 Advances in Bioscience and Biotechnology, 2013, 4, 63-74 ABB http://dx.doi.org/10.4236/abb.2013.410A3008 Published Online October 2013 (http://www.scirp.org/journal/abb/) Regulation of D-galacturonate metabolism in Caulobacter crescentus by HumR, a LacI-family transcriptional repressor Aaesha I. Sheikh, Deborah Caswell, Cynthia Dick, Spencer Gang, Justin Jarrell, Ankita Kohli, Amanda Lieu, Jared Lumpe, Meghan Garrett, Jennifer Parker, Craig Stephens Biology Department, Santa Clara University, 500 El Camino Real, Santa Clara, USA Email: Cstephens@scu.edu Received 28 July 2013; revised 28 August 2013; accepted 19 September 2013 Copyright © 2013 Aaesha I. Sheikh et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT The oligotrophic freshwater bacterium Caulobacter crescentus encodes a cluster of genes (CC_1487 to CC_1495) shown here to be necessary for metabolism of D-galacturonate, the primary constituent of pectin, a major plant polymer. Sequence analysis suggests that these genes encode a version of the bacterial he- xuronate isomerase pathway. A conserved 14 bp se- quence motif is associated with promoter regions of three operons within this cluster, and is conserved in homologous gene clusters in related alpha-Proteobac- teria. Embedded in the hexuronate gene cluster is a gene (CC_1489) encoding a member of the LacI fam- ily of bacterial transcription factors. This gene prod- uct, designated here as HumR (hexuronate metabo- lism regulator), represses expression of the uxaA and uxaC operon promoters by binding to the conserved operator sequence. Repression is relieved in the pre- sence of galacturonate or, to a lesser extent, by glu- curonate. Other genes potentially involved in pectin degradation and hexuronate transport are also under the con tro l of Hu mR. Ad option of a LacI-type repres- sor to co ntro l hex uro nate me tab olis m para llels the r e- gulation of xylose, glucose, and maltose utilization in C. crescentus, but is distinct from the use of GntR- type repressors to control pectin and hexuronate uti- lization in gamma-Proteobacteria such as Escherichia coli. Keywords: Caulobacter; Galacturonate; LacI 1. INTRODUCTION Pectin is an important plant polysaccharide that aids the cross-linking of cellulose and hemicellulose to maintain the integrity of plant cells and tissues [1]. The core pectin polymer consists of alpha-1, 4-linked D-galacturonate subunits, which are then modified by methylation and va- rious oligosaccharide side chains depending on the plant source [1]. Metabolism of galacturonate, which makes up 40% - 70% of the dry weight of pectin, has been studied extensively in just a few microbes [2,3]. In this work, we show that the oligotrophic, stalked alpha-Proteobacte- rium Caulobacter crescentus degrades galacturonate through the hexuronateisomerase pathway, and regulates galactu- ronate metabolism via LacI-family repressor. This sys- tem for galacturonate metabolism and regulation is con- served in several closely related alpha-Proteobacteria, al- though the metabolic pathway had previously been ob- served only in gamma-Proteobacteria such as Escheri- chia coli and Erwinia chrysanthemi. Microorganisms degrade pectin through a suite of se- creted enzymes, with pectatelyase and exo-polygalactu- ronidase primarily responsible for chain depolymeriza- tion [2,4]. The most extensively studied pectinolytic bac- terium is the phytopathogen Erwinia chrysanthemi, which attacks pectin to facilitate invasion of plant tissue during infection [5]. Erwinia extracts nutrients from pectin deg- radation products by consuming oligo-galacturonides, ga- lacturonate, and 5-keto-4-deoxyuronate through the hex- uronatei somerase pathway it shares with non-phytopatho- genic Enterobacteriaceae such as E. coli [3]. Caulobacters are dimorphic alpha-Proteobacteria that often exhibit an adhesive stalk and readily form biofilms [6]. It is likely that species such as C. crescentus colonize decaying vegetation in aquatic habitats, secreting hydro- lytic enzymes to access nutrients embedded in the inso- luble cell walls [7]. Whole genome sequencing of C. cre- scentus revealed numerous genes encoding secretable en- zymes for attacking cellulose, xylans, pectin, and other polymers [8]. Genomic analysis of closely related stalked OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 64 bacteria such as Caulobacter henriciistrain K31 [9] and Phenylobacterium zucineum [10] shows comparable ros- ters of secretable hydrolases. In C. crescentus, there is evidence for coordinated expression of gene sets that in- clude extracellular enzymes, transport systems, and cyto- plasmic catabolic pathways when cells are exposed to re- levant sugars, such as glucose and xylose [11]. For ex- ample, transcriptome analysis of C. crescentus metaboli- zing xylose revealed induction of xylanases and arabi- nosidases, putative outer and inner-membrane transport- ers, and a pathway for oxidative metabolism of xylose [11,12]. The xylose regulon is under the control of XylR, a member of the LacI family of bacterial transcription factors [13]. Our results reveal an analogous galacturo- nate regulon in C. crescentus that is under the control of another LacI family member, HumR. Given that the xy- lose operon promoter [14], in conjunction with XylR [13], has proven to be a robust tool for tightly controlled indu- cible gene expression in C. crescentus, the identification of HumR not only reveals commonalities in regulatory strategies, but may also expand the molecular genetic ar- senal available for research applications in this organism. 2. MATERIALS AND METHODS 2.1. Bacterial Strains and Growth Conditions Escherichia coli strains1 were grown in Luria-Bertani (LB) broth or agar, supplemented as necessary with kanamy- cin (50 μg/ml), or oxytetracycline (10 μg/ml). Caulobac- ter crescentus strains were routinely grown in PYE broth or agar [15], supplemented as necessary with kanamycin (20 μg/ml in agar, 5 μg/ml in broth), oxytetracycline (2 μg/ml in agar, 1 μg/ml in broth), and/or nalidixic acid (20 μg/ml in agar). When indicated, cultures were grown on M2 minimal salts medium [15] containing the carbon/ energy source at a final concentration of 10 mM. The D- isomer was used for all monosaccharides (galacturonate, glucuronate, glucose, galactose, and xylose). Two sourc- es of pectin were tested: pectin derived from citrus fruits (P9135, Sigma-Aldrich Chemical Co.), and pectin deriv- ed from apple (P8471, Sigma-Aldrich Chemical Co.). A final concentration of 1.94 g/l pectin was used in media. Growth curves were conducted in 125 ml baffled flasks containing 50 ml PYE or M2 media. Cultures were in- cubated at 30˚C in a bench top shaker at 250 - 300 rpm, and samples were taken periodically to monitor turbidity. Phenotype Microarrays (Biolog, Hayward, CA) [16] were also used to examine substrate utilization, as described previously [12]. The PM1 and PM2 plates contain 190 distinct carbon sources, and increases in culture turbidity were used to monitor productive substrate utilization. 2.2. Construction of Mutations Knockouts were made by either integrative disruption or non-polar in-frame deletion [17]. Polymerase chain reac- tion (PCR) primers were synthesized by Integrated DNA Technologies (IDT; San Diego, CA). PCR was done us- ing GoTaq thermostable polymerase (Promega Corp., Madison, WI). Chromosomal DNA was prepared using the DNEasy kit (Qiagen), according to the manufactu- rer’s instructions. Integrative disruptions were construct- ed as described previously [17], resulting in insertion of the kanamycin-resistant pBGS18T plasmid into the target gene. Construction of non-polar in-frame deletions was done by a PCR-mediated process [17], which results in a deletion of coding sequence and no insertion. For inte- grative disruptions, the presence of integrated plasmid DNA was verified by PCR. Stability of the integrated plas- mids was verified by growth under non-selective condi- tions (PYE), followed by plating in the absence or pres- ence of kanamycin. After growth under non-selective con- ditions (PYE) for at least 10 generations, less than 1% of the population had lost the integrated plasmid-encoded kanamycin resistance. Deletion constructs were also ve- rified by PCR. 2.3. Electrophoretic Mobility Shift Assays C. crescentus strains were grown in 500 ml PYE cultures at 30˚C to mid-log phase (OD600 ~ 0.4), and cell lysates were prepared as described previously [13]. Target DNA for binding assays was designed based on the 14 bp HumR operator sequence in the uxaC promoter region. The sequence was slightly modified outside of the puta- tive operator motif to reduce predicted secondary struc- ture, based on mFold [18]. Two 34-base oligonucleotides were synthesized: 5’-CTACGCGCGATGACACCGG- TTACCAGCAGACACG and 5’-CGTGTCTGCTGGT- AACCGGTGTCATCGCGCGTAG (putative HumR ope- rator underlined). The 5’ end of each was labeled with Cy5 (IDT). Oligonucleotides were annealed in equimolar amounts by first denaturating at 95˚C for 5 min, then cooling slowly for 60 minutes to room temperature. Bin- ding reactions consisted of 2.5 ng annealed duplex target DNA, binding buffer (150 mM KCl, 0.1 mM dithiothrei- tol, 0.1 mM EDTA and 10 mM Tris (pH 7.4)), 1 μg poly- dI: dC, and 22.5 μg protein in a total reaction volume of 15 μl. For binding inhibition assays, galacturonate, glu- curonate, glucose, xylose, or galactose were added to 10 mM final concentration as appropriate. Reactions were incubated at room temperature for 10 minutes, and then resolved on a 6% non-denaturing polyacrylamide gel made and run with 1× Tris-borate-EDTA buffer. Gels were run at 100 V for 50 min, visualized on a Typhoon imag- ing system (Molecular Dynamics) and quantified using Image Quant software. Background counts were deter- 1Detailed descriptions of bacterial strains, plasmid constructs, and pri- mers are available from the authors upon request. Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 65 mined in lanes where no protein was added, and sub- tracted from the counts for lanes that contained cell ex- tracts. 2.4. Expression Assays and Operator Mutants To examine regulation of gene expression in response to galacturonate, the regions upstream of the CC_1488 (uxaA) and CC_1490 (uxaC) genes, respectively, were amplified by PCR. For wild-type promoters, PCR prim- ers were designed to amplify up to 250 bp upstream of the start codons of these genes, along with a few hundred base pairs within the coding region. Synthetic EcoRI or HindIII sites were added to the 5’ end of the primers to allow for directional cloning. PCR products were initial- ly cloned into plasmid PCR2.1 using the TOPO Cloning System (Life Technologies), then subcloned into pRKlac290, a low copy-number, broad host range plas- mid to generate transcriptional fusions to the lacZ gene [19]. pRKlac290 constructs were transferred from E. coli S17.1 by conjugation into C. crescentus strains, with se- lection on PYE supplemented with nalidixic acid and oxytetracycline. Construction of targeted mutations in the putative operator for the uxaC promoter was done with a two-step PCR-based method [17]. Promoter con- structs containing the mutant operators were cloned as described above, and β-galactosidase activity was assay- ed as described previously [13,19]. 2.5. qRT-PCR Analysis of Gene Expression Quantitative real-time PCR (qRT-PCR) was used to ex- amine gene expression, starting with RNA isolated from C. crescentus strains grown in PYE medium, with or without galacturonate. RNA was extracted as described previously [11]. Isolated RNA was subjected to two con- secutive genomic DNA removal treatments by digesting with 2 U DNaseI (Ambion) per 10 μg RNA, incubated at 37˚C for 60 min. DNaseI was inactivated by addition of phenol:chloroform (5:1) and RNA was precipitated with 3M sodium acetate (pH 5.2) and 100% ethanol at −20˚C overnight. Pellets were obtained by centrifugation at 16,000 × g for 15 min, washed with 70% ethanol and re- suspended in RNase free water. RNA purity was deter- mined spectrophotometrically and agarose gels were run to verify RNA integrity. Complementary DNA (cDNA) was synthesized with 2.0 - 2.5 μg of RNA and SuperScriptII Reverse Tran- scriptase (200 units, Life Technologies) with random pri- mers (100 ng per 5 μg total RNA, Life Technologies) fol- lowing the manufacturer’s suggested protocol, including the optional RNaseH digestion (New England Biolabs). Primer and probe sequences for qRT-PCR were designed using the PrimerExpress v3.0 software (Life Technolo- gies) and synthesized by IDT. Because primers did not include the minor grove binder, the probe Tm parameter was adjusted to be 10˚C higher than the primer Tm values. Analyses for heterodimer and homodimer formation and secondary structure were done using IDT’s OligoAnaly- zer 3.1, and BLAST was used to examine the potential for non-target site primer/probe annealing. Specificity of primers and probes was subsequently verified by analy- zing qRT-PCR products on agarose gels. Probes were la- beled with the 5’ reporter dye 6-FAM and the 3’ quen- cher Iowa Black FQ. qRT-PCR was performed on a StepOne Real Time PCR System (Life Technologies) in 20 μl reaction volumes consisting of 1× TaqMan Gene Expression Master Mix, 900 nM forward and reverse pri- mers, 250 nM probe, and 500 ng of cDNA. Thermocy- cling conditions were: 95˚C for 10 min, followed by 40 cycles of 95˚C for 15 sec and 60˚C for 60 sec. Amplifi- cation efficiency of each primer and probe set was de- termined using a standard curve. The five dilutions of the standard curve were tested in triplicate and were serially diluted five-fold from a starting quantity of 125 ng/μl. In each experiment, samples were run in duplicate and two internal controls were used to normalize results [20]. The genes used for normalization were rpoA (CC_1272), which encodes the RNA polymerase alpha subunit, and secA (CC_3068), which encodes the pre-protein translo- case subunit SecA. These controls were chosen based on expected constitutive expression under all conditions us- ed, and because their functions are unrelated to the genes under investigation. In addition, raw RNA extract and CB15 genomic DNA were included to detect the pres- ence of contaminating genomic DNA. Threshold levels for each gene were adjusted to con- trol for inter-assay variation and the relative expression of each gene was determined with the comparative CT method [21]. The CTs for each sample and target were first obtained by normalizing the threshold cycle number [CT] to the mean CT of constitutively expressed rpoA and secA genes. Relative quantification (RQ) values were cal- culated using the equation RQ = (1 + E)−CT, where E is the amplification efficiency of each target determined by a standard curve and CT = CT reference − CT sample [22]. 2.6. RT-PCR of Hexuronate Metabolism Genes To determine whether closely spaced genes involved in galacturonate metabolism are encoded on the same tran- script, reverse transcriptase-mediated PCR was employ- ed for the following gene pairs: CC_1487-1488 (uxaB and uxaA), CC_1490-1491 (uxaC and kduI), and CC_1495-1496 (kdgA and kdgK). Primers were designed with the forward primer annealing 150 - 300 bp upstream of the 3’ end of the first gene, and the reverse primer an- nealing in the coding region of the putative linked down- stream gene. As a control, primers were designed for the Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 66 CC_3040-3041 gene pair (fliI and fliJ), which are known to be co-transcribed in a polycistronic mRNA. RNA sam- ples were prepared as described above and used as a template for cDNA synthesis. CB15 genomic DNA was included as a positive control. Templates were diluted to 100 ng/μl and PCR was performed in 25 μl reaction vo- lumes containing 1× Green GoTaq Buffer (Promega), 0.25 mM dNTPs, 1 μm each forward and reverse primer, 0.75 U GoTaq DNA Polymerase (Promega) and 1 μl tem- plate. Thermocycling conditions were: 95˚C for 2 min, followed by 30 cycles of 95˚C for 15 sec, 60˚C for 25 sec, and 72˚C for 50 sec, with a final 5 min extension at 72˚C. Products were visualized by agarose gel electrophoresis using 0.9% agarose, and the gels were subsequently stain- ed with ethidium bromide. 3. RESULTS 3.1. Growth of C. crescentus on Hexuronates Analysis of the C. crescentus CB15 genome sequence suggested that it encodes enzymes for the degradation of several major plant polysaccharides, including cellulose, xylan, lignin, glucan, and pectin [8]. To investigate pec- tin and hexuronate utilization, growth of C. crescentus was examined using pectin, galacturonate, and glucuronate as sole carbon sources. (The D-isomer of all monosaccha- rides was used in this work.) Pectin produced mixed re- sults. Under conditions in which glucose produced robust colonies in 3 days, pectin derived from apple peel yield- ed small colonies in 6 days, and pectin derived from cit- rus fruit did not support visible growth. Pectin from dif- ferent plant sources can differ significantly in structure and digestibility [1], but the results with apple peel pec- tin show that C. crescentus is capable of consuming at least some forms of this polymer. The core pectin chain is comprised of alpha-1,4-linked galacturonate. Pure ga- lacturonate (10 mM) supported growth of C. crescentus on plates, with small colonies visible after 4 - 5 days. In liquid M2 medium, the log phase generation time on ga- lacturonate was approximately 4.3 hours, roughly twice as long as doubling times on glucose (2.0 h) or xylose (2.3 h). Adaptation to growth on galacturonate was slow; wild-type cells transitioning from glucose to galacturo- nate in M2 medium experienced a prolonged lag phase of 24 h or more before initiating exponential growth, rough- ly three-fold longer than the lag phase in transitioning from glucose to xylose. Growth on glucuronate was com- parable to growth on apple pectin, suggesting that C. cre- scentus “prefers” galacturonate over glucuronate. 3.2. Genetic Characterization of Galacturonate Utilization In bacteria, pectate (demethylated pectin) can be cleaved by pectatelyase and/or exo-poly-galacturonidase to gen- erate oligo-galacturonides and galacturonate for import to the cytoplasm [2]. Two bacterial pathways have been identified for galacturonate metabolism [3]: the hexuro- nate isomerase pathway studied extensively in E. coli and E. chrysanthemi, and the oxidative pathway active in Ag- robacterium tumifaciens and certain Pseudomonas spe- cies. Although C. crescentus is much more closely relat- ed to A. tumifaciens (an alpha-Proteobacterium) than to E. coli or E. chrysanthemi (gamma-Proteobacteria) [23], the C. crescentus genome contains a set of genes encoding the gamma-Proteobacterial hexuronatei somerase pathway, and lacks a close relative of the galacturonate dehydroge- nase enzyme that is key to the A. tumifaciens pathway [24]. The genomic region containing the genes of the pu- tative C. crescentus hexuronate metabolism pathway is shown in Figure 1. This gene cluster is likely organized as several operons, based on orientation and gene spac- ing (Figure 1(A)). Reverse transcriptase-mediated PCR confirmed that CC_1488 and CC_1487 are transcribed on the same mRNA, as are CC_1490-1491-1492, and CC_1495-1496 (data not shown). Figure 1(B) shows the hexuronate isomerase pathway enabled by the products of this gene cluster [3,5]. The penultimate step generates 2- keto-3-deoxy-6-pho spho-D-gluconate, an intermediate shar- ed with the Entner-Doudoroff pathway, the sole route of glucose catabolism in C. crescentus [11,25]. The final step, catalyzed by keto-deoxyphosphogluconate aldolase (KdgA, CC_1495 gene product), yields pyruvate and gly- ceraldehyde-3-phosphate. We have shown previously that CC_1495 is necessary for glucose metabolism [11]. To determine whether genes in this cluster are neces- sary for growth on galacturonate, a series of mutant strains were constructed and tested for growth on M2 media using galacturonate as sole carbon and energy source (Table 1). As controls, the mutant strains were also tested for growth on glucose and xylose. Strains containing mutations in the CC_1487 (uxaB), CC_1488 (uxaA), CC_1490 (uxaC), and CC_1495 (kdgA) genes were incapable of growth on galacturonate. The kdgA disruption mutant, like the strains with point mutations in kdgA investigated previously, is also incapable of growth on glucose, as the encoded enzyme catalyzes an essential step in the Entner-Doudoroff pathway. Mutations in genes encoding enzymes catalyzing “upstream” reactions in the Entner-Doudoroff pathway (CC_2054-2057) [11] block- ed glucose utilization, but had no effect on galacturonate metabolism. The uxaB, uxaA, and uxaC mutants grow normally on glucose and xylose, and the only growth de- fects relative to wild-type C. crescentus apparent on Bi- olog Phenotype Microarrays were with galacturonate and glucuronate. (Pectin did not support significant growth of C. crescentus in this assay.) Strains containing mutations in CC_1491 (kd uI) and CC_1492 (kduD) grew normally on galacturonate, consistent with their inferred role in the Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 Copyright © 2013 SciRes. 67 Figure 1. A putative pathway for D-galacturonate degradation in C. crescentus. A)The CC_1487–1509 genome region in C. cres- centus CB15. The CC_1494 gene predicted in the original C. crescentus genome annotation is shown in white. The putative CC_1494 ORF (128 codons) has no significant relatives in GenBank. The CC_1497 to CC_1507 region is not shown [hash marks] as their predicted functions do not appear to be connected to hexuronate utilization; B) Proposed C. crescentus pectin/hexuronate deg- radation pathway.Polygalacturonides derived from pectin are degraded to yield monomeric galacturonate or 5-keto-4-deoxyuronate, which are processed through parallel pathways merging at 2-keto-3-deoxygluconate (KDG). The C. crescentus genes hypothesized to encode the enzyme responsible for each step in the pathway are indicated. Gene names are based on homology to genes of known function in E. coli and E. chrysanthemi. Ta b l e 1 . Growth of C. crescentus strains containing mutations in genes encoding putative enzymes of the galacturonate metabolic pathway. “−” indicates that no individual colonies formed on M2 minimal salts agar with the indicated sole carbon and energy source within 7 days. “±” indicates that colonies less than half the size of the wild-type control after 7 days. “+” indicates colonies that formed at a similar rate and size to wild-type. Growth with sole carbon/energy source: Disrupted gene Annotated function Galacturonate Glucose Xylose CC_1487 uxaB D-tagaturonate reductase − + + CC_1488 uxaA D-altronate dehydratase − + + CC_1489 humR Transcriptional regulator, LacI-GalR family + + + CC_1490 uxaC Glucuronate isomerase − + + CC_1491 kduI 5-keto-4-deoxyuronate isomerase + + + CC_1492 kduD 2-deoxy-D-gluconate 3-dehydrogenase + + + CC_1493 ppc Phosphoenolpyruvate carboxylase − − + CC_1495 kdgA 2-keto-3-deoxyphosphogluconate aldolase − − + CC_1496 kdgK 2-dehydro-3-deoxyglucokinase ± ± + parallel branch that processes 5-keto-4-deoxyuronate, a product of pectate lyase. The CC_1496 mutant strain grew slower on galactu- ronate relative to wild-type, but still formed detectable colonies, indicating that this gene product is important but not essential for galacturonate metabolism. We des- ignated CC_1496 as kdgK, based on its genome location and weak sequence similarity to KdgK (2-keto-3-deoxy- gluconate kinase) gene products, but the knockout results suggest that it may not be the only gene product able to carry out this phosphorylation reaction. The ppc gene (CC_1493) is necessary for growth of C. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 68 crescentus on glucose and galactose [11], and was found here to be essential for growth on galacturonate. CC_ 1493 encodes phosphoenolpyruvate (PEP) carboxylase, which catalyzes the addition of CO2 to PEP to generate oxaloacetate. During growth on minimal media with glu- cose or galacturonate as the sole carbon input, PEP car- boxylase may serve an anaplerotic function, replenishing oxaloacetate levels to allow the TCA cycle to continue as intermediates are withdrawn for use in other biosynthetic pathways. 3.3. Regulation of Galacturonate Metabolism To determine whether expression of genes implicated in galacturonate metabolism is induced in response to ga- lacturonate in the culture medium,the regions upstream of uxaA (CC_1488) and uxaC (CC_1490) were inserted next to a promoterless lacZ reporter on a low copy-num- ber plasmid vector. β-galactosidase activity was assayed in wild-type C. crescentus grown in PYE medium [15] before and after exposure to 10 mM galacturonate, glu- curonate, or other sugars. In plain PYE, promoter activity was very low in both constructs (PuxaA = 14 ± 24 Miller units, PuxaC = 54 ± 29). In the presence of galacturonate, promoter activity increased dramatically (PuxaA = 239 ± 139, 17-fold increase; PuxaC = 1150 ± 214, 21-fold in- crease). Subsequent analysis by quantitative real-time PCR (qRT-PCR) showed that both promoters were fully induced within 5 minutes of exposure to galacturonate. PuxaA was relatively weak compared to PuxaC, but the be- havior of the two promoters was qualitatively similar. Glucuronate also induced expression of both promoters, though somewhat less effectively than galacturonate (PuxaA = 15-fold increase; PuxaC = 5-fold increase). Nei- ther glucose, galactose, or xylose induced either pro- moter, nor did they prevent induction of these promoters by galacturonate. Expression of these promoters was also examined in cultures grown in M2 medium, with similar results (data not shown). The CC_1489 gene, located in the midst of the hex- uronate metabolism gene cluster, encodes a member of the LacI repressor family. The polypeptide sequence en- coded by CC_1489 is similar to C. crescentus XylR (CC_3065; 38% identity over 438 amino acids with CC_1489), which controls a gene set for xylose metabo- lism [11,13]. To determine whether CC_1489 controls genes involved in galacturonate metabolism, an integra- tive disruption was constructed. The disruption mutant exhibited a log phase growth rate virtually identical to wild-type on M2 minimal medium supplemented with 10 mM glucose, xylose, galacturonate, or glucuronate, but transitioned more quickly to logarithmic growth when transferred from glucose to galacturonate or glucuronate. When expression of the uxaA and uxaC promoters (PuxaA and PuxaC, respectively) was examined in the CC_1489 disruption mutant, both became highly active under non- inducing conditions, exceeding the activity of the promo- ters in wild-type CB15 in the presence of galacturonate (Table 2 line 1; PuxaA results not shown). In the CC_1489 mutant background, addition of galacturonate did little to increase promoter activity beyond that in plain PYE. These observations are consistent with the CC_1489 pro- duct acting as a repressor of PuxaA and PuxaC in the ab- sence of galacturonate. We therefore have designated the CC_1489 gene product HumR, for hexuronate metabo- lism regulator. MEME [26] was used to search for conserved se- quences in the intergenic regions of the CC_1487-1496 cluster, to identify candidate binding sites for potential regulatory factors. The annotated CC_1494 coding re- gion was included in the search, since this small ORF is not conserved at the protein level in any other species and we suspect that it may not actually encode a poly- peptide. A conserved 14 bp sequence (TGACACC| GGTTACC) was identified upstream of the uxaA, uxaC, and kdgA genes in the C. crescentus genome, and in simi- lar locations upstream of these genes in C. henricii K31, Tabl e 2. Regulation of PuxaC-lacZ expression by HumR and galacturonate. Promoter constructs with identical 5’ and 3’ ends and se- quences, save for the mutations shown, were inserted upstream of a promoterless lacZ gene in the low copy number pRKlac290 re- porter vector. The resulting β-galactosidase activities (Miller units) are shown in the table. CB15 (wild-type) humR disruption Promoter Operator PYE PYE + gal Ratio (+/− gal) PYE PYE + gal Ratio (+/− gal) PuxaC TGACACCGGTTACC 52 ± 28 1090 ± 165 21 3200 ± 54 4900 ± 498 1.5 Mut A TGACAGAGGTTACC 3310 ± 208 3540 ± 374 1.1 3610 ± 132 3160 ± 177 0.9 Mut B TGAGGCCGGTTACC 4810 ± 891 4520 ± 578 0.9 4970 ± 191 4410 ± 112 0.9 Mut C TGACACCTAGTACC 2810 ± 692 2580 ± 287 0.9 2500 ± 280 2250 ± 289 0.9 Mut D TGACACCGGTTTCA 43 + 15 85 ± 4 2.0 −1 −1 −1 1Promoter activity was not detectable, as measured β-galactosidase activity was at or below background levels for the parental strain. Because of this, some activity ratios could not be calculated for this construct. Copyright © 2013 SciRes. OPEN ACCESS 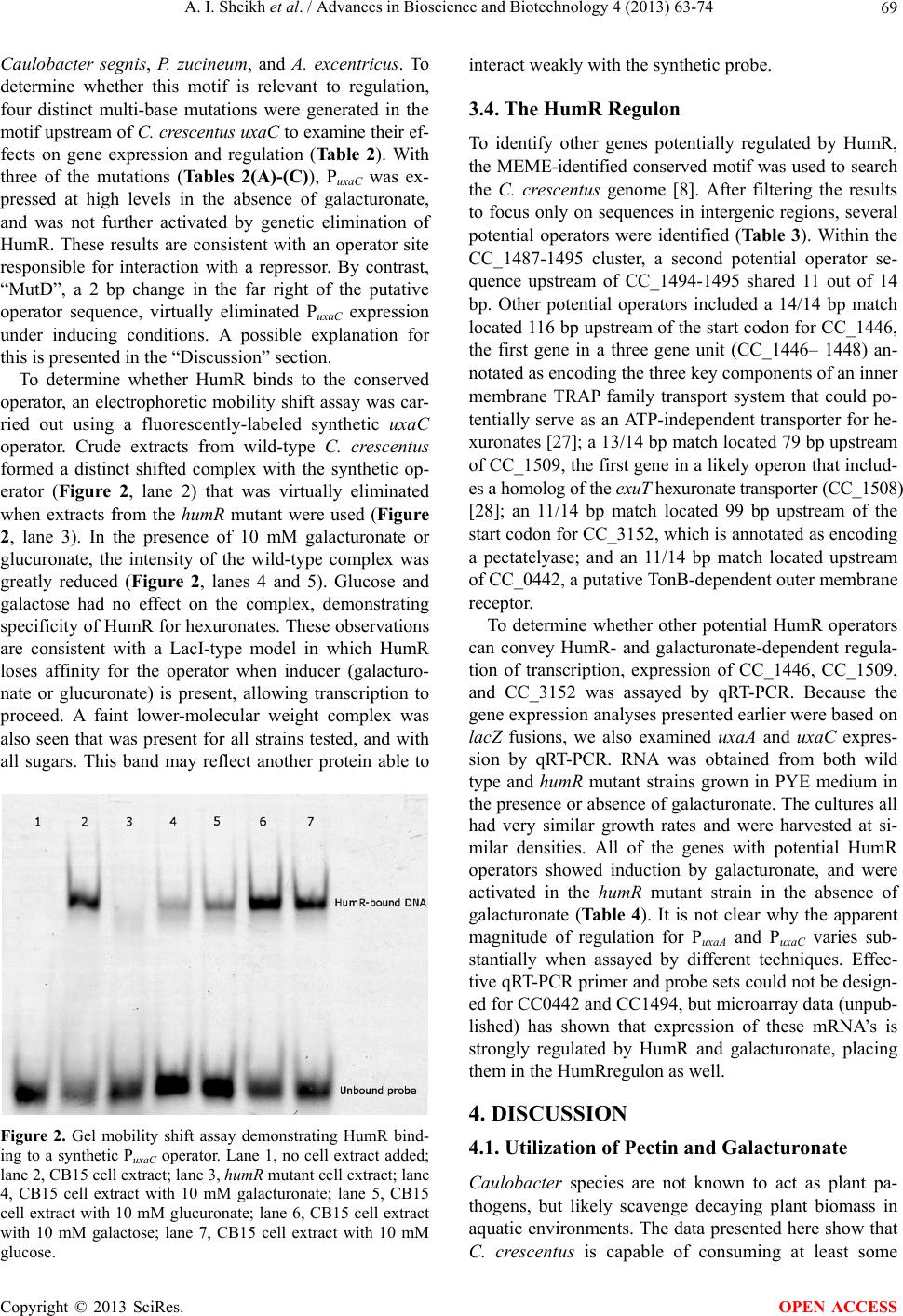 A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 69 Caulobacter segnis, P. zucineum, and A. excentricus. To determine whether this motif is relevant to regulation, four distinct multi-base mutations were generated in the motif upstream of C. crescentus uxaC to examine their ef- fects on gene expression and regulation (Ta b l e 2). With three of the mutations (Tables 2(A)-(C)), PuxaC was ex- pressed at high levels in the absence of galacturonate, and was not further activated by genetic elimination of HumR. These results are consistent with an operator site responsible for interaction with a repressor. By contrast, “MutD”, a 2 bp change in the far right of the putative operator sequence, virtually eliminated PuxaC expression under inducing conditions. A possible explanation for this is presented in the “Discussion” section. To determine whether HumR binds to the conserved operator, an electrophoretic mobility shift assay was car- ried out using a fluorescently-labeled synthetic uxaC operator. Crude extracts from wild-type C. crescentus formed a distinct shifted complex with the synthetic op- erator (Figure 2, lane 2) that was virtually eliminated when extracts from the humR mutant were used (Figur e 2, lane 3). In the presence of 10 mM galacturonate or glucuronate, the intensity of the wild-type complex was greatly reduced (Figure 2, lanes 4 and 5). Glucose and galactose had no effect on the complex, demonstrating specificity of HumR for hexuronates. These observations are consistent with a LacI-type model in which HumR loses affinity for the operator when inducer (galacturo- nate or glucuronate) is present, allowing transcription to proceed. A faint lower-molecular weight complex was also seen that was present for all strains tested, and with all sugars. This band may reflect another protein able to Figure 2. Gel mobility shift assay demonstrating HumR bind- ing to a synthetic PuxaC operator. Lane 1, no cell extract added; lane 2, CB15 cell extract; lane 3, humR mutant cell extract; lane 4, CB15 cell extract with 10 mM galacturonate; lane 5, CB15 cell extract with 10 mM glucuronate; lane 6, CB15 cell extract with 10 mM galactose; lane 7, CB15 cell extract with 10 mM glucose. interact weakly with the synthetic probe. 3.4. The HumR Regulon To identify other genes potentially regulated by HumR, the MEME-identified conserved motif was used to search the C. crescentus genome [8]. After filtering the results to focus only on sequences in intergenic regions, several potential operators were identified (Ta b l e 3 ). Within the CC_1487-1495 cluster, a second potential operator se- quence upstream of CC_1494-1495 shared 11 out of 14 bp. Other potential operators included a 14/14 bp match located 116 bp upstream of the start codon for CC_1446, the first gene in a three gene unit (CC_1446– 1448) an- notated as encoding the three key components of an inner membrane TRAP family transport system that could po- tentially serve as an ATP-independent transporter for he- xuronates [27]; a 13/14 bp match located 79 bp upstream of CC_1509, the first gene in a likely operon that includ- es a homolog of the exuT hexuronate transporter (CC_1508) [28]; an 11/14 bp match located 99 bp upstream of the start codon for CC_3152, which is annotated as encoding a pectatelyase; and an 11/14 bp match located upstream of CC_0442, a putative TonB-dependent outer membrane receptor. To determine whether other potential HumR operators can convey HumR- and galacturonate-dependent regula- tion of transcription, expression of CC_1446, CC_1509, and CC_3152 was assayed by qRT-PCR. Because the gene expression analyses presented earlier were based on lacZ fusions, we also examined uxaA and uxaC expres- sion by qRT-PCR. RNA was obtained from both wild type and humR mutant strains grown in PYE medium in the presence or absence of galacturonate. The cultures all had very similar growth rates and were harvested at si- milar densities. All of the genes with potential HumR operators showed induction by galacturonate, and were activated in the humR mutant strain in the absence of galacturonate (Ta bl e 4 ). It is not clear why the apparent magnitude of regulation for PuxaA and PuxaC varies sub- stantially when assayed by different techniques. Effec- tive qRT-PCR primer and probe sets could not be design- ed for CC0442 and CC1494, but microarray data (unpub- lished) has shown that expression of these mRNA’s is strongly regulated by HumR and galacturonate, placing them in the HumRregulon as well. 4. DISCUSSION 4.1. Utilization of Pectin and Galacturonate Caulobacter species are not known to act as plant pa- thogens, but likely scavenge decaying plant biomass in aquatic environments. The data presented here show that C. crescentus is capable of consuming at least some Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 70 Ta bl e 3 . Possible HumR operator sequences upstream of genes potentially involved in galacturonate acquisition, transport, and de- gradation. The sequences shown are all in intergenic regions. See “Results” section for more on predicted functions of each gene. Adjacent gene Putative operator sequence Orientation of operator relative to promoterDistance from start of operator to translational start [bp] CC_1488 TGA CAC CGG TTA CC + 69 CC_1490 TGA CAC CGG TTA CC + 75 CC_1495 TGA CAC CGG TTA CC − 385 CC_1495 TGA CAC CGG TGG CA + 4351 CC_0442 TGA CAC CGG TGG CA + 129 CC_1446 TGA CAC CGG TTA CC + 115 CC_1509 TGA CAC CGG TTT CC + 78 CC_3152 TGA CAC CGG TGGAC + 96 1Distance to the annotated CC_1494 translational start is 44 bp. As described in the legend to Figure 1, the legitimacy of CC-1494 as a functional coding region is uncertain. Tab le 4. qRT-PCR analysis of gene expression and regulation by HumR and galacturonate. Strains used were wild-type CB15, and a humR disruption mutant in the CB15 background. Cul- tures were grown in PYE medium, and RNA was isolated from cultures at OD600nm ~ 0.3. “Induced” cultures had galacturonate added to 10 mM final concentration 1 hour before the culture reached the target density. See “Results” section for more on predicted functions of each gene. Gene humR−/WT ratio Induced/uninduced ratio CC_1488 53.2 ± 19.8 7.00 ± 2.85 CC_1490 50.0 ± 25.9 6.51 ± 2.79 CC_1446 2.48 ± 0.51 2.01 ± 0.41 CC_1509 111 ± 30 73.8 ± 20.5 CC_3152 14.1 ± 2.2 9.14 ± 1.46 forms of pectin, and grows well on galacturonate, pec- tin’s core monosaccharide. Genetic evidence indicates that C. crescentus metabolizes galacturonate via a pathway starting with hexuronateisomerase, while a parallel path- way processes 5-keto-4-deoxyuronate. The two branches start with distinct pectin breakdown products, probably to accommodate the use of both pectate lyase and poly- galacturonidase. Analysis of the C. crescentus genome shows two putative pectate lyases (CC_3152 and CC_2035) and a putative exo-poly-alpha-galacturonosidase (“exo- PG”, CC_0572), each predicted by SignalP to contain an N-terminal signal peptide whose presence is consistent with secretion. In E. chrysanthemi, exo-PG and pectate- lyase are coordinately regulated, and are both needed for maximally efficient breakdown of pectin [29]. Whether this is true in C. crescentus requires further investigation. The optimization of C. crescentus for utilization of ga- lacturonate, rather than glucuronate, would be sensible if galacturonate, as a component of pectin, is more abundant than glucuronate in niches this freshwater microbe is likely to inhabit. The underlying physiological explana- tion for this adaptation would require comprehensive analysis of transport, regulation, and enzymatic activities. Since glucuronate is capable of inducing the HumR re- gulon, it presumably is imported into the cell, suggesting that the reason for inefficient growth is due to differential metabolism in the cytoplasm. 4.2. Regulation by HumR Expression of several genes implicated in galacturonate utilization in C. crescentus is controlled by HumR, a member of the LacI family of bacterial transcription fac- tors [30]. There are 11 LacI family members encoded in the C. crescentus genome. Other are known to control uptake and/or metabolism of xylose (CC_3065), glucose (CC_2053), and maltose (CC_2284) [11,13,21]. Data presented here indicates that HumR binds to cognate operators in the absence of galacturonate, and interferes with effective RNA polymerase interaction with the re- spective promoter. The data are consistent with galactu- ronate binding to HumR, stabilizing a conformation with reduced affinity for operator DNA and releasing it from the promoter region to allow gene expression. Glucuro- nate also appears to productively interact with HumR as well, both in vitro and in vivo. HumR functions similarly to the C. crescentus xylose repressor, XylR [13]. The XylR regulon includes a xylose transporter and an operon encoding enzymes for xylose metabolism, as well as putative secretive xylanases and arabinosidases [11]. The HumR operator (TGACACC| GGTTACC) and XylR operator (TGTTAGC|GCTACCA) sequences are the same length (14 bp), and identical at 7 out of 14 positions [13]. Given that LacI-type repressors bind as dimers, dyad symmetry is expected in the opera- tor. All of the identified HumR operators are identical in Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 71 one half-site (TGACACC), which we suspect may be the optimal binding sequence for a HumR subunit. We have been unable to experimentally determine the 5’ end of a HumR-regulated transcript, but alignment of PuxaC se- quences from the Caulobacter group (Figure 3(a)) re- veals additional conserved sequences flanking the op- erator that match the −10 and −35 elements of Pxyl (Fig- ure 3(b)), for which the 5’ end is known. In Pxyl, the XylR operator is positioned between the −35 and −10 regions, overlapping the predicted downstream contact site for the σ submit of RNA polymerase. If the aligned sequences represent the same functional promoter ele- ments in Pxyl and PuxaC, then bound HumR would like- wise overlap the −10 region, allowing it to block critical contacts with the σ subunit. If that is true, the sequence of the right half-site of the HumR operator in PuxaC is probably under dual selection for binding HumR and σ. This is consistent with the effect of OpMutD, a 2 bp change on the far right side of the operator (Table 2) that actually makes the sequence of the HumR operator half site more similar to the left side consensus, but changes the putative −10 region away from that shared with Pxyl. Table 2 shows that promoter activity was dramatically reduced by this mutation, even under conditions where HumR binding should be eliminated, supporting the hy- pothesis that the operator overlaps with a critical pro- moter element. As with the XylR regulon, some HumR-regulated pro- moters show mild reductions in inducibility when glu- cose is present, but glucose does not preclude induction of XylR or HumR-regulated promoters. There are no sig- nificantly conserved sequence elements in these promot- ers other than the identified operators, so direct involve- ment of additional glucose-responsive transcription factors such as CAP (E. coli) or CcpA (Bacillus subtilis kduID operon [31]) seems unlikely. As an oligotroph adapted to low-nutrient habitats, there may be minimal reward for C. crescentus to prioritize glucose utilization at the expense of other available sugars. The HumR regulon in C. crescentus includes at least five inferred operons (CC_1488-1487, CC_1490-1491- 1492, CC_1495-1496, CC_1509-1508 and CC_1446- 1448) and two singleton genes (CC_3152 and CC_0442). Based on annotated functions, the components of the HumR/galacturonate regulon fit the pattern previously established for the XylR/xylose regulon [11,13], and for glucose-dependent gene expression [11], in that they all include extracellular polysaccharide-active enzymes, TonB- dependent outer membrane receptors (TBDR’s), inner membrane transport systems, and a cytoplasmic catabolic pathway. Thus, each repressor likely controls a complete system for extracellular polysaccharide degradation, up- take of breakdown products, and intracellular catabolism of monosaccharides to yield energy and central metabo- lites. The use of TBDR’s for scavenging polysaccharide degradation products is shared by phytopathogenic Xan- thomonas species, and Blanvillain et al. [32] recognized that TBDR’s tend to be genetically clustered with “carbo- hydrate active enzymes” and inner membrane transport systems to create “CUT” (carbohydrate-utilization) loci. Caulobacter crescentus apparently did not maintain tight genetic linkage of CUT systems over evolutionary time, but still coordinates expression of components through repressors that recognize specific monosaccharides as in- duction signals. In E. coli and Erwinia chrysanthemi, pectin degrada- tion and hexuronate utilization are primarily controlled by the ExuR, UxuR, and KdgR transcription factors spe- (a) (b) Figure 3. The uxaC promoter and HumR operator. a) Alignment of uxaC promoter region in C. crescentus and related species (C. henricii K31, C. segnis and P. zucineum). Distance is shown to the start codon, as transcription start sites could not be determined; b) lignment of C. crescentus Pxyl [14] and PuxaC. Numbering for Pxyl is shown relative to the transcription start site [14]. A Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 72 cies, which are members of the GntR class of transcrip- tion factors [5,33]. As in C. crescentus, the hexuronate- regulon in E. chrysanthemi includes the hexuronate iso- merase pathway, extracellular enzymes for pectin break- down, and inner membrane transporters (ExuT) [34,35]. Neither C. crescentus, its close relatives, or the Xantho- monas species encode KdgR-like genes. Instead, humR is embedded within the hexuronate metabolic gene cluster in the genomes of close C. crescentus relatives (C.segnis, C. henricii strain K31, Maricaulis maris, and Asticcacau- lisexcentricus). Phenylobacterium zucineum has a modi- fied cluster, with the HumR-regulated operon homolo- gous to CC_1509-1508 (exuT) inserted between humR and uxaC. Based on the presence of HumR operators, the HumR regulon is largely conserved in these species, with the exception of CC_1446 and CC_3152, which are uni- que to C. crescentus. Xanthomonas campestris and X. axonopodis, which like Erwinia chrysanthemi are gam- ma-Proteobacterial phytopathogens [36], may also utilize a HumR-type regulatory system, as their genomes have a HumR homolog located immediately adjacent to kduI and kd uD, and possible operators located upstream of the kduID operon and the kdgK gene (not shown). Genomic clustering of the genes for hexuronate me- tabolism (CC_1487 – CC_1496) undoubtedly has facili- tated their co-regulation, and potential to be horizontally transferred as a functional unit. The breakdown of syn- teny outside of this gene cluster is consistent with the hypothesis that horizontal transfer introduced this path- way into the Caulobacter lineage. The inclusion of a sim- ple transcription control system in the cluster was likely beneficial in that it ensured appropriate regulation of the pathway in a new host. Price et al. [37] found in E. coli that most “neighbor regulators”—transcription factors that are genetically adjacent to the genes they regulate— were likely acquired by horizontal gene transfer with their target genes. A similar pattern applies to other-LacI- type repressors in the Caulobactergroup, since the hex- uronate, glucose, and maltose-responsive repressors are clustered with genes they control in the C. crescentus ge- nome, and XylR is clusted with the xyl operon in the C. henricii K31 genome. Such clustering should help in elu- cidating the regulatory function of the many uncharac- terized LacI homologs in the Caulobacter group. 5. CONCLUSION Caulobacter crescentus encodes genes (CC_1487 to CC_1495) necessary for metabolism of galacturonate through the hexuronate isomerase pathway. A conserved 14 bp operator upstream of three operons within this clu- ster binds HumR, a LacI-type repressor encoded by the CC_1489 gene. HumR represses these promoters in the absence of galacturonate, and releases the operator in its presence. Several additional genes likely involved in pectin degradation and hexuronate transport also appear to be part of the HumR regulon, which joins a growing roster of plant polysaccharide utilization systems con- trolled by LacI repressor homologs in C. crescentus. 6. ACKNOWLEDGEMENTS This work was supported by U.S. National Science Foundation grant MCB-0818934 to CS, and by financial support for the BIOL 176 Re- combinant DNA Technology course from Santa Clara University. We gratefully acknowledge the help of numerous BIOL 176 students who contributed to this project through preparation of reagents and helpful discussions. We also thank Virginia Kalogeraki, Eduardo Abeliuk, Bo Zhou, and Harley McAdams for assistance with microarray experi- ments and discussion of data. REFERENCES [1] Mohnen, D. (2008) Pectin structure and biosynthesis. Current Opinion in Plant Biology, 11, 266-277. http://dx.doi.org/10.1016/j.pbi.2008.03.006 [2] Abbott, D.W. and Boraston, A.B. (2008) Structural biolo- gy of pectin degradation by Enterobacteriaceae. Microbi- ology and Molecular Biology Reviews, 72, 301-316. http://dx.doi.org/10.1128/MMBR.00038-07 [3] Richard, P. and Hilditch, S. (2009) D-galacturonic acid catabolism in microorganisms and its biotechnological re- levance. Applied Microbiology and Biotechnology, 82, 597-604. http://dx.doi.org/10.1007/s00253-009-1870-6 [4] Yadav, S., Yadav, P., Yadav, D. and Yadav, K. (2009) Pectin lyase: A review. Process Biochemistry, 44, 1-10. http://dx.doi.org/10.1016/j.procbio.2008.09.012 [5] Hugouvieux-Cotte-Pattat, N. and Robert-Baudouy, J. (1987) Hexuronate catabolism in Erwinia chrysanthemi. Journal of Bacteriology, 169, 1223-1231. [6] Entcheva-Dimitrov, P. and Spormann, A.M. (2004) Dy- namics and control of biofilms of the oligotrophic bacte- rium Caulobacter crescentus. Journal of Bacteriology, 186, 8254-8266. http://dx.doi.org/10.1128/JB.186.24.8254-8266.2004 [7] Poindexter, J. (1964) Biological properties and classifica- tion of the Caulobacter group. Bacteriology Reviews, 28, 231-295. [8] Nierman, W.C., Feldblyum, T.V., Laub, M.T., Paulsen, I.T., Nelson, K.E., Eisen, J., et al. (2001) Complete ge- nome sequence of Caulobacter crescentus. Proceedings of the National Academy of Sciences of USA, 98, 4136-4141. http://dx.doi.org/10.1073/pnas.061029298 [9] Mannisto, M.K., Tiirola, M.A., Salkinoja-Salonen, M.S., Kulomaa, M.S. and Puhakka, J.A. (1999) Diversity of chlo- rophenol-degrading bacteria isolated from contaminated boreal groundwater. Archives of Microbiology, 171 , 189- 197. http://dx.doi.org/10.1007/s002030050698 [10] Luo, Y., Xu, X., Ding, Z., Liu, Z., Zhang, B., Yan, Z., Sun, J., Hu, S. and Hu, X. (2008) Complete genome of Phenylobacterium zucineum, a novel facultative intracel- Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 73 lular bacterium isolated from human erythroleukemia cell line K562. BioMed Central Genomics, 9, 386. http://dx.doi.org/10.1186/1471-2164-9-386 [11] Hottes, A.K., Meewan, M., Yang, D., Arana, N., Romero, P., McAdams, H.H. and Stephens, C. (2004) Transcrip- tional profiling of Caulobacter crescentus during growth on complex and minimal media. Journal of Bacteriology, 186, 1448-1461. http://dx.doi.org/10.1128/JB.186.5.1448-1461.2004 [12] Stephens, C., Christen, B., Fuchs, T., Sundaram, V., Wa- tanabe, K. and Jenal, U. (2007) Genetic analysis of a no- vel pathway for D-xylose metabolism in Caulobacter cre- scentus. Journal of Bacteriology, 189, 2181-2185. http://dx.doi.org/10.1128/JB.01438-06 [13] Stephens, C., Christen, B., Watanabe, K., Fuchs, T. and Jenal, U. (2008) Regulation of D-xylose metabolism in Caulobacter crescentus by a LacI-type repressor. Journal of Bacteriology, 189, 8828-8834. http://dx.doi.org/10.1128/JB.01342-07 [14] Meisenzahl, A.C., Shapiro, L. and Jenal, U. (1997) Isola- tion and characterization of a xylose-dependent promoter from Caulobacter crescentus. Journal of Bacteriology, 179, 592-600. [15] Ely, B. (1991) Genetics of Caulobacter crescentus. Meth- ods in Enzymology, 204, 372-384. http://dx.doi.org/10.1016/0076-6879(91)04019-K [16] Bochner, B.R. (2003) New technologies to assess geno- type-phenotype relationships. Nature Reviews Genetics, 4, 309-314. http://dx.doi.org/10.1038/nrg1046 [17] West, L., Yang, D. and Stephens, C. (2002) Use of the Caulobacter crescentus genome sequence to develop a method for systematic genetic mapping. Journal of Bac- teriology, 184, 2155-2166. http://dx.doi.org/10.1128/JB.184.8.2155-2166.2002 [18] Zuker, M. (2003) Mfold web server for nucleic acid fold- ing and hybridization prediction. Nucleic Acids Research, 31, 3406-3415. http://dx.doi.org/10.1093/nar/gkg595 [19] Stephens, C. and Shapiro, L. (1993) An unusual promoter controls cell-cycle regulation and dependence on DNA replication of the Caulobacter fliLM early flagellar op- eron. Molecular Microbiology, 9, 1169-1179. http://dx.doi.org/10.1111/j.1365-2958.1993.tb01246.x [20] Hruz, T., Wyss, M., Docquier, M., Pfaffl, M., Masanetz, S., Borghi, L., et al. (2011) RefGenes: identification of reliable and condition specific reference genes for RT- qPCR data normalization. BMC Genomics, 12, 156. http://dx.doi.org/10.1186/1471-2164-12-156 [21] Lohmiller, S., Hantke, K., Patzer, S.I. and Braun, V. (2008) TonB-dependent maltose transport by Caulobac- ter crescentus. Microbiology, 154, 1748-1754. http://dx.doi.org/10.1099/mic.0.2008/017350-0 [22] Livak, K.J. and Schmittgen, T.D. (2001) Analysis of rela- tive gene expression data using real-time quantitative PCR and the 2[-Δ Δ C[T]] method]. Methods, 25, 402-408. http://dx.doi.org/10.1006/meth.2001.1262 [23] Williams, K.P., Sobral, B. and Dickerman, A. (2007) A robust species tree for the Alphaproteobacteria. Journal of Bacteriology, 189, 4578-4586. http://dx.doi.org/10.1128/JB.00269-07 [24] Boer, H., Maaheimo, H., Koivula, A., Penttila, M. and Richard, P. (2009) Identification in Agrobacterium tume- faciens of the galacturonic acid dehydrogenase gene. Ap- plied Microbiology and Biotechnology, 86, 901-909. http://dx.doi.org/10.1007/s00253-009-2333-9 [25] Riley, R.G. and Kolodziej, B.J. (1976) Pathway of glu- cose catabolism in Caulobacter crescentus. Microbios, 1, 219-226. [26] Bailey, T., Bodén, M., Buske, F.A., Frith, M., Grant, C.E., Clementi, L., et al. (2009) MEME SUITE: Tools for mo- tif discovery and searching. Nucleic Acids Research, 37, W202-W208. http://dx.doi.org/10.1093/nar/gkp335 [27] Fischer, M., Zhang, Q.Y., Hubbard, R.E. and Thomas, G.H. (2010) Caught in a TRAP: Substrate-binding pro- teins in secondary transport. Trends in Microbiology, 18, 471-478. http://dx.doi.org/10.1016/j.tim.2010.06.009 [28] Haseloff, B.J., Freeman, T.L., Valmeekam, V., Melkus, M.W., Oner, F., Valachovic, M.S. and San Francisco, M.J. (1998) The exuT gene of Erwinia chrysanthemi EC16: Nucleotide sequence, expression, localization, and rele- vance of the gene product. Molecular Plant-Microbe In- teractions, 11, 270-276. http://dx.doi.org/10.1094/MPMI.1998.11.4.270 [29] Collmer, A., Whalen, C., Beer, S. and Bateman, D. (1982) An exo-poly-α-D-galacturonosidase implicated in the re- gulation of extracellular pectatelyase production in Erwi- niachrystanthemi. Journal of Bacteriology, 149, 626-634. [30] Swint-Kruse, L. and Matthews, K. (2009) Allostery in the LacI/GalR family: Variations on a theme. Current Opin- ions in Microbiology, 12, 129-137. http://dx.doi.org/10.1016/j.mib.2009.01.009 [31] Lin, J.-S. and Shaw, G.-C. (2007) Regulation of the kduID operon of Bacillus subtilis by the KdgR repressor and the ccpA gene: Identification of two KdgR-binding sites with- in the kdgR-kduI intergenic region. Microbiology, 153, 701-710. http://dx.doi.org/10.1099/mic.0.2006/002253-0 [32] Blanvillain, S., Meyer, D., Boulanger, A., Lautier, M., Guynet, C., Denanc, Ä.N., Vasse, J., Lauber, E. and Arlat, M. (2007) Plant carbohydrate scavenging through TonB- dependent receptors: A feature shared by phytopathoge- nic and aquatic bacteria. PLoS One, 2, e224. http://dx.doi.org/10.1371/journal.pone.0000224 [33] Hugouvieux-Cotte-Pattat, N., Condemine, G., Nassar, W. and Reverchon, S. (1996) Regulation of pectinolysis in Erwinia chrysanthemi. Annual Review of Microbiology, 50, 213-257. http://dx.doi.org/10.1146/annurev.micro.50.1.213 [34] Rodionov, D.A., Gelfand, M.S. and Hugouvieux-Cotte- Pattat, N. (2004) Comparative genomics of the KdgR re- gulon in Erwinia chrysanthemi 3937 and other gamma- proteobacteria. Microbiology, 150, 3571-3590. http://dx.doi.org/10.1099/mic.0.27041-0 [35] Rodionov, D.A., Mironov, A.A., Rakhmaninova, A.B. and Gelfand, M.S. (2000) Transcriptional regulation of tran- sport and utilization systems for hexuronides, hexuronat- es and hexonates in gamma purple bacteria. Molecular Microbiology, 38, 673-683. http://dx.doi.org/10.1046/j.1365-2958.2000.02115.x Copyright © 2013 SciRes. OPEN ACCESS  A. I. Sheikh et al. / Advances in Bioscience and Biotechnology 4 (2013) 63-74 Copyright © 2013 SciRes. 74 OPEN ACCESS [36] Da Silva, A.C., Ferro, J.A., Reinach, F.C., Farah, C.S., Furlan, L.R., Quaggio, R.B., et al. (2002) Comparison of the genomes of two Xanthomonas pathogens with differ- ing host specificities. Nature, 417, 459-463. http://dx.doi.org/10.1038/417459a [37] Price, M.N., Dehal, P. and Arkin, A. (2008) Horizontal gene transfer and the evolution of transcriptional regula- tion in Escherichia coli. Genome Biology, 9, R4. http://dx.doi.org/10.1186/gb-2008-9-1-r4
|