Email: dlp123@tust.edu.cn, xdg@tust.edu.cn Received 2012 ABSTRACT (1→3)-β-D-glucan from the inner cell wall of Saccharomyces cerevisiae is considered a member of a class of drugs known as bio- logical response modifiers (BRM). However the glucan was an insoluble polysaccharide, which could be the major barrier to the utilization of glucan. In this case, the insoluble glucan was convent into a soluble form by four kind of solubilizing processes. The yield, solubility, chemistry structure and immunoprophylaxis efficacy of the soluble products were compared. Our date suggest that urea has a significant e ffect on yield, and D MSO has a sign ificant effect o n solubilit y. FT-IR s pectra, 13C NMR spectra an d helix-coil transition analysis demonstrate that the chemistry structure of native and solubilizing glucans have no significant difference. They still have the triple helical structure. The solubility and immunoprophylaxis efficacy assay indicate that the introduction of phosphate group not only enhanced the solubility of glucan, but also improved the survival rate of mice challenged with E. coli. Keywords: Glucan; Solubility; FT-IR Spectra; 13C NMR Spectra; Immunoprophylaxis Efficac y 1. Introduction Glucan is a β-linked polyglucose immune stimulant that can be isolated from the cell wall of Saccha rom yces cerevisi ae [1,2]. It can be used as fat replacer, gell ing agent, thickening agent [3-6]. It is also considered a member of a class of drugs known as biological response modifiers (BRM) [7]. It has been reported that (1→3)-β-glucan enhances the innate host defense response by binding to a specific receptor on the plasma membrane of macrophages, resulting in their activation, and this activity is associated with anti-tumor, anti-bacterial and wound-healing activities [8-12]. However, the native glucan was an insoluble polysaccharide, whi ch became a major b arrier to the utilization of (1→3)-β-glucan as BRMs. If (1→3 )-β-glucan is to become clinically applicable, it has to be converted into biologically effective, water-soluble form that can be safely administered via the systemic route. Numerous studies have therefore fo- cused on converting these glucans into a water-soluble form through chemical modification such as amination [13], sulfation [14] and phosphorylation [15]. It has also been demonstrated that such modified polysaccharides exhibit anti-inflammatory, anti-tumor, anti-viral and immunomodulatory activities [16-18]. In this study, we have attempted to convert insoluble yeast glucans into a soluble form through phosphorylation process. However, in this process we found that only DMSO or DMSO plus urea (without H3PO4) could conver t i nso lub l e glu can i nt o a soluble form. Therefore, four kind of preparation of soluble glucan process were investigated. And four different soluble products were compared by solubility, FT-IR spectra, 13C NMR spectra, and helix-coil transition analysis and immunoprophy- laxis ef ficacy assay. 2. Materials and Methods 2.1. Materials Isolated cell walls fro m Baker’s yeast were sup plied as “spray- dried” powder by Angel Yeast LLC, China. Male ICR/HSD mice (18-20 g) were supplied by the Experimental Animal Center of the Academy of Military Medical Science (AMMS), Beijing, China. Mice were housed in a specific pathogen-free (SPF) environment. All other major reagents were purchased from Sigma (St. Louis, MO, USA). 2.2. Preparation of Particulate Glucans Particulate glucans were isolated from the cell wall of S. cere- visiae by a modification of the method of Williams et al [15]. Briefly, the cell wall of S. cerevisiae was dispersed in 0.3 L of 0.75 M (3 %) sodium hydroxide (NaOH), and extracted in wa- ter bath at 75 ºC for 2 hours. The suspension was cooled to room temperature and centrifuged (4000 rpm, 10 min), the resulting pellet collected and this material then washed three times using 0.5 L distilled water. The residue was then mixed with 0 .2 L of 0.75 M NaOH, and ext racted in bo iling water for 3 hours. Distilled water was then added to the cooled suspen- sion, the pH adjusted to 7 with HCl, and the supernatant after centrifugation discarded. This water wash was repeated until the residue became white and flocculent, and finally the pellet was washed with abso lute ethanol u ntil the sup ernatant became colorless. Particulate glucan was obtained by drying the washed pellets. 2.3. Preparation of Soluble Glucans Different soluble glucans were prepared by using the improved  L. P. DU ET AL. Copyright © 2012 SciRes. E NG 185 method of Williams et al [1 5]. Briefly, particulate glucan (1 g) was suspended in 60 ml dimethyl sulfoxide (DMSO) containing 15 g u rea. H3PO4 (85%, 15 ml) was added dropwise to the solu- tion at room temperature (total volume 75 ml, volume ratio between H3PO4 and DMSO 1:4). The solution was then heated to 100 ºC for 5 hours, while the solution was constantly stirred, to bring about the reaction. A crystalline precipitate (presuma- bly urea phosphate) formed after about 1.5 hours. The reaction mixture was then cooled to ambient temperature, and 500 ml distilled water added to dilute the solution. The solution was then dialyzed against deionizer water for 5 days to remove Me2SO, H3PO4, urea and any other small compounds, and then filtrated , concentrated, and freeze-dried to obtain the pure sam- ples. Different soluble glucans were generated by varying the dosage of urea and H3PO4, which were tabulated in Table 1. 2.4. Determination of Water Solubility The water solubility of glucans was measured using a modifica- tion of the method of Chang et al [19]. Briefly, each sample (1 g) was suspended in distilled water (5 ml) and the resulting suspension agitated at 25 ºC for 24 hours. The sample was cen- trifuged at 1600 × g for 15 min. The supernatant (2 ml) was mixed with three volumes of ethanol to precipitate the glucan. The precipitates were recovered by centrifugation at 1600 × g for 15 min, vacuum-dried at 60 ºC, and weighed. The solubility was calculated using the formula below: Solubility(%)100(2.5Dry precipitate weight)(Dry sample weight)=×× 2.5. Infrared Spectroscopy IR spectra were recorded using the KBr-disk method with a Bruker VECTOR 22 Fourier transform infrared (FTIR) spec- trometer (Bruker, Germany) in the range 400 to 4000 cm-1. Sixteen scans at a reso lution of 4 cm-1 were averaged an d refe- renced agai nst air. 2.6. Nuclear Magnetic Resonance Spectroscopy (NMR) 13C NMR spectra were recorded on Bruker AVANCE 600 MHz spectrometers (Bruker BioSpin, Swiss) at room temperature. All samples were dissolved in Me2SO-d6 (heat at 80 ºC for 30 min) to a final concentration of 80 mg/ml. Tetramethylsilane (TMS) was used as internal standard. 2.7. Analysis of Helix-co il Transition The conformational structure of the glucans in solution was determined by characterizing Congo red-polysaccharide com- plexes. The transition from a triple-helical arrangement to the single-stranded conformation was examined by measuring the Table 1. Dosage of ure a and H3PO4. Group Gl u c an name Urea (g) H3PO4 (ml) A Glucan A 0 0 B Glucan B 18 0 C Glucan C 0 10 D Glucan D 18 10 λmax of Congo red-polysaccharide solutions at NaOH concen- trations ranging from 0.0025 to 0.5 M. Polysaccharide aqueous solutions (0.5 mg/ml) containing 2 ml of 312 μM Congo red were treated with different concentrations of NaOH (2 ml). Visible absorption spectra were recorded with a UV/visible spectrophoto meter (type 2401PC) at each alkali concentrati on. 2.8. Immunoprophylaxis Efficacy Assay Fifty ICR/HSD mice were divided into five groups. Groups A to D were injected intraperitoneally with Glucan A, Glucan B, Glucan C and Glucan D respectively at a dose of 200 mg/kg (4 mg per mouse) 24 hours prior to intraperitoneal challenge with 1×109 viable E. coli. While group O was injected with isovo- lumetric dextrose 24 hours prior to challenge with E. coli served as contr ol. The same clini cal isolate of E. coli was used in all cases, and was maintained in our laboratory. E. coli was cultured in trypticase soy broth for 18 hours at 37 ºC in an air bath shaker at 180 rpm. The culture was then centrifuged (3500 rp m, 8 min ), the cel l pell et washe d three ti mes, and this mixtu re diluted to provide 1 × 109 E. coli/ml. Survival of each group of mice was monitored hourly during the 24 hour period. 3. Results and Discussion 3.1. The Yield and Solubility of Glucan Samples The yield and solubility of glucans were shown in Figure 1. The yield of glucan A and C was very low (~18%). However, the yield of glucan B and D was higher. This could be con- clud ed that DMSO and urea had effects o n the yield of glucan s. DMSO and urea was used as denaturant could facilitate the breakdown of the intermolecular hydrogen bonds between the glucan chains. As a result, more hydroxyl group exposed, which was beneficial to the solubility of glucan. Meanwhile the influence of urea on the yield was more powerful than DMSO. The solubility of glucan C and D was significantly higher than glucan A and B, which could be attribute to the introduction of phosphate group. 3.2. FT-IR Spectra of Di fferent Glucan Samples The FT-IR spectra of the glucans are shown in Figure 2. The Glucan sample AB C D Yield(%) 0 20 40 60 80 100 Solubility(%) 0 20 40 60 80 100 Yield Solubility Figure 1. The yield an d solu bil it y o f glucan sam p l es . 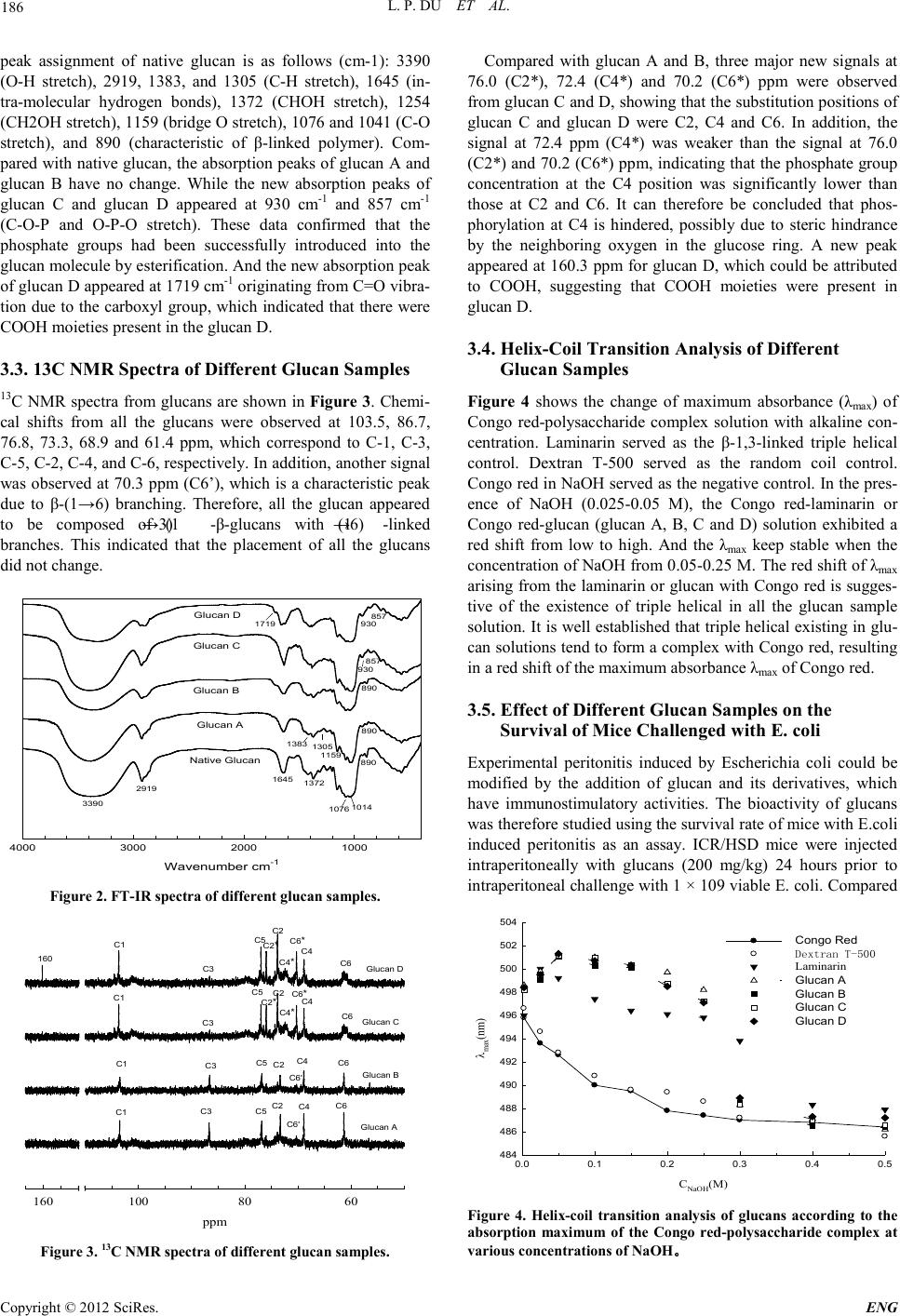 L. P. DU ET AL. Copyright © 2012 SciRes. ENG 186 peak assignment of native glucan is as follows (cm-1): 3390 (O-H stretch), 2919, 1383, and 1305 (C-H stretch), 1645 (in- tra-molecular hydrogen bonds), 1372 (CHOH stretch), 1254 (CH2OH stretch), 1159 (bridge O stretch), 1076 and 1041 (C-O stretch), and 890 (characteristic of β-linked polymer). Com- pared with native glucan, the absorption peaks of glucan A and glucan B have no change. While the new absorption peaks of glucan C and glucan D appeared at 930 cm-1 and 857 cm-1 (C-O-P and O-P-O stretch). These data confirmed that the phosphate groups had been successfully introduced into the glucan molecule by esterification. And the new absorption peak of glucan D appeared at 17 19 cm-1 originating from C=O vibra- tion due to the carboxyl group, which indicated that there were COOH moieties pres ent in the glucan D. 3.3. 13C NMR Spectra of Different Glucan Samples 13C NMR spectra from glucans are shown in Figure 3. Ch emi- cal shifts from all the glucans were observed at 103.5, 86.7, 76.8, 73.3, 68.9 and 61.4 ppm, which correspond to C-1, C-3, C-5, C-2, C-4, and C-6, respectively. In addition, another signal was observed at 70.3 ppm (C6’), which is a characteristic peak due to β-(1→6) branching. Therefore, all the glucan appeared to be composed of (1→3) -β-glucans with (1→6) -linked branches. This indicated that the placement of all the glucans did not cha ng e. Wavenumber cm -1 1000200030004000 Native Glucan Glucan A Glucan B Glucan C Glucan D 3390 2919 1645 1372 890 1159 1076 1014 1383 1305 930 930 857 857 1719 890 890 Figure 2. FT-IR spectra of different glucan samples. ppm 6080100160 Glucan A Glucan B Glucan C Glucan D C1 C3 C5 C2 C6 C4 C1 C3 C5 C2 C4 C6 C1 C1 C3 C3 C5 C5 C2 C2 C4 C4 C6 C6 C6 * C6 * C4 * C4 * C2 * C2 * 160 C6' C6' Figure 3. 13C NMR spectra of different glucan samples. Compared with glucan A and B, three major new signals at 76.0 (C2*), 72.4 (C4*) and 70.2 (C6*) ppm were observed from glucan C and D, showing that the substitution positions of glucan C and glucan D were C2, C4 and C6. In addition, the signal at 72.4 ppm (C4*) was weaker than the signal at 76.0 (C2*) and 70.2 (C6*) ppm, indicating that the phosphate group concentration at the C4 position was significantly lower than those at C2 and C6. It can therefore be concluded that phos- phorylation at C4 is hindered, possibly due to steric hindrance by the neighboring oxygen in the glucose ring. A new peak appeared at 160.3 ppm for glucan D, which could be attributed to COOH, suggesting that COOH moieties were present in glucan D. 3.4. Helix-Coil Transition Analysis of Different Glucan Sa mple s Figure 4 shows the change of maximum absorbance (λmax ) of Congo red-polysaccharide complex solution with alkaline con- centration. Laminarin served as the β-1,3-linked triple helical control. Dextran T-500 served as the random coil control. Congo red in NaOH served as the negative co ntrol. In the pres- ence of NaOH (0.025-0.05 M), the Congo red-laminarin or Congo red-glucan (glucan A, B, C and D) solution exhibited a red shift from low to high. And the λmax keep stable when the concentration of NaOH from 0.05-0.25 M. The red shift of λmax arising from the laminarin or glucan with Congo red is sugges- tive of the existence of triple helical in all the glucan sample solution. It is well established that triple helical existing in glu- can solutions tend to form a complex with Congo red, resulting in a red shift of the maximum absorbance λmax of Congo red. 3.5. Effect of Different Glucan Samples on the Survival o f Mice Cha lle nged with E. c oli Experimental peritonitis induced by Escherichia coli could be modified by the addition of glucan and its derivatives, which have immunostimulatory activities. The bioactivity of glucans was therefor e stu di ed usin g the survival rat e of mice with E. col i induced peritonitis as an assay. ICR/HSD mice were injected intraperitoneally with glucans (200 mg/kg) 24 hours prior to intraperitoneal challenge with 1 × 109 viable E. coli. Compared CNaOH(M) 0.0 0.1 0.2 0.3 0.4 0.5 λmax(nm) 484 486 488 490 492 494 496 498 500 502 504 Congo Red Dextran T-500 Laminarin Glucan A Glucan B Glucan C Glucan D Figure 4. Helix-coil transition analysis of glucans according to the absorption maximum of the Congo red-polysaccharide complex at various con cen t r at ions of NaOH 。  L. P. DU ET AL. Copyright © 2012 SciRes. E NG 187 Time(h) 05 10 15 20 Survival(%) 0 20 40 60 80 100 Dextrose Glucan A Glucan B Glucan C Glucan D Figure 5. Effects of different glucan samples on survival rate of mice cha llenged wit h E. co li . to the control group (dextrose), all glucans had a potent immu- noprophylaxis efficacy that significantly (p < 0.05) enhanced the survival rate and the survival time of ICR/HSD mice chal- lenged with E. coli (Figure 5). The survival rate of mice was 60 %, 40 %, 70 % and 80 % respectively, for glucan A, glucan B, glucan C and glucan D. In contrast, the control group mice showed 0 % survival after 9 hours. This indicated that introduc- tion of phosphate group not only enhanced the solubility, but also improved the immunoprophylaxis efficacy. 4. Conclusion Four kinds of soluble glucans were successfully prepared by four processes. DMSO and urea was used as denaturant could facilitate the breakdown of the intermolecular hydrogen bonds between the glucan chains. As a result, more hydroxyl group exposed, which was beneficial to the solubility and substitution reaction of glucan. Meanwhile, urea has a significant effect on yield, and DMSO has a significant effect on solubility. From FT-IR and 13C NMR spectra we can see that there was no dif- ference between native and glucan A, B. Glucan C and D was successfully phosphorylated. And the substitution positions were C2, C4 and C6. However the hydroxyl group of glucan D at C6 was partially oxidized into a carboxyl group, which was caused by urea and H3PO4 together. All the glucans had the triple helical structure and the immunoprophylaxis efficacy. However the immunoprophylaxis efficacy of phosphorylated glucan was better. The introduction of phosphate group not only enhanced the solubility of glucan, but also improved the survival rate of mice challenged with E. coli. 5. Acknowledgements This study was financially supported by the 863 (Hi-tech re- search and development program of China) program under contract NO. 2012AA021505. We also would like to express our deepest appreciation to Tianjin University of Science & Technology for the financial support (Grant number:20100208) REFERENCES [1] W. Z. Hassid, M. A. Joslyn, and R. M. McCready, “The molecu- lar constitution of an insoluble polysaccharide from yeast, Sac- charomyces cerevisiae,” Journal of the American Chemical So- ciety, vol. 63, pp. 295-298, 1941. [2] N. R. Di Luzio, D. L. Williams, R. B. McNamee et al., “Com- parative tumor-inhibitory and anti-bacterial activity of soluble and particulate glucan,” International Journal of Can ce r, vol. 24, pp. 773-779, 1979. [3] N. Sahan, K. Yasar, and A. Hayaloglu, “Physical, chemical and flavour quality of non-fat yogurt as affected by a [beta]-glucan hydrocolloidal composite during storage,” Food Hydrocolloids, vol. 22, pp. 1291-1297, 2008. [4] R. Santipanichwong, and M. Suphantharika, “Carotenoids as colorants in reduced-fat mayonnaise containing spent brewer's yeast [beta]-glucan as a fat replacer,” Food Hydrocolloids, vol. 21, pp. 565-574, 2007. [5] S. Worrasinchai, M. Suphantharika, S. Pinjai et al., “[be- ta]-Glucan prepa red from sp ent b rewer's yeast a s a fat replacer in mayonnaise,” Food Hydrocolloids, vo l . 20, pp. 68-78, 2006. [6] A. Xi u, M. Zhou, B. Zhu et al., “Rh eologica l prop erties o f Sale- can as a new source of thickening agent,” Food Hydrocolloids, vol. 25, pp. 1719-1725, 2011. [7] J. A. Bohn, and J. N. BeMiller, “(1-->3)-[beta]--Glucans as bio- logical response modifiers: a review of structure-functional ac- tivity relationships,” Carbohydrate Polymers, vol. 28, pp. 3-14, 1995. [8] L. Elyakova, V. Isakov, L. Lapshina et al. , “En zymati c tran sfor- mation of biologically active 1,3;1,6-β-D-glucan. Structure and acti vity of result ing fragm ents,” Biochemistry (Moscow), vol. 72, pp. 29-36, 2007. [9] A. Mueller, J. Raptis, P. J. Rice et al., “The influence of glucan polymer structure and solution conformation on binding to (1-->3)-beta-D-glucan receptors in a human monocyte-like cell line,” Glycobiology, vol. 10, pp. 339-346 , 2000. [10] N. Ohno, T. Miura, N. N. Miura et al., “Structure and biological activities of hypochlorite oxidized zymosan,” Carbohydrate Po- lymers, vol. 44, pp. 339-349, 200 1. [11] E. Tsiapali, S. Whaley, J. Kalbfleisch et al., “Glucans exhibit weak antioxidant activity, but stimulate macrophage free radical activity,” Free Radical Biology and Medicine, vol. 30, pp. 393-402, 2001. [12] D. L. Williams, H. A. Pretus, R. B. McNamee et al., “Develop- ment, physicochemical characterization and preclinical efficacy evaluation of a water soluble glucan sulfate derived from Sac- charomyces cerevisiae,” Immunopharmacology, vol. 22, pp. 139-156, 1991. [13] M. S. Shin, S. Lee, K. Y. Lee et al., “Structural and biological characterization of aminated-derivatized oat beta-glucan,” Jour- nal o f Agri cultur al and Food C hemist ry, vol. 53, pp. 5 554-5558, 2005. [14] D. L. Williams, H. A. Pretus, R. B. McNamee et al., “Develop- ment of a water-soluble, sulfated (1 --> 3)-[beta]-d-glucan bio- logical response modifier derived from Saccharomyces cerevi- siae,” Carbohydrate Research, vol. 235, pp. 247-257, 1992. [15] D. L. Williams, R. B. McNamee, E. L. Jones et al., “A method for the solubilization of a (1-->3)-[beta]--glucan isolated from Saccharomyces cerevisiae,” Carbohydrate Research, vol. 219, pp. 203-213, 1991. [16] X.-M. Chen, J. Zhang, and G.-Y. Han, “Studies on synthesis and antitumor activity of phosphorylated achyranthes bidentata poly- saccharide (P-AbPS),” Chinese Journal of Chemistry, vol. 20, pp. 1406-1410, 2002. [17] O. I. Lyuksutova, E. D. Murphey, T. E. Toliver-Kinsky et al., “Glucan phosphate treatment attenuates burn-induced in flamma- tion and improves resistance to Pseudomonas aeruginosa burn  L. P. DU ET AL. Copyright © 2012 SciRes. ENG 188 wound infection,” Shock, vol . 23, pp. 224-232, Mar, 2005. [18] D. L. Williams, T. Ozment-Skelton, and C. Li, “Modulation of the phosphoinositide 3-Kinase signaling pathway alters host re- sponse to sepsis, inflammation, and ischemia/reperfusion injury,” Shock, vol. 25, pp. 432-439, 2006. [19] P. S. Chang, and G. B. Cho, “Oxidation of primary alcohol groups of polysaccharides with 2,2,6,6-tetramethyl-1-piperidine oxoammonium ion.,” Korean Journal of Food Science and Technology, vol. 29, pp. 446-451, 1997. |