 Theoretical Economics Letters, 2013, 3, 18-25 http://dx.doi.org/10.4236/tel.2013.35A3003 Published Online October 2013 (http://www.scirp.org/journal/tel) Why Are Children Impatient? Evolutionary Selection of Preferences Junji Kageyama Department of Economics, Meikai University, Urayasu, Japan Email: kagejun@meikai.ac.jp Received August 1, 2013; revised September 1, 2013; accepted September 10, 2013 Copyright © 2013 Junji Kageyama. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT This study aims to explain why children are impatient. Using a biological framework called the life history theory, the study investigates the evolutionary root of time preference, paying particular attention to childhood. The results show that the biologically endowed rate of time preference is equal to the mortality rate not only in adulthood but also in childhood, reflecting the ch ang e in the b iolog ical valu e of su rviv al. Mortality is the b aselin e fo r time pr efer en ce through the entire course of life. These results are consistent with the findings in previous empirical and experimental studies that the discount rate is U-shaped in age, and account for why young children, in particular, are impatient. In addition, the difference in time preference between adults and children provides a biological explanation for the parent-offspring conflict, in which the higher discount rate among children causes parents and their children to disagree over intertem- poral allocation of resources in collective decision-making particularly within the household. Keywords: Time Preference; Value of Survival; Life History Theory; Mortality; Parent-Offspring Conflict 1. Introduction The decision-making process within a household typi- cally involves multiple individuals such as the wife, hus- band and children. As a result, collective decisions made in the household depend on preferences of household members. For example, Dauphin et al. [1] showed that children as well as parents influenc e household econo mic decisions1. Nevertheless, children, who sometimes make up the larger portion of the family, seldom receive much atten- tion in the study of economics. Considering that even an infant is capable of tilting household decisions toward his/her preferred choices using his/her limited but pow- erful strategies, this gap needs to be addressed. As a step toward this objective, this study investigates the intertemporal choice of children, focusing on the cause of their impatience. As found in Bettingera and Slonim [4] and Steinberg et al. [5], patience increases with age during childhood, suggesting that young chil- dren are particularly impatien t2. To be more specific, this study searches for the evolu- tionary root of impatience of children, employing a bio- logical framework. An increasing body of literature uses this method to search for the biological basis of prefer- ences. The basic idea of this literature comes from the bio- logical finding that preferences or, more broadly, geno- types and associated phenotypes (strategies), are the end- products of natural selection. Since preferences that are successful in reproduction spread over the population in the evolutionary time scale, preferences in the current human population can be deduced from the preferences that maximized fitness in the environment where we evolved. Such environment is considered to be the Afri- can savannah where our genus Homo appeared two mil- lion years ago and stayed for most of our history as hunter-gatherers. Human-specific characteristics are con- sidered to have evolved in this ancestral environment. Time preference is a leading topic in this literature. Hansson and Stuart [7] showed that the marginal rate of substitution in utility is equ al to the marginal rate of sub- stitution in fitness, suggesting that resource allocation maximizing utility corresponds to allocation maximizing fitness. Later, Rogers [8] applied this idea to the in- tertemporal allocation of resources, and explained the 1To justify the unitary model in which the unitary decision maker maximizes household utility, we often require restrictive assumptions. The rotten kid theorem (Becker [2]) is such an example. See Bergstrom [3] for details. 2For other psychological studies, see Teuscher and Mitchell [6] for a review. C opyright © 2013 SciRes. TEL  J. KAGEYAMA 19 evolutionary origin of time preference. More recently, technical similarities for studying aging in bio-de- mography and the intertemporal allocation in economics have allowed biologists, demographers and economists to enter this hybrid field, leading to the ex amination of how the time-discounting behavior relates to senescence (So- zou and Seymour [9]) and to intergenerational transfers (Chu et al. [10]), why it is hyperbolic (Robson and Samuelson [11]), the rationale for social discounting (Sozou [12]), how it depends on age (Kageyama [13]), and how it relates to extrinsic mortality (Chowdhry [14]) 3. To investigate the impatience of children, the present study follows this literature and extends Kageyama [13] by incorporating childhood, i.e., the growth period to maturity. In previous studies, Chu et al. [10] built a bio- logical model incorporating the growth period and showed that the impatience relates to the productivity growth in childhood. Similarly, Robson et al. [16] ex- amined the impatience of children, but paid more atten- tion to adulthood and did not explicitly consider the role of the growth period. Despite these differences, both of these previous studies concluded that the impatience of children does not relate to the absolute level of mortality. This pre- sents a sharp contrast to the result that mortality is a major factor associated with time preference in adult- hood. The present study, on the other hand, finds that the impatience of children relates to the mortality rate, as does time preference in adulthood. The same logic ap- plies in both childhood and adulthood. Explaining time preference with evolutionary biology, however, does not negate the relationship between time- discounting behavior and non-biological factors. As Becker and Mulligan [17] argue, social factors such as culture and education affect time discounting. This paper aims to assess the biological basis of time preference, referred to as the “endo wed discount factor” in the abo ve study. The rest of the paper is organized as follows. Section 2 examines the evolutionary optimal strategies using a biological framework. Based on the results in Section 2, Section 3 provides a biological explanation for time preference, and shows that the optimal rate of time pref- erence is equal to the mortality rate in the entire life course. This implies that the biolog ically endowed rate of time preference is U-shaped in age, as is the morta l it y rate, and that children and old adults are, by nature, less pa- tient than young adults. Section 4 concludes. 2. The Model 2.1. The Basic Structure The model in this study is based on life history theory, i.e., an analytical framework in biology to study spe- cies-specific life-history strategies such as the age-tra- jectories of fertility and mortality, presuming that life- history traits are the end-products of natural selection. Technically, it solves for the fitness-maximizing strate- gies under given constraints to deduce species-specific life-history traits4. In particular, as in a standard life-history model, I con- sider a model with the following properties. First, the population is stationary at the upper limit of the carrying capacity. This is to follow the carrying-capacity argu- ment that human population in our evolutionary past was confined by the environmental capacity5. Second, repro- duction is asexual. This is simply to avoid complexities related to matching between females and males. With these specifications, the measure of fitness is given by the expected number of offspring at the begin- ning of life, which is expressed as 0 0Rlxmx dx (1) where l(x) is the survival probability to age x and m(x) is the reproductive output at age x6. With the pressure of natural selection, genotypes and associated phenotypes that generate a higher value of R(0) spread over and fix in the population. Note that, in a stationary population where the p opulation growth rate converges to zero, R(0) is equivalent to the reproductive value at birth and con- verges to one. The lifetime exp ected number of offsp r ing is, ex post, just sufficient to replace the current individual. Survival and reproduction depend on age and the amount of energetic resources respectively invested in. Once consumed, resources are physiologically allocated to either survival investment or reproductive investment. Therefore, given that 0ˆˆ exp d x lxx x (2) where is the mortality rate at age x, the depend- ence of survival and reproduction on resources can be expressed as , wx x and ,x xmxmv where w(x) and v(x) are respec- tively survival and reproductive investments. To avoid unnecessary technical complexity, I assume that both 3Acharya and Balvers [15] examined time preference in an economic framework, assuming that utility captures the effect of consumption on mortality. Their model can also be interpreted as a biological model that assumes that reproductive success solely depends on the length o life. 4See Stearns [18] for the general introduction of life history theory, and Perrin and Sibly [19] for the technic a l i n t roduction. 5It is technically possible to examine the case that the population growth rate takes non-zero constant values. See Taylor et al. [20]. 6Equation (1) does not imply that lifespan is infinite. Copyright © 2013 SciRes. TEL  J. KAGEYAMA 20 investments exhibit diminishing marginal returns, satis- fying , , v, and ,0 wwx x ,0xx ,0 ww wx x mv ,0 vv mvxx 0 where the sub- script indicates a partial derivative, and also that the in- dividual will certainly die without survival investment and will not have any reproductive output without re- productive investment. In addition, reproduction depends on growth invest- ment, z(x), in the earlier stages of life. Investing in growth enhances the reproductive capacity and increases the reproductive efficiency in the later stages. To incorporate this relationship, I focus on determinate growers, i.e. , organisms that stop growing at maturity, as in the case of humans7. Thus, denoting the age of matur- ity by α, we have for all x < α and z(x) = 0 for all vx x . Besides, I assume that ,mvx x can be separated into the age-dependent reproductive capacity, A(x), and the contribution of resources, vx , such that vxAx f ,mvx x . (3) Note that vx ,x x is concave in v(x) as is . In this setting, the development of the re- productive capacity during the growth period can be ex- pressed as mv 0ˆˆˆ 0d x xABxfzx () x (4) where A(0) is the innate reproductive capacity and B(x) is the age-dependent growth efficiency. Here, the contribu- tion of resources to growth follows the same law as the one to reproduction, and, thus, both reproductive and growth investments share the same dx vxzxw . Turning to the budget constraint, the model incorpo- rates resource transfers. The extensive use of resource transfers, such as intergenerational transfers between parents and children and food sharing between house- holds, is one of the most distinctive human characteris- tics8. In this case, the budget constraint does not neces- sarily hold at each point in time, and is given by the life- time budget constraint, 00 lxyx xl x xd (5) where y(x) is the amount of resources that the individual obtains at age x. Note that y(x) is exogenous, and that th e interest rate is equal to zero since keeping resources does not generate any return. With these conditions, we can solve for the optimal allocation of resources, expr essed by v(x), z(x), w(x), and α, using the Lagrangian method. Given the objective function (1), the budget constraint (5), and the Lagran- gian multiplier, λ, the Lagrangian is defined as 0 0 d d Llxmxx lx yxvxzxwxx (6) where l(x), m(x), and A(x) are respectively specified by Equations (2), (3), and (4). Using the Volterra derivative (see Ryder and Heal [24]) to examine the effect of changes in the investments around a particular age x, we obtain the first-order conditions: 0for v LlxAxfvxlxx vx , (7) 0for x< z LBx fzxlx zx , (8) ,0 w Lwx xRxkxlx wx , (9) and 0d Llx yxvxzxwxx 0 (10) where ˆˆˆ dlxf vxx ˆ , (11) ˆˆ ˆˆ d x RxlxAx fvxx , (12) and ˆˆ ˆˆˆ d x kxlx yxvxzxwxx . (13) Here, represents the benefit of an increment in the reproductive capacity, and R(x) and k(x) express the benefits of an increase in survival at age x respectively for reproduction and production. At the same time, the age of maturity is determined by the returns of growth and reproductive investments. These returns are given by Bx fzxx and lxAxf vx , which respectively represent the increase in the future reproduction due to a greater re- productive capacity and the increase in immediate re- production. The individual switches from the growth phase to the reproductive phase when the return for re- production overtakes the one for growth. Therefore, the age of maturity is implicitly given by Bl 7To account for determinate growth, we can, for example, assume that the transition from the growth phase to the reproductive phase, or the other way around, incurs significant costs. This is consistent with the fact that determinate growers go through some sort of metamorphosis at maturity. 8See, e.g., Wiessner [21,22] and Gurven et al. [23] for the importance of food sharing in modern foraging p op ulat ions . A . (14) Copyright © 2013 SciRes. TEL 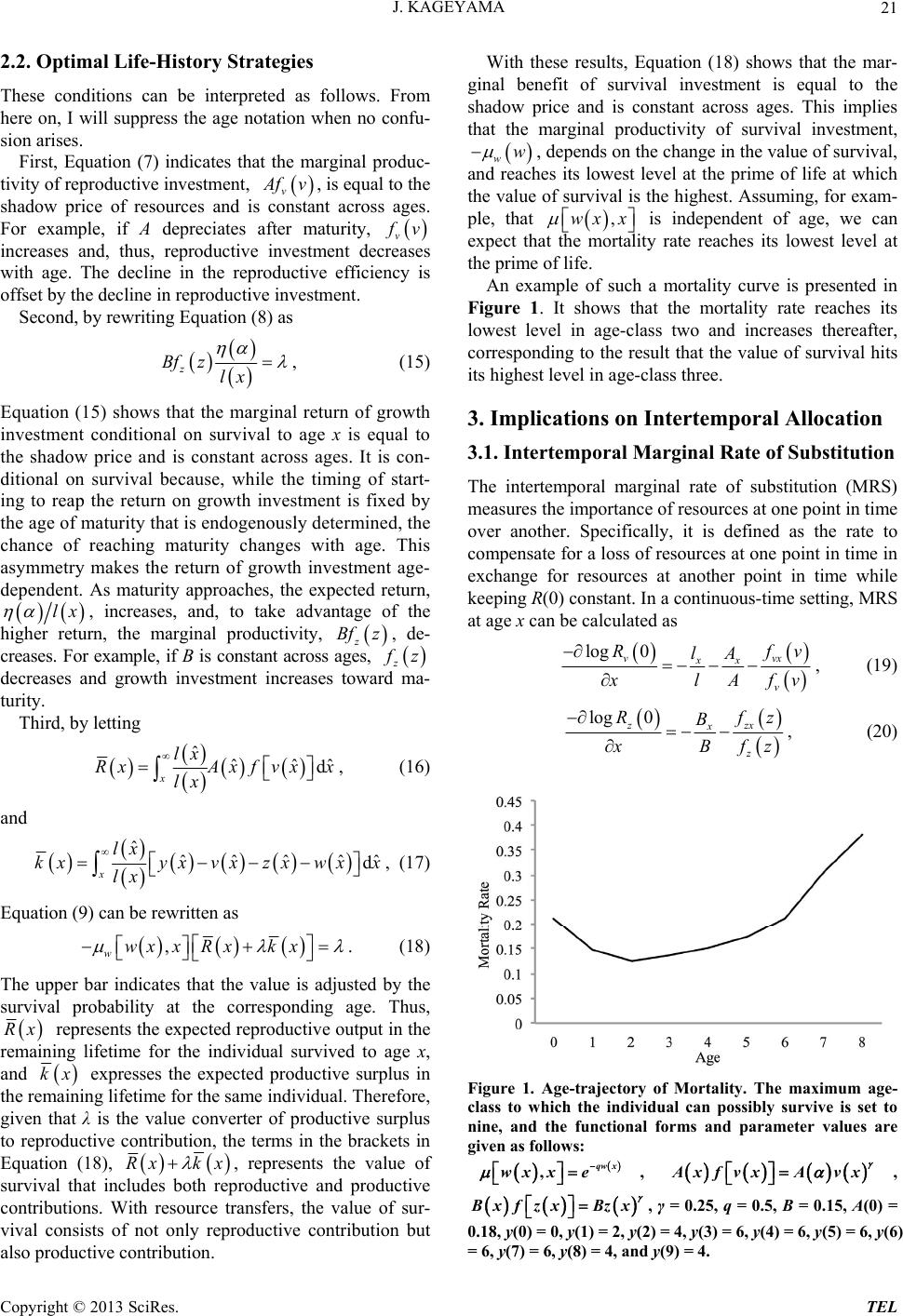 J. KAGEYAMA 21 2.2. Optimal Life-History Strategies These conditions can be interpreted as follows. From here on, I will suppress the age notation when no confu- sion arises. First, Equation (7) indicates that the marginal produc- tivity of reproductive investmen t, v fv, is equal to the shadow price of resources and is constant across ages. For example, if A depreciates after maturity, v v increases and, thus, reproductive investment decreases with age. The decline in the reproductive efficiency is offset by the decline in reproductive investment. Second, by rewriting Equation (8) as z Bfz lx , (15) Equation (15) shows that the marginal return of growth investment conditional on survival to age x is equal to the shadow price and is constant across ages. It is con- ditional on survival because, while the timing of start- ing to reap the return on growth investment is fixed by the age of maturity that is endogenously determined, the chance of reaching maturity changes with age. This asymmetry makes the return of growth investment age- dependent. As maturity approaches, the expected return, lx , increases, and, to take advantage of the higher return, the marginal productivity, z Bfz , de- creases. For example, if B is constant across ages, z z decreases and growth investment increases toward ma- turity. Third, by letting ˆˆˆ d x lx RxAxfvxx lx ˆ , (16) and ˆˆˆˆ ˆ d x lx kxyxvxzx wxx lx ˆ , (17) Equation (9) can be rewritten as , wwxxRxk x . (18) The upper bar indicates that the value is adjusted by the survival probability at the corresponding age. Thus, Rx represents the expected reproductive output in the remaining lifetime for the individual survived to age x, and kx expresses the expected productive surplus in the remaining lifetime for the same individual. Therefore, given that λ is the value converter of productive surplus to reproductive contribution, the terms in the brackets in Equation (18), Rxk x , represents the value of survival that includes both reproductive and productive contributions. With resource transfers, the value of sur- vival consists of not only reproductive contribution but also productive cont ri b ut io n. With these results, Equation (18) shows that the mar- ginal benefit of survival investment is equal to the shadow price and is constant across ages. This implies that the marginal productivity of survival investment, ww , depends on the change in the value of survival, and reaches its lowest level at the prime of life at which the value of survival is the highest. Assuming, for exam- ple, that ,wx x is independent of age, we can expect that the mortality rate reaches its lowest level at the prime of life. An example of such a mortality curve is presented in Figure 1. It shows that the mortality rate reaches its lowest level in age-class two and increases thereafter, corresponding to the result that the value of survival hits its highest level in age-class three. 3. Implications on Intertemporal Allocation 3.1. Intertemporal Marginal Rate of Substitution The intertemporal marginal rate of substitution (MRS) measures the importance of resources at one point in time over another. Specifically, it is defined as the rate to compensate for a loss of resources at one point in ti me in exchange for resources at another point in time while keeping R(0) constant. In a continuous-time setting, MRS at age x can be calculated as log 0 vv xx v Rf lA x v lAfv , (19) log 0 z x z R Bzx fz Bfz , (20) Figure 1. Age-trajectory of Mortality. The maximum age- class to which the individual can possibly survive is set to nine, and the functional forms and parameter values are given as follows: ,qw x wx xe , Ax vxAvx , Bx zx Bzx , γ = 0.25, q = 0.5, B = 0.15, A(0) = 0.18, y(0) = 0, y(1) = 2, y(2) = 4, y(3) = 6, y(4) = 6, y(5) = 6, y(6) = 6, y(7) = 6, y(8) = 4, and y(9) = 4. Copyright © 2013 SciRes. TEL  J. KAGEYAMA 22 and log 0 wwx xx w Rw Rkl wl Rk (21) where , , and are respectively the marginal effects of reproductive, growth, and survival investments on R(0). Note that, with respect to survival investment, the change in resource surplus must be taken into account. 0 v R 0 z R 0 w R These equations show that MRS depends on various factors. Equation (19) illustrates that, in adulthood , MRS consists of three components; the mortality rate, the change in the reproductive efficiency, and the change in the marginal contribution of resources. This result is consistent with Sozou and Seymour [9], Chu et al. [10], and Kageyama [13]. In childhood, on the other hand, Equation (20) shows that MRS consists of two components; the change in the growth efficiency and the change in the marginal contri- bution of resources. As in Chu et al. [10], the mortality rate does not appear in MRS in childhood. Furthermore, Equation (21 ) shows that, throughout the life course, MRS can be calculated as the sum of three components; the change in the marginal contribution of resources, the change in the value of survival, and the mortality rate. Despite these variations, however, the values of MRS derived in Equations (19)-(21) are all equal to the mor- tality rate when resources are optimally allocated. In Equation (19), the second and the third terms cancel out on the optimal path as indicated in Equation (7), and MRS is reduced to the mortality rate. Similarly, as in Equation (8), the two terms in Equation (20) sum up to the mortality rate. Turning to Equation (21), it is also reduced to the mortality rate since, as indicated in Equa- tion (18), the first two terms cancel out. This result is intuitive in the reproductive period. The change in the reproductive efficiency is neutralized by the change in reproductive investment on the optimal path. However, giving up a unit of current reproductive investment for future investment still accompanies the mortality risk. Thus, to keep R(0) constant, the mortality risk needs to be compensated. By contrast, the result that MRS is equal to the morta- lity rate during the growth period might be counter-in- tuitive. As children do not yet reproduce, the timing of growth investment seems irrelevan t as long as they build up the same level of reproductive capacity at maturity. The reason for this result is that the marginal produc- tivity, , changes with age. As shown in Subsec- tion 2.2, the marginal productivity decreases as the chance of reaching maturity increases, and its changing rate is exactly equal to the mortality rate. This is in tuitive considering that the change in the marginal productivity is originated by the change in the survival probability to maturity. For example, if B is constant across ages, the marginal productivity decreases with age to satisfy z Bf z zx zx zfz ll ,d . To give up a unit of current growth investment for future investment, the reduction in the marginal productivity must be compensated, and this compensation rate is given by the mortality rate. For this reason, MRS in childhood is, as in adulthood, equal to the mortality rate. 3.2. Growth Process, Mortality, and MRS The result that MRS in childhood on the optimal path is equal to the mortality rate does not depend on the type of growth process. For example, consider as in Chu et al. [10] that the amount of resources, y(x), is determined by body size, which in turn depends on growth investment up to age x. I assume, for simplicity, that y(x) is equal to body size at age x. In this case, the growth process can be described as 0ˆˆˆ 0 x yxygsx x x (22) where s(x) is growth investment allocated for increasing body size and , sx x is the contribution of the investment at each point in time. Here, I assume that , sx x is sufficiently concave in s(x) to have an interior solution9. Incorporating this size effect into the lifetime budget con- straint in Equation (5), we obtain the first-order conditions: , s gsxxLx 1 (23) for all x where ˆ x Lxlx lxdx ˆ , i.e., the re- maining life expectancy at age x. This shows that the marginal return of investing in body size is equal to its cost. The return is measured by the amount of resources that the individual is expected to accrue in the future, and the cost is equal to one, representing a unit of resource necessary for the investment. Equation (23) further illustrates that the change in the marginal productivity, s s, depends on the change in the remaining life expectancy. While the remaining life expectancy is high, the marginal produ ctivity is low so as to take advantage of the higher expected return. There- fore, if the growth efficiency is independent of age, s s increases and s decreases with age10. This is con- sistent with Chu et al. [10]. The next step is to calculate MRS. By applying the same method as before, it is given by 9We can also assume that y(x) depends on knowledge learned in the ast and interpret , sx x as the increment of knowledge. See g [25] for how knowledge affects fitness. 10The results that the two types of growth investment, s and z, change in the opposite direction suggests that the age-trajectory of whole growth investment is indeterminate. Copyright © 2013 SciRes. TEL  J. KAGEYAMA 23 log 0 ssx x s Rgs Ll gsL l . (24) The first term represents the change in the marginal pro- ductivity, the second term expresses the change in the remaining life expectancy, and the third term is the mor- tality rate. As indicated in Equation (23), the first two terms cancel out on the optimal path. As a result, MRS in childhood is again given by the mortality rate. 3.3. Time Preference While the intratemporal allocation of resources is invol- untary depending mostly on physiology, the intertempo- ral allocation is voluntary. It primarily depends on be- havior since we can deliberately control the allocation by deciding how much to consume at present. This implies that, to further analyze the intertemporal allocation, we need to move on to the economic frame- work that allows us to study voluntary behavior. We need to remember that humans do not intend to maximize fitness but rather behav e in a utility-maximizing manner, and that those who had preferences that coincided with the fitness-maximizing behavior spread over and fixed in the population. To do this, I employ a standard economic model in which the objectiv e function is given by lifetime utility, 0,dUxucxx x , (25) where ,x x is the discount factor for future utility, is a strictly concave instantaneous utility function, and is consumption, which can be re- garded as the sum of growth, reproductive, and survival investments. Note that the lifetime budget constraint presented in Equation (5) is still relevant. uc cx Here, the instantaneous utility directly d epends on age. Given that preferences were shaped by natural selection, the instantaneous utility should reflect the effect of con- sumption on reproductive success at the corresponding age. To incorporate this aspect into the model, I assume that the marginal utility on the fitness-maximizing con- sumption path is constant across ag es. This type of utility induces the individual to consume more while its mar- ginal effect on fitness is high. In this setting, MRS in utility is given by , , cx x c ucxx x xucxx . (26) The individual applies this rate to evaluate future con- sumption and determines the intertemporal allocation. Thus, for the voluntary allocation to match the fitness- maximizing allocation, this rate must be consistent with MRS in fitness obtained in Subsection 3.1, and must be equal to the mortality rate when it is evaluated on the fitness maximizing consumption path. In particular, when the instantaneous utility depends on age as described above, , cx ucxx is equal to zero on the fitness- maximizing path, and, thus, x x , i.e., the pure rate of time preference, is equal to the mortality rate11. These results further imply that, considering that time preference is psychologically embedded, the endowed rate of time preference is equal to the mortality rate in our evolutionary past when we existed as hunter-gather- ers. As a result, presuming that the age-trajectory of mortality in our evolutionary past is similar to the one in modern hunter-gatherer populations, we can predict that the endowed rate of time preference is U-shaped in age, reaching its lowest level in early adulthood, as is the age- trajectory of the mortality rate in modern hunter-gatherer populations (e.g., Hill and Hurtado [26]; Hill et al. [27]). This is consistent with empirical findings. As dis- cussed in Section 1, the discount rate decreases in child- hood. In adulthood, on th e other hand, while still empiri- cally inconclusive, Trostel and Taylor [28] and Read and Read [29] found that the discount rate increases with senescence12. The present study focuses on the biological discount rate ignoring the effect of social factors such as education, and may overestimate the discount rate in adulthood in modern human populations. Nevertheless, the age-trajectory of time preference predicted in the present analysis is still in line with the empirical find- ings. 4. Concluding Remarks This paper aims to understand the mechanism that coor- dinates intertemporal choice, paying particular attention to the impatience of children. To do this, it examines human life-history strategies, incorporating the growth period to maturity. The results show that the endowed rate of time preference is equal to the mortality rate in th e entire life course and, thus, is U-sh ap ed in ag e, ind ica ting that it is higher for children than for young adults. At the behavioral level, this provides a biological ex- planation as to why parents and children often have con- flicts in the allocation of resources when they make col- lective decisions. Due to the difference in the discount rate, parents, who follow their own time preference, would allocate less to the present than their children would. Namely, the difference in time preference gener- 11If we correctly perceive the survival probability and discount future utilities accordingly, the pure rate of ti me preference w ould be equal to zero. However, there is no biological basis to suppose that we are able to correctly perceive the survival probability. Given that time dis- counting is not limited to humans, it makes more sense to consider that time preference is psyc h olo gica lly embedded. 12These studies, however, differ in the age at which the discount rate reaches the lowest level. While Trostel and Taylor [28] found that the discount rate is lowest among individuals in their twenties, Read and Read [29] argued that it is lowest in their forties. Thus, the age at which time preference reaches its lowest level is still an open question. Copyright © 2013 SciRes. TEL  J. KAGEYAMA 24 ates parent-offspring conflict over the intertemporal al- location of resources, affecting the decision-making process particularly within the household13. One limitation of this study, however, is that it treats the growth period as one state and compresses the re- productive capacity into one parameter. This is equiva- lent to assuming that a unit of growth investment is ex- changeable between any points in time withi n the growth period although such a transaction may incur additional costs. In reality, it is often not the case. A deficiency in a particular type of growth investment at a certain age, for example, may not be compensated by the same kind of investment at a later age, and may have a permanent ef- fect. In such cases, intertemporal choice would not be as simple as the present analysis suggests, and the impa- tience of children, in particular, would be underesti- mated. Nevertheless, as the first-order approximation, this study provides an explanation for the age-trajectory of time preference, connecting it to mortality in both child- hood and adulthood. The future is discounted because survival is uncertain (Yaari [31]), and survival is uncer- tain because the future is discounted (Kirkwood [32]; Kirkwood and Rose [33]). Both propositions hold in the entire life course because time discounting and survival uncertainty are two sides of the same coin, reflecting the change in the value of survival. There are other factors, including the variability of the environment, sexual re- production and genetic relatedness, that can possibly affect time preference, but mortality is the baseline for time preference through the entire course of life. This study also suggests that the traditional view that time discounting is somethin g unfavorable is not entirely accurate. Time discounting generally carries negative connotations and has been described as cognitive defi- ciency, impatience, shortsightedness, myopia, irrational- ity, and so forth. For example, Ramsey [34] regarded time-discounting behavior as “a practice which is ethi- cally indefensible and arises merely from the weakness of the imagination (p. 543)”. However, as discussed in this paper, being impatient is not a deficiency, but an optimal trait in the biological sense that was acquired in the course of evolution. Our surrounding environment has changed since our evolutionary past, and the en- dowed rate of time preference is no longer optimal in the economic sense. Nevertheless, being too patient is not necessarily favorable either as it would cause other prob- lems such as dynamic inefficiency in macroeconomics and the postponement of reproduction that, together with the decline in reproductive efficiency, results in below- replacement fertility. Given our human nature, an appro- priate level of impatience may in fact be beneficial to our well-being. 5. Acknowledgements I thank Akihiko Matsui and Takashi Shimizu for their helpful suggestions in developing ideas for this paper, and Kazuhiko Kato for his helpful comments on the ear- lier draft of this paper. Any remaining errors are my own. This study is supported by Grant-in-Aid for Challenging Exploratory Research from the Ministry of Education, Culture, Sports, Science and Technology in Japan (No. 23653055). REFERENCES [1] A. Dauphin, A. R. El Lahga, B. Fortin and G. Lacroix, “Are Children Decision-Makers within the Household?” Economic Journal, Vol. 121, No. 553, 2011, pp. 871-903. http://dx.doi.org/10.1111/j.1468-0297.2010.02404.x [2] G. S. Becker, “A Theory of Social Interactions,” Journal of Political Economy, Vol. 82, No. 6, 1974, pp. 1063- 1093. http://dx.doi.org/10.1086/260265 [3] T. C. Bergstrom, “A Fresh Look at the Rotten Kid Theo- rem—And Other Household Mysteries,” Journal of Po- litical Economy, Vol. 97, No. 5, 1989, pp. 1138-1159. http://dx.doi.org/10.1086/261646 [4] E. Bettinger and R. Slonim, “Patience among Children,” Journal of Public Economics, Vol. 91, No. 1-2, 2007, pp. 343-363. http://dx.doi.org/10.1016/j.jpubeco.2006.05.010 [5] L. Steinberg, S. Graham, L. O’Brien, J. Woolard, E. Cauffman and M. Banich, “Age Differences in Future Orientation and Delay Discounting,” Child Development, Vol. 80, No. 1, 2009, pp. 28-44. http://dx.doi.org/10.1111/j.1467-8624.2008.01244.x [6] U. Teuscher and S. H. Mitchell, “Relation between Time Perspective and Delay Discounting: A Literature Re- view,” Psychological Record, Vol. 61, No. 4, 2011, pp. 613-632. [7] I. Hansson and C. Stuart, “Malthusian Selection of Pref- erences,” American Economic Review, Vol. 80, No. 3, 1990, pp. 529-544. [8] A. R. Rogers, “Evolution of Time Preference by Natural Selection,” American Economic Review, Vol. 84, No. 3, 1994, pp. 460-481. [9] P. D. Sozou and R. M. Seymour, “Augmented Discount- ing: Interaction between Ageing and Time-Preference Behaviour,” Proceedings of the Royal Society B, Vol. 270, No. 1519, 2003, pp. 1047-1053. http://dx.doi.org/10.1098/rspb.2003.2344 13The parent-offspring conflict in the present study differs from the original parent-offspring conflict in Trivers [30]. While the ultimate cause of the original parent-offspring conflict is the difference in re- latedness, the ultimate cause in the present study is the difference in life-history phase. [10] C. Y. C. Chu, H. K. Chien and R. D. Lee, “The Evolu- tionary Theory of Time Preference and Intergenerational Transfers,” Journal of Economic Behavior and Organiza- tion, Vol. 76, No. 3, 2010, pp. 451-464. Copyright © 2013 SciRes. TEL  J. KAGEYAMA Copyright © 2013 SciRes. TEL 25 http://dx.doi.org/10.1016/j.jebo.2010.09.011 [11] A. J. Robson and L. Samuelson, “The Evolution of Time Preference with Aggregate Uncertainty,” American Eco- nomic Review, Vol. 99, No. 5, 2009, pp. 1925-1953. http://dx.doi.org/10.1257/aer.99.5.1925 [12] P. D. Sozou, “Individual and Social Discounting in a Vis- cous Population,” Proceedings of the Royal Society B, Vol. 276, 1669, 2009, pp. 2955-2962. http://dx.doi.org/10.1098/rspb.2009.0401 [13] J. Kageyama, “The Intertemporal Allocation of Consump- tion, Time Preference, and Life-History Strategies,” Jour- nal of Bioeconomics, Vol. 13, No. 2, 2011, pp. 79-95. http://dx.doi.org/10.1007/s10818-011-9104-2 [14] B. Chowdhry, “Possibility of Dying as a Unified Expla- nation of Why We Discount the Future, Get Weaker with Age, and Display Risk-Aversion,” Economic Inquiry, Vol. 49, No. 4, 2011, pp. 1098-1103. http://dx.doi.org/10.1111/j.1465-7295.2009.00271.x [15] A. K. Acharya and R. J. Balvers, “Time Preference and Life Cycle Consumption with Endogenous Survival,” Economic Inquiry, Vol. 42, No. 4, 2004, pp. 667-678. http://dx.doi.org/10.1093/ei/cbh088 [16] A. J. Robson, B. Szentes and E. Iantchev, “The Evolu- tionary Basis of Time Preference: Intergenerational Transfers and Sex,” American Economic Journal: Micro- economics, Vol. 4, No. 4, 2012, pp. 172-201. http://dx.doi.org/10.1257/mic.4.4.172 [17] G. S. Becker and C. B. Mulligan, “The Endogenous De- termination of Time Preference,” Quarterly Journal of Economics, Vol. 112, No. 3, 1997, pp. 729-758. http://dx.doi.org/10.1162/003355397555334 [18] S. C. Stearns, “The Evolution of Life Histories,” Oxford University Press, Oxford, 1992. [19] N. Perrin and R. M. Sibly, “Dynamic Models of Energy Allocation and Investment,” Annual Review of Ecology and Systematics, Vol. 24, 1993, pp. 379-410. http://dx.doi.org/10.1146/annurev.es.24.110193.002115 [20] H. M. Taylor, R. S. Gourley, C. E. Lawrence and R. S. Kaplan, “Natural Selection of Life History Attributes: An Analytical Approach,” Theoretical Population Biology, Vol. 5, No. 1, 1974, pp. 104-122. http://dx.doi.org/10.1016/0040-5809(74)90053-7 [21] P. Wiessner, “Leveling the Hunter: Constraints on the Status Quest in Foraging Societies,” In: P. Wiessner and W. Schiefenhövel, Eds., Food and the Status Quest: An Interdisciplinary Perspective, Berghahn Books, Oxford, 1996, pp. 171-191. [22] P. Wiessner, “Hunting, Healing, and hxaro Exchange: A Long-Term Perspective on! Kung (ju/'hoansi) Large- Game Hunting,” Evolution and Human Behavior, Vol. 23, No. 6, 2002, pp. 407-436. http://dx.doi.org/10.1016/S1090-5138(02)00096-X [23] M. Gurven, W. Allen-Arave, K. Hill and M. Hurtado, “‘It’s a Wonderful Life’: Signaling Generosity among the Ache of Paraguay,” Evolution and Human Behavior, Vol. 21, No. 4, 2000, pp. 263-282. http://dx.doi.org/10.1016/S1090-5138(00)00032-5 [24] H. E. Ryder and G. M. Heal, “Optimal Growth with In- tertemporally Dependent Preferences,” Review of Eco- nomic Studies, Vol. 40, No. 1, 1973, pp. 1-33. http://dx.doi.org/10.2307/2296736 [25] Y. K. Ng, “The Paradox of the Adventurous Young and the Cautious Old: Natural Selection vs. Rational Calcula- tion,” Journal of Theoretical Biology, Vol. 153, No. 3, 1991, pp. 339-352. http://dx.doi.org/10.1016/S0022-5193(05)80574-0 [26] K. Hill and A. M. Hurtado, “Ache Life History,” Aldine De Gruyter, New York, 1996. [27] K. Hill, A. M. Hurtado and R. S. Walker, “High Adult Mortality among Hiwi Hunter-Gatherers: Implications for Human Evolution,” Journal of Human Evolution, Vol. 52, No. 4, 2007, pp. 443-454. http://dx.doi.org/10.1016/j.jhevol.2006.11.003 [28] P. A. Trostel and G. A. Taylor, “A Theory of Time Pref- erence,” Economic Inquiry, Vol. 39, No. 3, 2001, pp. 379- 395. http://dx.doi.org/10.1093/ei/39.3.379 [29] D. Read and N. L. Read, “Time Discounting over the Lifespan,” Organizational Behavior and Human Decision Processes, Vol. 94, No. 1, 2004, pp. 22-32. http://dx.doi.org/10.1016/j.obhdp.2004.01.002 [30] R. L. Trivers, “Parent-Offspring Conflict,” American Zoologist, Vol. 14, No. 1, 1974, pp. 249-264. [31] M. E. Yaari, “Uncertain Lifetime, Life Insurance, and the Theory of the Consumer,” Review of Economic Studies, Vol. 32, No. 2, 1965, pp. 137-150. http://dx.doi.org/10.2307/2296058 [32] T. B. L. Kirkwood, “Evolution of Ageing,” Nature, Vol. 270, No. 5635, 1977, pp. 301-304. http://dx.doi.org/10.1038/270301a0 [33] T. B. L. Kirkwood and M. R. Rose, “Evolution of Senes- cence: Late Survival Sacrificed for Reproduction,” Phi- losophical Transactions of the Royal Society B, Vol. 332, No. 1262, 1991, pp. 15-24. http://dx.doi.org/10.1098/rstb.1991.0028 [34] F. P. Ramsey, “A Mathematical Theory of Saving,” Eco- nomic Journal, Vol. 38, No. 152, 1928, pp. 543-559. http://dx.doi.org/10.2307/2224098
|