H. Narumi et al. / Advances in Bioscience and Biotechnology 4 (2013) 896-899
898
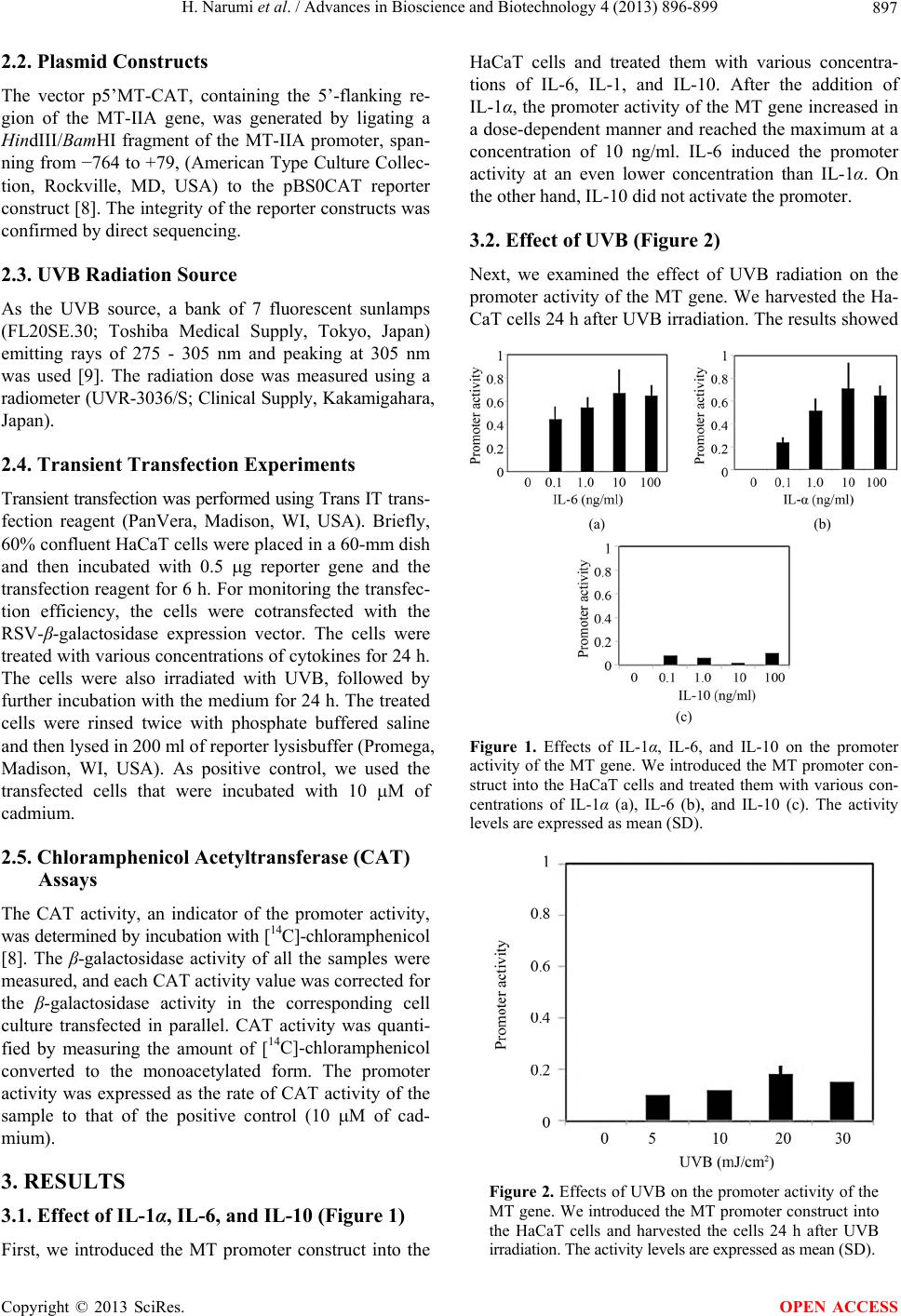
that UVB had no or little enhancing effect on the pro-
moter activity at doses ranging from 5 to 30 mJ/cm2.
Furthermore, we collected the treated cells several times
after irradiation, but we could not observe any strong
induction of the promoter activity.
4. DISCUSSION
IL-1α has many physiological functions in the immune,
metabolic, and hematopoietic systems. Keratinocytes are
a major source of IL-1α. As a proinflammatory cytokine,
IL-1α is involved in inflammatory and allergic skin dis-
eases such as psoriasis and contact dermatitis. It also
plays an important role in many recently defined autoin-
flammatory diseases [10]. IL-6 acts both as a pro- and
anti-inflammatory cytokine. It is also secreted by kerati-
nocytes, and it stimulates the immune response in both
normal and abnormal skin conditions [11]. Furthermore,
IL-1α and IL-6 are known to increase the MT gene ex-
pression and thereby, its protein expression. In this study,
we first examined the effects of IL-1α and IL-6 on the
MT promoter activity in epidermal keratinocytes. Our
results show that both cytokines induced promoter activ-
ity in a dose-dependent manner. We also investigated the
effect of IL-10 on MT expression, and as expected, we
did not find any significant effect of IL-10 on the pro-
moter activity.
The epidermis is the outermost layer of the skin. Kerati-
nocytes form a majority, i.e., about 95%, of the epider-
mis’ cells and are major targets of solar radiation. UVB
benefits humans by catalyzing the production of vitamin
D, but it also causes sunburn, photoaging, and skin can-
cers. UVB is responsible for the production of many cy-
tokines in the keratinocytes [12]. Furthermore, UVB is
known to induce MT expression in the epidermal kerati-
nocytes, and IL-6 is a possible mediator of MT induction
by UV radiation [13]. Therefore, we investigated whether
UVB radiation could induce MT promoter activity. Our
results showed, interestingly, that UVB radiation has no
or little effects on the promoter activity, although IL-6
clearly induced the promoter activity of the MT gene in
our study. This discrepancy implies non-transcriptional
mechanisms such as an increase in mRNA stability, sug-
gesting complex molecular regulation of the MT gene.
5. ACKNOWLEDGEMENTS
The authors would like to thank Ms Yuka Toyomaki, Mrs Yukiko
Tamura, Mrs Yuriko Takagi, and Ms Nanako Seitoh for their excellent
technical assistance. This work was supported in part by Grants-in-Aid
from the Ministry of Education, Science, Sports, and Culture of Japan.
REFERENCES
[1] Masters, B.A., Kelly, E.J., Quaife, C.J., Brinster, R.L. and
Palmiter, R.D. (1994) Targeted disruption of metallothio-
nein I and II genes increases sensitivity to cadmium.
Proceedings of the National Academy of Sciences of the
United States of America, 91, 584-588.
doi:10.1073/pnas.91.2.584
[2] Lazo, J.S., Kondo, Y., Dellapiazza, D., Michalska, A.E.,
Choo, K.H. and Pitt, B.R. (1995) Enhanced sensitivity to
oxidative stress in cultured embryonic cells from trans-
genic mice deficient in metallothionein I and II genes.
Journal of Biological Chemistry, 270, 5506-5510.
doi:10.1074/jbc.270.10.5506
[3] Hanada, K., Sawamura, D., Tamai, K., Baba, T., Hashi-
moto, I., Muramatsu, T., Miura, N. and Naganuma, A.
(1998) Novel function of metallothionein in photoprotec-
tion: Metallothionein-null mouse exhibits reduced toler-
ance against ultraviolet B injury in the skin. Journal of
Investigative Dermatology, 111, 582-585.
doi:10.1046/j.1523-1747.1998.00342.x
[4] Angel, P., Poting, A., Mallick, U., Rahmsdorf, H.J.,
Schorpp, M. and Herrlich, P. (1986) Induction of metal-
lothionein and other mRNA species by carcinogens and
tumor promoters in primary human skin fibroblasts. Mo-
lecular and Cellular Biology, 6, 1760-1766.
[5] Shiraishi, N., Yamamoto, H., Takeda, Y., Kondoh, S.,
Hayashi, H., Hashimoto, K. and Aono, K. (1986) In-
creased metallothionein content in rat liver and kidney
following X-irradiation. Toxicology and Applied Phar-
macology, 85, 128-134.
doi:10.1016/0041-008X(86)90106-7
[6] Baba, T., Nakano, H., Tamai, K., Sawamura, D., Hanada,
K., Hashimoto, I. and Arima, Y. (1998) Inhibitory effect
of beta-thujaplicin on ultraviolet B-induced apoptosis in
mouse keratinocytes. Journal of Investigative Dermatol-
ogy, 110, 24-28. doi:10.1046/j.1523-1747.1998.00078.x
[7] Sato, M., Sasaki, M. and Hojo, H. (1994) Differential
induction of metallothionein synthesis by interleukin-6
and tumor necrosis factor-alpha in rat tissues. Interna-
tional Journal of Immunopharmacology, 16, 187-195.
doi:10.1016/0192-0561(94)90075-2
[8] Nakano, H., Ikenaga, S., Aizu, T., Kaneko, T., Matsuzaki,
Y., Tsuchida, S., Hanada, K. and Arima, Y. (2006) Hu-
man metallothionein gene expression is upregulated by
beta-thujaplicin: Possible involvement of protein kinase
C and reactive oxygen species. Biological and Pharma-
ceutical Bulletin, 29, 55-59. doi:10.1248/bpb.29.55
[9] Hanada, K. (2000) Photoprotective role of metallothio-
nein in UV-injury-metallothionein-null mouse exhibits
reduced tolerance against ultraviolet-B. Journal of De-
rmatological Science, 23, S51-S56.
doi:10.1016/S0923-1811(99)00078-X
[10] Goldbach-Mansky, R. and Kastner, D.L. (2009) Autoin-
flammation: The prominent role of IL-1 in monogenic
autoinflammatory diseases and implications for common
illnesses. Journal of Allergy and Clinical Immunology,
124, 1141-1149. doi:10.1016/j.jaci.2009.11.016
[11] Sawamura, D., Meng, X., Ina, S., Sato, M., Tamai, K.,
Hanada, K. and Hashimoto, I. (1998) Induction of kerati-
nocyte proliferation and lymphocytic infiltration by in
vivo introduction of the IL-6 gene into keratinocytes and
Copyright © 2013 SciRes. OPEN ACCESS