Y. YAMADA, S. KOSAKA
throughout the entire forest, while the fragmentation of the light
areas continued. The network of dark areas at 40 years ap-
peared to be strengthened and expanded compared to the pre-
sent and after 20 years.
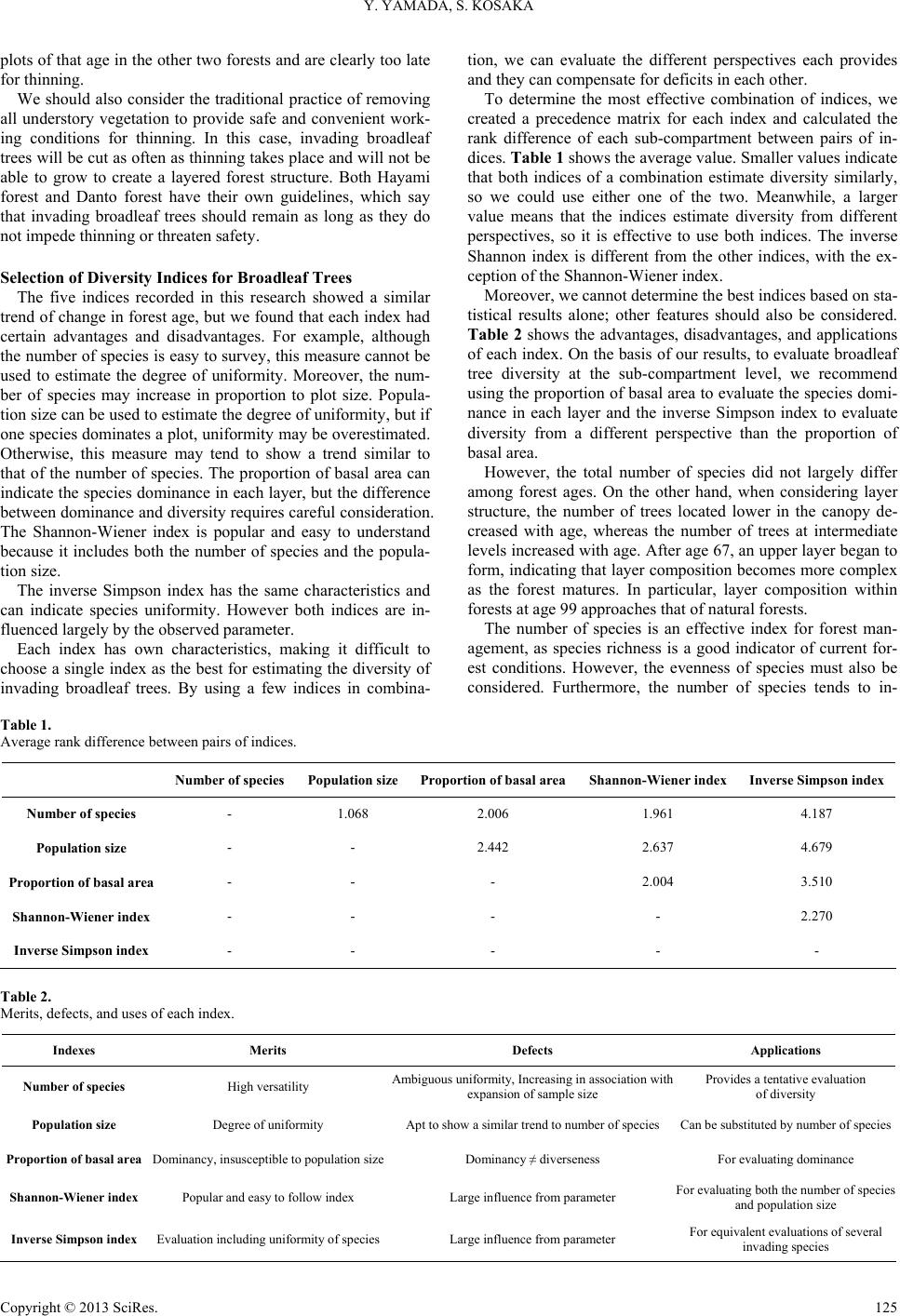
Figure 9 shows the change in proportion of the sub-com-
partments by stage. The proportion of mature stage sub-com-
partments decreased to 22% after 10 years under the 80-year
rotation scenario and then recovered gradually to 42% after 40
years. Meanwhile, the proportion of young stage sub-compart-
ments increased gradually to 35% after 30 years and then de-
creased. Middle stage sub-compartments occupied 44% of the
area at present, increased to 48% after 10 years, and thereafter
showed a downward trend. The overall percentages of high-
diversity stages (i.e., the sum of the young and mature stages)
were 56% at present, 51% after 10 years, 57% after 20 years,
72% after 30 years, and 65% after 40 years.
In this study, we created a simple scenario in which all sites
more than 80 years old were cut completely; thus, the first dec-
ade was on the overcutting side, and, after 80 years, the sites re-
turned to the starting point. Therefore, this scenario is strongly
affected by the present site age and location, and we cannot
change the arrangement of sites to improve the spatiotemporal
diversity of invading broadleaf trees. Clearly, if we set a dif-
ferent rotation scenario in Step 4, the results may change.
Conclusion
Our sub-compartment-scale analysis suggested that we can
classify each site into one of three stages according to the rich-
ness of diversity of the invading broadleaf trees: young stage,
less than 25 years with rich diversity but small height; middle
age, more than 26 years and less than 60 years with poor diver-
sity; and mature stage, more than 61 years with rich diversity
and a developed layer. For a Scots pine (Pinussylvestris) stand
in northern Scotland, Mason (2004) recognized four phases of
stand development: 1) stand initiation, from 0 to 20 years; 2)
stem exclusion, from 20 to 80 years; 3) understory re-initiation,
from 80 to 150 years; and (4) old growth, from 150 to 350 years.
Busing and Garman (2002) concluded that most wood quantity,
wood quality, and ecological objectives can be met with long
rotations of approximately 260 years. The ranges of our pro-
posed classification are shorter than those in previous studies.
Invading broadleaf trees may still be in the process of growing
even in our mature stage. In Japan, there are artificial forests
21%
29% 31% 35%
23%
44%
48% 43% 28%
35%
35%
22% 26%
37% 42%
0%
20%
40%
60%
80%
100%
presentafter 10after 20after 30after 40
Pr oportion of subcom partm ents
Elapsed year s
Mature stage
Middle stage
Young stage
Figure 9.
Change in the proportion of sub-compartments by stage.
older than 200 years, but the conventional long-rotation system
in Japan is more than 70 and less than 100 years, considering
economic efficiency and management conditions. Busing and
Garman (2002) noted that certain objectives could be met with
shorter rotations of 80 - 150 years, when treatments of thinning
and canopy tree retention were applied. Hartley (2002) argued
that during establishment, forest managers should consider in-
novations in snag and reserve tree management, in which ma-
ture native trees and/or understory vegetation were left unhar-
vested or allowed to regenerate. The same idea applies to our
study area: invading broadleaf trees should be retained as long
as they do not impede thinning or risk safety.
In this study, we examined five diversity indices for broad-
leaf trees and studied the features of each index. We then ex-
amined combinations of the diversity indices. The proportion of
basal area and inverse Simpson index were identified as the
index pair for evaluating the diversity of invading broadleaf
trees at a single site. In future, we plan to test additional diver-
sity indices to identify and confirm the most suitable indices for
evaluating broadleaf tree diversity in artificial forests.
From the results of our basin-scale analysis, we proposed a
model for evaluating the diversity richness of invading broad-
leaf trees in whole forests. The simulation showed a spatio-
temporal change in richness. Regarding the importance of a
basin-scale perspective, Kupfer et al. (2006) noted that the
study of forest fragmentation effects was shifting away from a
patch-based perspective focused on factors to a landscape-mo-
saic perspective that recognizes the importance of gradients in
habitat conditions. Fischer et al. (2006) suggested that a land-
scape should include structurally characteristic patches of na-
tive vegetation, corridors, and stepping stones between them, a
structurally complex matrix, and buffers around sensitive areas.
Although our GIS-based model can show a basin-scale per-
spective, we could not quantitatively evaluate the fragmentation
of sites as a structurally complex matrix and network of rich
diversity sites as corridors in this study. In future, we plan to
improve the rotation scenario of our proposed model by incor-
porating a road-construction plan and felling plan that specifies
when and which sites to fell and plant.
REFERENCES
Brockerhoff, E., Jactel, H., Parrotta, J., Quine, C., & Sayer J. (2008).
Plantation forests and biodiversity: Oxymoron or opportunity? Bio-
diversity Conservation, 17, 925-951.
http://dx.doi.org/10.1007/s10531-008-9380-x
Burslem, D. F. R. P. (2004). Plant diversity in forests. Encyclopedia of
Forest Sciences, 1, 40-44.
http://dx.doi.org/10.1016/B0-12-145160-7/00021-1
Busing, R. T., & Garman, S. L. (2002). Promoting old-growth charac-
teristics and long-term wood production in Douglas-fir forests. For-
est Ecology and Management, 16 0 , 161-175.
http://dx.doi.org/10.1016/S0378-1127(01)00443-1
Chubu Regional Forest Office (2011). Forest for people; National For-
est.
http://www.rinya.maff.go.jp/chubu/introduction/gaiyou_kyoku/pamp
hlet/pdf/20110223kyoku-panhu.pdf
Ehrlich, P. R. (1996). Conservation in temperate forests: What do we
need to know and do? Forest Ecology and Management, 85, 9-19.
http://dx.doi.org/10.1016/S0378-1127(96)03746-2
El-Keblawy, A. (2005). Artificial forests as conservation sites for the
native flora of the UAE. Forest Ecology and Management, 213, 288-
296. http://dx.doi.org/10.1016/j.foreco.2005.03.058
FAO (2012). Global forest resources assessment 2010.
http://www.fao.org/forestry/fra/fra2010/en/
Copyright © 2013 SciRes. 127