Sexual Reproduction One Billion Years Ago 385
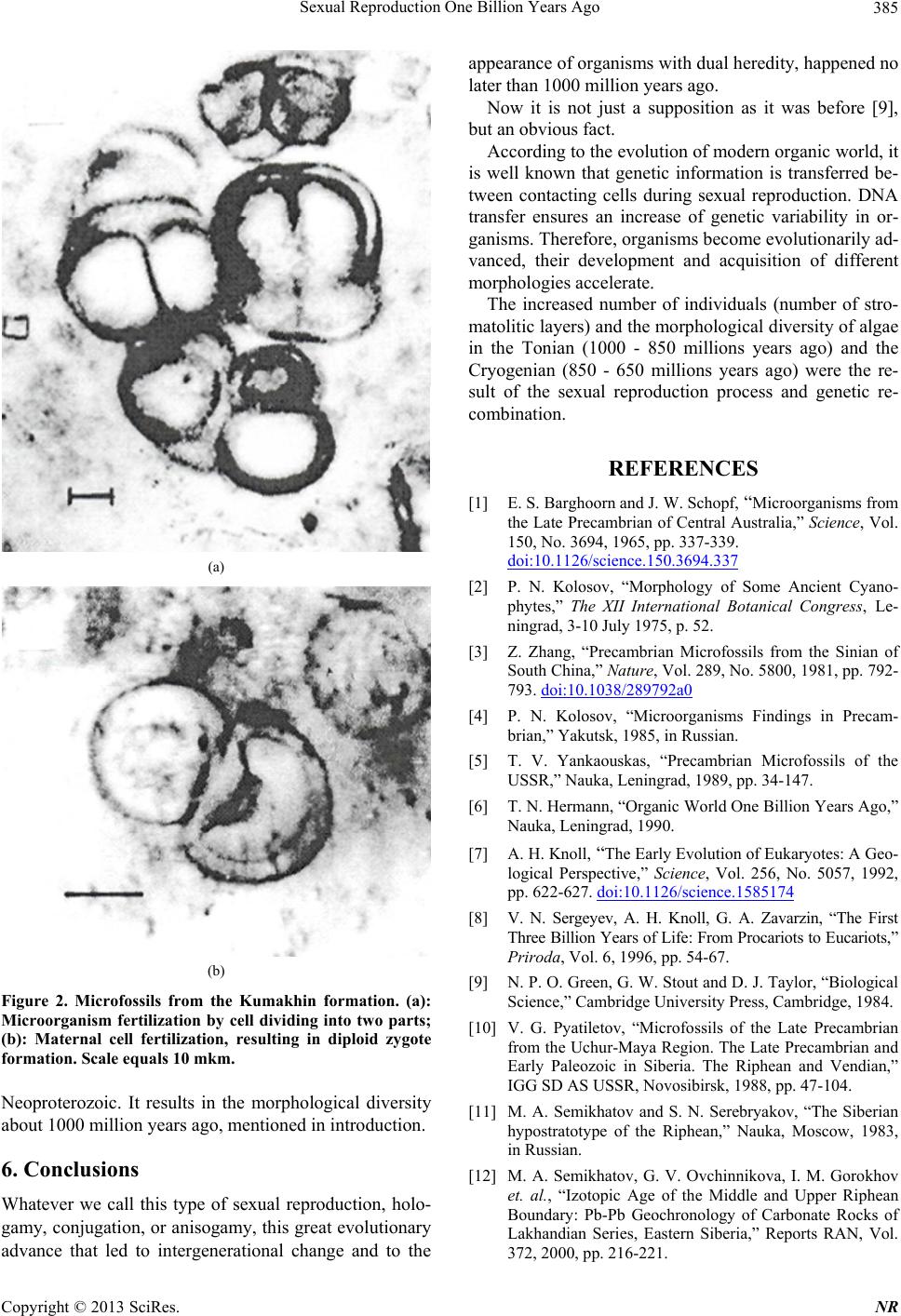
(a)
(b)
Figure 2. Microfossils from the Kumakhin formation. (a):
Microorganism fertilization by cell dividing into two parts;
(b): Maternal cell fertilization, resulting in diploid zygote
formation. Scale equals 10 mkm.
Neoproterozoic. It results in the morphological diversity
about 1000 million years ago, mentioned in introduction.
6. Conclusions
Whatever we call this type of sexual reproduction, holo-
gamy, conjugation, or anisogamy, this great evolutionary
advance that led to intergenerational change and to the
appearance of organisms with dual heredity, happened no
later than 1000 million years ago.
Now it is not just a supposition as it was before [9],
but an obvious fact.
According to the evolution of modern organic world, it
is well known that genetic information is transferred be-
tween contacting cells during sexual reproduction. DNA
transfer ensures an increase of genetic variability in or-
ganisms. Therefore, organisms become evolutionarily ad-
vanced, their development and acquisition of different
morphologies accelerate.
The increased number of individuals (number of stro-
matolitic layers) and the morphological diversity of algae
in the Tonian (1000 - 850 millions years ago) and the
Cryogenian (850 - 650 millions years ago) were the re-
sult of the sexual reproduction process and genetic re-
combination.
REFERENCES
[1] E. S. Barghoorn and J. W. Schopf, “Microorganisms from
the Late Precambrian of Central Australia,” Science, Vol.
150, No. 3694, 1965, pp. 337-339.
doi:10.1126/science.150.3694.337
[2] P. N. Kolosov, “Morphology of Some Ancient Cyano-
phytes,” The XII International Botanical Congress, Le-
ningrad, 3-10 July 1975, p. 52.
[3] Z. Zhang, “Precambrian Microfossils from the Sinian of
South China,” Nature, Vol. 289, No. 5800, 1981, pp. 792-
793. doi:10.1038/289792a0
[4] P. N. Kolosov, “Microorganisms Findings in Precam-
brian,” Yakutsk, 1985, in Russian.
[5] T. V. Yankaouskas, “Precambrian Microfossils of the
USSR,” Nauka, Leningrad, 1989, pp. 34-147.
[6] T. N. Hermann, “Organic World One Billion Years Ago,”
Nauka, Leningrad, 1990.
[7] A. H. Knoll, “The Early Evolution of Eukaryotes: A Geo-
logical Perspective,” Science, Vol. 256, No. 5057, 1992,
pp. 622-627. doi:10.1126/science.1585174
[8] V. N. Sergeyev, A. H. Knoll, G. A. Zavarzin, “The First
Three Billion Years of Life: From Procariots to Eucariots,”
Priroda, Vol. 6, 1996, pp. 54-67.
[9] N. P. O. Green, G. W. Stout and D. J. Taylor, “Biological
Science,” Cambridge University Press, Cambridge, 1984.
[10] V. G. Pyatiletov, “Microfossils of the Late Precambrian
from the Uchur-Maya Region. The Late Precambrian and
Early Paleozoic in Siberia. The Riphean and Vendian,”
IGG SD AS USSR, Novosibirsk, 1988, pp. 47-104.
[11] M. A. Semikhatov and S. N. Serebryakov, “The Siberian
hypostratotype of the Riphean,” Nauka, Moscow, 1983,
in Russian.
[12] M. A. Semikhatov, G. V. Ovchinnikova, I. M. Gorokhov
et. al., “Izotopic Age of the Middle and Upper Riphean
Boundary: Pb-Pb Geochronology of Carbonate Rocks of
Lakhandian Series, Eastern Siberia,” Reports RAN, Vol.
372, 2000, pp. 216-221.
Copyright © 2013 SciRes. NR


