A. G. YAKOVLEV, V. A. SHUVAL OV
Copyright © 2013 SciRes. OJAppS
P* state. It is clear that further studies are need in this
question.
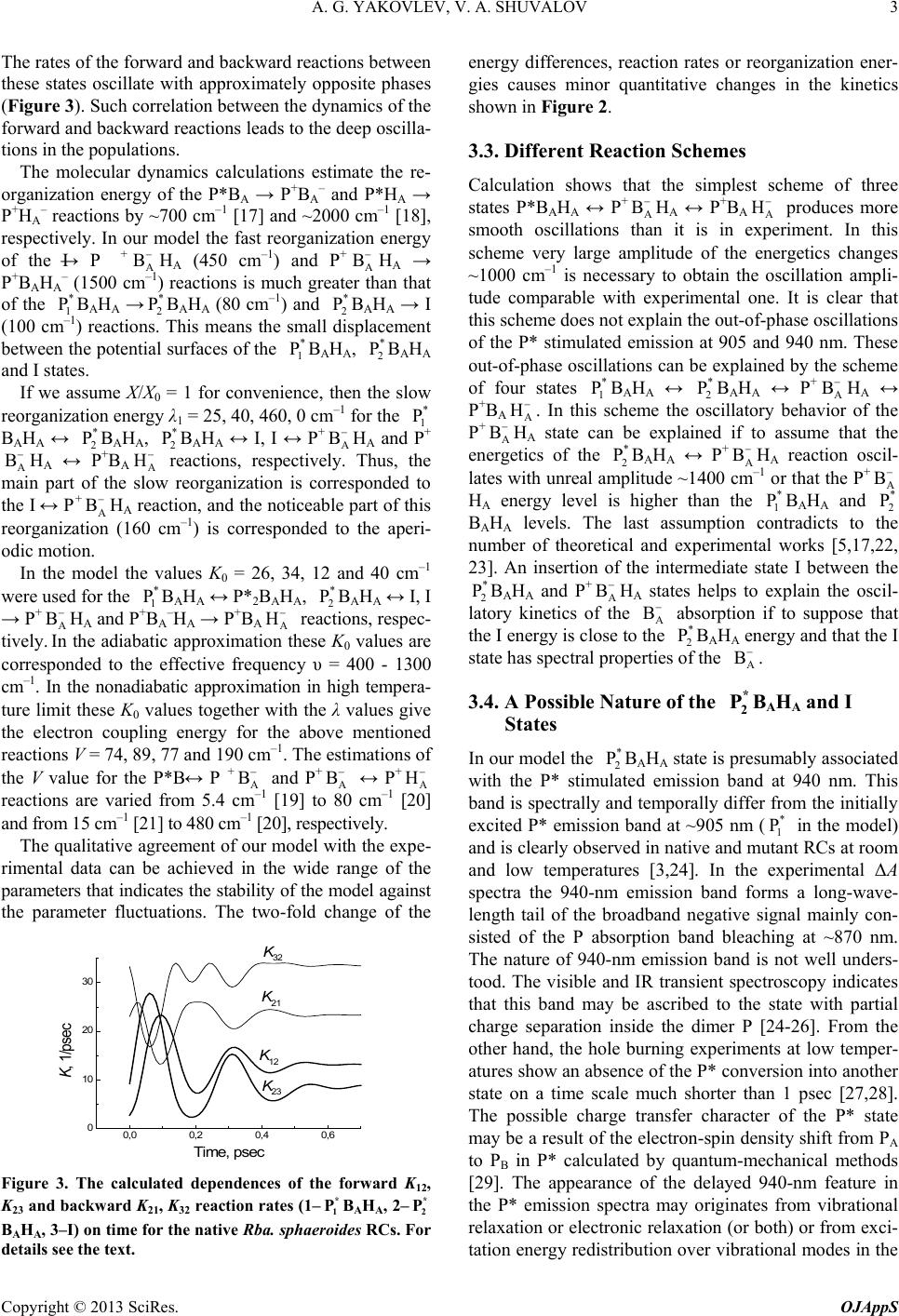
As it was mentioned above, the I state was introduced
into the model for better fitting of the experimental
absorption kinetics at 1020 nm. The population dynamics
of the I state is close to the oscillatory component of the
experimental
absorption band dynamics. The cal-
culated sum of the I and P+
HA populations is close to
the experimental A
B− kinetics. These results indicate
that the I state may contain
or ВА
δ–. One may spe-
culate that I is the state with charge transfer character
between P* and BA. To distinguish the I and P+A
B−HA
states in experiment is very difficult because of their
spectral and temporal closeness. No clear experimental
evidence of the I state existence is available now.
In our opinion, an advantage of the schemes with the
BAHA and I intermediates consists in the high rates of
the reactions in which these states are involved. The
BAHA and I states act as fast mediators between the in-
itially excited (
BAHA) and charge separated states. A
stabilization of electron occurs lately in the P+
state
and partially in the P+
state.
3.5. A Nature of the Oscillations
In our model the oscillations in the populations are
caused by external modulation of the reaction energetics.
A nature of this modulation is beyond the scope of the
model. The nuclear motion (not only inside P) can be a
possible source of this modulation. Shortly after the ex-
citation of P by broadband femtosecond pulse the nuclear
motion has a coherent character. When the coherence is
rapidly damped this motion may continue incoherently
due to the nuclear inertia. One can speculate that the in-
coherent motion may exist a longer timescale than the
coherent one, but the latter produces the greater ampli-
tude of the oscillations. The observed oscillations may
reflect both the coherent and incoherent motion. The
presented model shows th at at t > 150 fsec the oscillatory
features observed in the kinetics of the excited and
charge separated states can be explained by incoherent
modula t ion of the reaction energetics.
4. Acknowledgements
We are grateful to L.G. Vasilieva and A.Ya. Shkuropatov
for preparing RC samples. We acknowledge the partial
financial support of the Russian Foundation for Basic
Research.
REFERENCES
[1] W. W. Parson and A. Warshel, “Mechanism of Charge
Separation in Purple Bacterial Reaction Centers,” In: C. N.
Hunter, F. Daldal, M. C. Thurnauer and J. T. Beatty, Eds.,
The Purple Phototrophic Bacteria, Springer Science +
Business Media B.V., Dordreht, 2009, pp. 355-377.
http://dx.doi.org/10.1007/978-1-4020-8815-5_19
[2] V. A. Shuvalov, “Primary Light Energy Conversion in
Reaction Centers of Photosynthesis,” Nauka, Moscow,
2000.
[3] M. H. Vos, M. R. Jones, C. N. Hunter, J. Breton, J.-C.
Lambry and J.-L. Martin, “Coherent Dynamics during the
Primary Electron-Transfer Reaction in Membrane-Bound
Reaction Centers of Rhodobacter sphaeroides,” Bioche-
mistry, Vol. 33, 1994, pp. 6750-6757.
http://dx.doi.org/10.1021/bi00188a002
[4] A. G. Yakovlev, A. Ya. Shkuropatov and V. A. Shuvalov,
“Nuclear Wavepacket Motion Producing a Reversible
Charge Separation in Bacterial Reaction Centers,” FEBS
Letters, Vol. 466, 2000, pp. 209-212.
http://dx.doi.org/10.1016/S0014-5793(00)01081-4
[5] V. I. Novoderezhkin, A. G. Yakovlev, R. van Grondelle
and V. A. Shuvalov, “Coherent Nuclear and Electronic
Dynamics in Primary Charge Separation in Photosynthet-
ic Reaction Centers: A Redfield Theory Approach,” The
Journal of Physical Chemistry B, Vol. 108, 2004, pp.
7445-7457. http://dx.doi.org/10.1021/jp0373346
[6] D. S. Shchepetov, D. S. Cher navsky , V. V. Gorokhov, N.
P. Grishanova, V. Z. Paschenko and A. B. Rubin, “The
Nature of Oscillations in the Kinetics of Electron Transfer
in the Reaction Center of Purple Bacteria,” Doklady Bio-
chemistry and Biophysics, Vol. 425, 2009, pp. 87-90.
http://dx.doi.org/10.1134/S1607672909020082
[7] J. M. Jean and G. R. Fleming, “Competition between
Energy and Phase Relaxation in Electronic Curve Cross-
ing Processes,” Journal of Chemical Physics, Vol. 103,
1995, pp. 2092-2101. http://dx.doi.org/10.1063/1.469684
[8] K. Ando and H. Sumi, “Nonequilibrium Oscillatory Elec-
tron Transfer in Bacterial Photosynthesis,” The Journal of
Physical Chemistry B, Vol. 102, 1998, pp. 10991-11000.
http://dx.doi.org/10.1021/jp982659l
[9] X. Gu, M. Hayashi, S. Suzuki and S. H. Lin, “Vibrational
Coherence and Relaxation Dynamics in the Primary Do-
nor State of the Mutant Reaction Center of Rhodobacter
capsulatus: Theoretical Analysis of Pump -Probe Stimu-
lated Emission,” Biochimica et Biophysica Acta, Vol.
1229, 1995, pp. 215-224.
http://dx.doi.org/10.1016/0005-2728(94)00197-D
[10] S. H. Lin, M. Hayashi, S. Suzuki, X. Gu, W. Xiao and M.
Sugawara, “Theoretical Analyses on Femtosecond Time-
Resolved Spectra of Initial Electron Transfer of Photo-
synthetic Reaction Centers at Low Temperatures,” Che-
mical Physics, Vol. 197, 1995, pp. 435-455.
http://dx.doi.org/10.1016/0301-0104(95)00186-R
[11] J. A. Leegwater, “Theory of Coherent Oscillations in the
Photosynthetic Reaction Center,” The Journal of Physical
Chemistry, Vol. 99, 1995, pp. 11605-11611.
http://dx.doi.org/10.1021/j100029a044
[12] A. Lucke, C. H. Mak, R. Egger, J. Ankerhold, J. Stock-
burger and H. Grabert, “Is the Direct Observation of Elec-
tronic Coherence in Electron Transfer Reactions Possi-
ble?” Journal of Chemical Physics, Vol. 107, 1997, pp.
8397-8408.