 Open Journal of Genetics, 2013, 3, 224-233 OJGen http://dx.doi.org/10.4236/ojgen.2013.33025 Published Online September 2013 (http://www.scirp.org/journal/ojgen/) First steps on technological and genetic improvement of European abalone (Haliotis tuberculata) based on investigations in full-sib families Valérie Roussel1, Julien Charreyron1, Sylvain Labarr e 2, Alain Van Wormhoudt1, Sylvain Huchette2 1MNHN, UMR 7208 BOREA, Station de Biologie Marine, Concarneau, France 2France Haliotis, Plouguerneau, France Email: vroussel@mnhn.fr Received 24 July 2013; revised 20 August 2013; accepted 28 August 2013 Copyright © 2013 Valérie Roussel et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT The European abalone Haliotis tuberculata is of eco- nomical importance in Europe, and recently in France where most of the consumed abalones re- mained wild animals fished on the coast. Recently, the creation of hatcheries allowed the production of cul- tured animals, and aquaculture is in progress. To optimize selective breeding programs, different stud- ies were performed on adults and on their progenies. First, ten adult families, assumed to be bi-parental and produced in 2007 were analyzed. As these fami- lies were developed at the beginning of the hatchery production, the parentage of the individual necessi- tated to be tested. The progenies parentage assign- ment was done by using eight microsatellite DNA loci. In fine, the heritability was estimated from the analy- sis of variance of seven full-sib families which include from 23 to 27 progenies. Heritability estimate was the higher for length, width and weight (0.37, 0.29 and 0.40 respectively). A high correlation was also shown between these heritable traits which can be useful during animal breeding. Shell color traits were also analyzed, using image treatment procedure. Two traits were selected according to the global color of the shell: red color and hue. The distributio n of these traits evidenced a polygenic genetic control of shell color and measure of heritability evidenced a high to medium value for red color (Cr) and Hue (H) (0.33 and 0.20 respectively). No correlation was shown be- tween growth and this parameter. In a second time, juveniles families produced in 2010 were studied from parents issued of the first selection. A correlation be- tween size and density at juvenile stage was estab- lished but a statistical analysis demonstrated, by us- ing density as co-variable a significant effect of family on size. In these conditions, growth is highly heritable (0.74). This study is a first step toward the use of ge- netic ma rkers fo r select ion, but a lso a step toward the breeding improvement of European abalone. Keywords: European Abalone; Microsatellite; Heritability; Morphological Traits; Shell Color Traits; Juvenile 1. INTRODUCTION Abalones are marine gastropods living worldwide in temperate or tropical waters [1] and appreciated for its comestible foot. Because of their great economical im- portance, the first cultures of Asian abalone began in the 1950’s [2]. Researches on hatchery methods started in Japan in the 1960’s and later in California and Taiwan in the 1960’s, Korea, France and Ireland in the 1970’s, New Zealand and Australia in the 1980’s [3] and South Africa in the 1990’s [4]. Two ways can be considered for the improvement of abalone aquaculture, including techno- logical and genetic approaches [5,6]. The first one com- prises the hatchery conditions (temperature, food for larvae or adult, cages size and management) and the second would be based on Markers Assisted Selection (MAS). The technical improvement of hatchery condi- tions started in the 90’s for tropical abalone, with a first step related to fertilization and larval care, and a second step related to the survival and growth of post-settlement larvae [2]. These investigations are occuring for other genus of abalones [7,8], but no studies were devoted to European abalones. Concerning the second way of im- provement, the genetic way, the first studies started in the 2000’s for indo-pacific abalones [9,10], and the most advanced investigations are done on the Pacific abalone (Haliotis discus hannai) and the blacklip abalone (Haliotis OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 225 rubra) for which genetic linkage maps are already avail- able [11-14]. In Europe, abalone farming still remains new. Since the 1980’s, land based recirculation grow-out techniques were imported and adapted from South Africa or New Zealand to start an industry in Ire- land [15], but with little success so far. In the 2000’s, land based and sea based grow-out systems were developed in Western France yielding better results. A study concerning the optimization of MAS pro- grams evidenced the high interest of such techniques to improve genetic gain and cost benefits for Australian abalone [16]. But to date, no programs of MAS were developed on temperate abalones. The distribution of European Abalone Haliotis tuberculata spreads in a large temperate region, from the Morocco Atlantic coast to the English Channel and the Mediterranean sea [17,18]. The chromosome number of European abalone is 2n = 28, whereas the Indo-Pacific and North-Pacific species have a higher chromosome number with 2n = 32 or 36 [19]. These characteristics corroborate the important differ- ence between European and tropical abalones and strengthen the need of specific technical and genetic im- provement methods targeted to this species. In western France (Plouguerneau), hatchery techniques for H. tu- berculata were adapted from [20]. Nowadays, this spe- cies is at the first levels of domestication with three gen- erations of mass selection, with a choice of individuals based on morphological traits, and particularly growth rate. Genetic improvement can be efficient with mass selection in early phases of selection [21], but MAS would be more efficient in later stages, or for other traits, as it is now clearly established on several cattle or plant species. The aim of the present investigation was to study pa- rameters linked to the early stages of abalone develop- ment, and to estimate heritability of quantitative traits in this species, which are the first steps of a MAS pro- grams. 2. MATERIAL AND METHODS 2.1. Families Production and Growing Conditions 2.1.1. Adult Families Adult families were produced in “France Haliotis” hat- chery premises in Plouguerneau, and obtained from par- ents collected from wild stocks in North Western French Brittany (Plouguerneau and Roscoff). Each family was produced from a parental pair and kept in separated nursery tanks for the first year. Crossing experiments were done in individuals baskets to ensure the avoidance of contamination of sperm and egg, or mixing of larvae or juveniles among families. Then the juveniles of the 10 selected families of approximately 20 mm of maximum length were reared in small sea cages (Figure 1(a)) at about 10 m below chart datum for 2 years prior to sam- pling. Each family was reared in separate rearing units at the same density. The sea cages were lifted and the aba- lone fed every other week with fresh seaweed (mostly Palmaria palmata). 2.1.2. Juvenile Families Juvenile families (Table 1) were produced in 2010 in “France Haliotis” in Plouguerneau, and obtained from parents belonging to the company stock (Table 2). As the objective of the work was to have QTL analyses, the different crosses were done in order to optimize the ge- netic variability and the phenotypic variability in the progenies. In most cases, an individual belonging to a family representative of big individuals was crossed with an individual belonging to a family representative of small or medium individuals. Moreover, individuals were chosen according to the genetic differences evi- denced with the 8 SSR markers developed by [22] with the objective of crossing as genetically different indi- viduals as possible. Twelve full-sib families were thus obtained in May during the reproduction period, and were issued from the 10 full-sib families previously as Figure 1. Cages (a) and juvenile plates (b) used for rearing the families. The plates are 60 cm × 60 cm. Copyright © 2013 SciRes. OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 226 Table 1. Genitors selected for each of the 12 juvenile families, and size of their shells length. A2 to A8 corresponds to genitors presents in the company stock, and wild parents correspond to genitors coming from the Plouguerneau Coast in North Brit- tany. Juvenile Family Crossing Size (cm) J1 J2 J3 J4 J5 J6 J7 J8 J9 J10 J11 J12 Wild × A6 A6 × A8 A3 × A4 A5 × A3 A3 × A8 A3 × Wild A6 × A2 A1 × A2 A3 × A6 A3 × A2 A3 × A8 A6 × A2 6.2 × 6.7 5.9 × 6.3 6.8 × 5.9 5.8 × 6.7 5.8 × 6.7 6.4 × 6.7 6.4 × 6.6 6.1 × 6.2 6.3 × 6.1 5.6 × 6.6 5.8 × 6.0 6.8 × 6.6 previously described. Each of these juvenile families was disposed in a separate tank. All tanks comprised six bas- kets containing 20 plates each (Figure 1(b)), covered with Ulvella lens [20]. The juveniles were feed with Palmaria palmata, Ulva lactuca, and Saccharina sac- charina. 2.2. Sampling, DNA Extraction and Genotyping of Adult Families Thirty individuals were randomly chosen from the 10 adult families. An epipodial clip was taken from each individual and conserved in 70% ethanol for DNA ex- traction. DNA was extracted with the CTAB method [23], and was used to characterized COI mtDNA as previously described [24], and to amplify eight microsatellite loci in two quadriplex, as described in [22]. These loci are highly polymorphic and some of them present null al- leles. Amplified products were diluted in formamide containing GENESCAN-350(ROX) (Applied Biosystem) size standard, and size polymorphisms were screened using an ABI Prism 3130 DNA sequencer (Applied Bio- system). DNA fragments were analyzed using Gene- mapper software version 4.0 (Applied biosystem). 2.3. Trait Evaluation for Adult Families Direct measurements of length, width and weight were performed on three years old animals. Condition factor [25] was calculated using: 2.99 weight CF 5575 length mm g To evidence the color characteristics of the shells, and study the correlation between these characteristics and genetic parameters, one photograph of the entire outer shell of each abalone was taken using an Olympus Camedia C400 in a room of “France Haliotis” hatchery. Yelow color was chosen as background color after pre- Table 2. Number of individuals studied for each step of the work (N: number of individuals by sampled; n: number of indi- viduals genotyped; np: number of individuals used for pheno- typic traits, nc: number of individuals used for color traits). Adult Family N n np n c A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 30 30 30 30 30 30 30 30 30 30 0 27 25 23 0 26 25 24 27 0 0 27 25 23 0 26 25 24 27 0 0 26 22 20 0 19 24 19 17 0 liminary test. All the pictures were taken during daytime from the morning to the evening, inducing a difference of light intensity. This effect was eliminated by using appropriate color spaces components which did not take in account this phenomenon and allow to work on inde- pendent traits like: Cb (blue) and Cr (red) from YCbCR color space, and H (Hue) from HSV color space (the value of red and blue is included between 0 and 255, and hue value between 0 and 359). These pictures were treated by using classical methods of image processing (Figure 2). A binary image was obtained by using Otsu’s threshold method [26], and then, the background was separated from the area of interest to exclude the non-animal part of the picture. Next step of processing is cleaning of organisms at- tached to the shell (tags, algae, or small shellfishes), us- ing mathematical morphology tools, which could biased the result of color analysis processing. These processes produced a binary cleaned image of the abalone. In fine, this image was superposed to red, blue and hue picture permitting to exclude pixels of background and parasitic element of shell. The result was the selection and value of pixels of interest. The three averages of components were calculated. For the images programming, all the calculations were conduced with the software Octave (http://www.gnu.org/software/o ctave/). 2.4. Trait Evaluation for Juvenile Families During the last stage of biological development, the rela- tionship between growth and density on the plates was studied. For that, the density and size of individuals were determined at 90 days after fecundation for the 12 fami- lies. For each family, 120 individuals were evaluated on 6 different plates of 3600 cm², to estimate the variability of the density. For each plate, individual size was meas- ured on 20 individuals. 2.5. Data Analysis For parentage assignment, a chi-square test was perfor- Copyright © 2013 SciRes. OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 227 Figure 2. Image treatment of the abalone pictures: (a) Initial picture; (b) Threshold picture (with the background to 0); (c) Red levels of the picture; (d) Final picture. med to verify if the allelic frequencies observed in a family were in accordance with those expected in theory. Analysis of variance were carried out following the standard procedure of a fixed model with only the family effect for adult and with family and density effect for juveniles. Densities were ranked into 8 classes from 20 to 548 animals per plate. For adults: Yi = µ + Fi + ei For juveniles: Yij = µ + Fi + Dj + Fi·Dj + eij With µ = Grand mean, Fi = Family effect, Dj = Den- sity effect, Fi·Dj = Density within Family nested effect, and e = residual error. For both groups (adults and juveniles) and trait, full-sib heritability and its standard deviations was cal- culated after analysis of variance according to Falconer (1989) as described in [27]: 1212 2 AD FS ICE p VVV V hV with VA, VD, VI, VEC and VP: Additive, Dominant, Epistatic, Error and total phenotypic variance respectively. 2 FS V hV 2 22 2F FS w h and 12 211 ? 211 FS nt hnn N with: hFS: Full Sub heritability for each trait σ²F: variance between the progeny of different families σ²w: variance between the individuals within a family n: number of offspring per family ( for morpho- logical traits and 24 19 for color traits) N: number of families t: full-sib interclass correlation (12 S h) For juveniles, an analysis of covariance was per- formed to test the relationship between individual size and density on the plate. Correlation study and one way full-sib analysis of variance with family as the main ef- fect were performed using the R software package (http://cran.r-project.o rg/). 3. RESULTS 3.1. Adult Families 3.1.1. F am ily Assi gnment Two individuals from each family were randomly chosen and analyzed with COI and sperm lysin gene, evidencing that all were of H. t. tuberculata subspecies (data not shown). For each family, all progeny of 30 randomly chosen individuals was studied with SSR following three steps. The first one corresponds to the elimination of families which present more than four alleles with high frequency. As we want to work on bi-parental families, each parent can bring a maximum of two alleles, and as this species is diploid and as the SSR markers used were known to amplify a non duplicated portion of the ge- nome [22] the progeny must present a maximum of four different alleles. Three families (30%) with too many major alleles were eliminated. The second step corre- sponds to the elimination of individuals which brings alien alleles, never (or rarely) presents in other individu- als. This strategy has allowed the elimination of indi- viduals without parentage relationship with the others (10 to 16.6 % of the progeny). And the last step corre- sponds to the elimination of individuals which possess the same alleles than the major ones, but which present an association not compatible with the parental geno- types found (0% to 6% of the progeny). For the seven bi-parental families (excluding the pluri-parental fami- lies), the number of full-sib individuals ranged from 23 to 27 (76.6% to 90%) after the elimination of foreign individuals evidenced with the SSR markers. None of the families have common parent. 3.1.2. Heritabi l i ti es 1) Morphological Traits All three investigated morphological traits and the two Copyright © 2013 SciRes. OPEN ACCESS 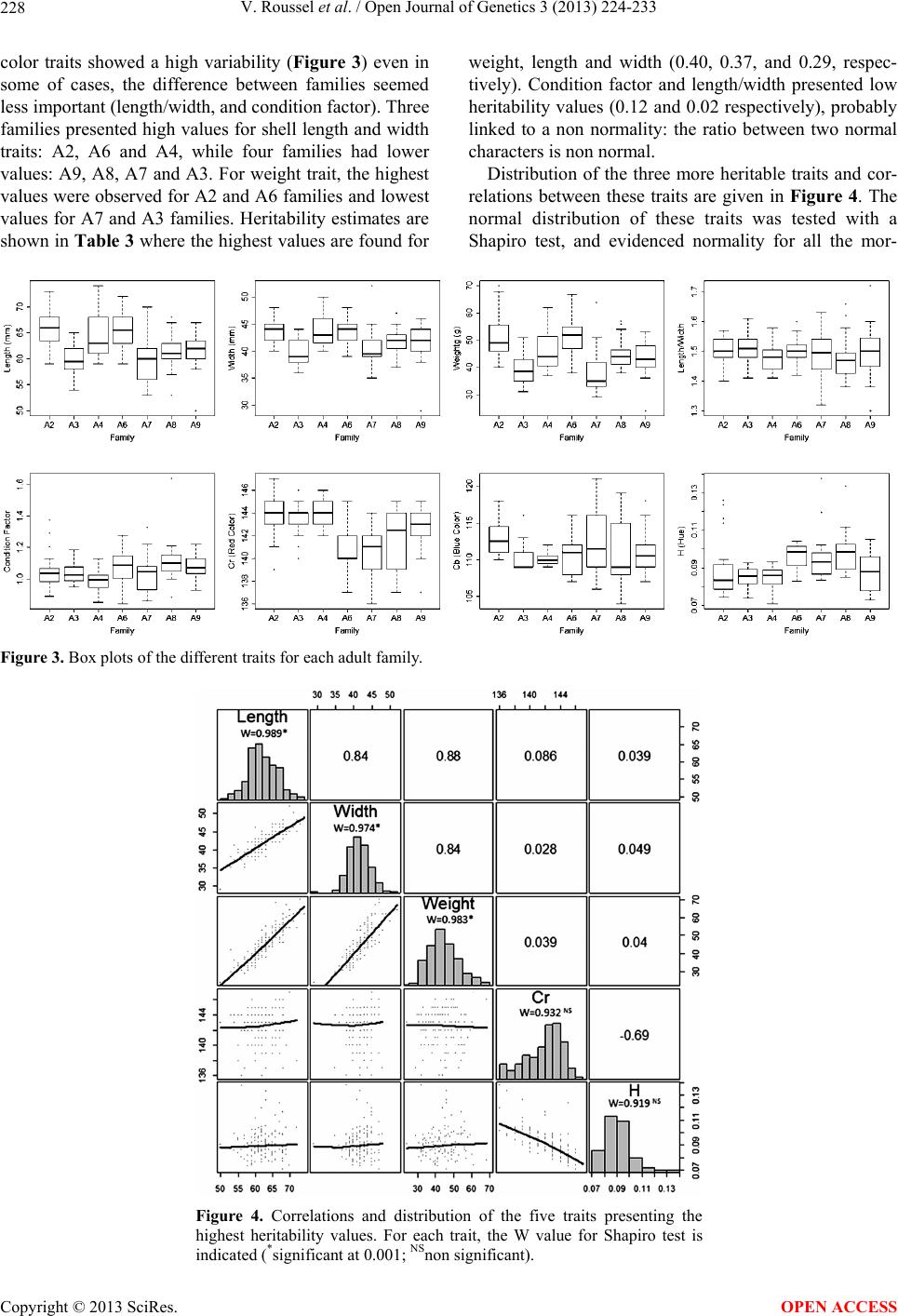 V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 Copyright © 2013 SciRes. 228 color traits showed a high variability (Figure 3) even in some of cases, the difference between families seemed less important (length/width, and condition factor). Three families presented high values for shell length and width traits: A2, A6 and A4, while four families had lower values: A9, A8, A7 and A3. For weight trait, the highest values were observed for A2 and A6 families and lowest values for A7 and A3 families. Heritability estimates are shown in Table 3 where the highest values are found for weight, length and width (0.40, 0.37, and 0.29, respec- tively). Condition factor and length/width presented low heritability values (0.12 and 0.02 respectively), probably linked to a non normality: the ratio between two normal characters is non normal. Distribution of the three more heritable traits and cor- relations between these traits are given in Figure 4. The normal distribution of these traits was tested with a Shapiro test, and evidenced normality for all the mor- Figure 3. Box plots of the different traits for each adult family. Figure 4. Correlations and distribution of the five traits presenting the highest heritability values. For each trait, the W value for Shapiro test is indicated (*significant at 0.001; NSnon significant). OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 229 phological traits. A high correlation (ranging from 0.84 to 0.88) between them was also evidenced, the most im- portant one was found between weight and length. 2) Color Traits As some visual observations in the natural environ- ment seem to indicate that the red abalones are taller and heavier than the others, we decided to investigate color parameters. For that, most of the individuals (Table 2) were also used to be described with image treatments. We do not observe different colors, but only one (red- brown) with different intensities. Two color traits (red color: Cr, and Hue: H) presented a high variability (Fig- ure 3). These traits presented a low heritability with h² value of 0.33 and 0.2 respectively (Table 3). The correlation between these two traits was equal to 0.69 (Figure 4). However, no correlation was shown between morpho- logical and color traits. The blue color (Cb) presented a low variability and heritability. 3.2. Juvenile Families The results on size and density on the plates is repre- sented on Figure 5 and evidenced a high variability be- tween families. Five of them presented an important size (J1, J3, J5, J7 and J9) and the other five presented a lower size (J4, J6, J8, J11 and J12). For density, the number of animals per 3600 cm2 presented a lower vari- ability between families and a higher level of in- tra-family variability. The correlation and covariance between these two parameters evidenced a negative link (r = −0.45, cov = −45.15) between size and density. Size heritability for juveniles was calculated and presented a high value of 0.74. 4. DISCUSSION 4.1. Adult Family Assignment Our results showed that the assignment of individuals after crossing in hatchery is necessary. Due to the high number of crossing it is not excluded that fertilization of eggs with sperm coming from different individuals may occur. It was the case in three progenies among the 10 selected. In this study, it was important to determine the genetic relationship between individuals of the progenies. During the nursery period, small larvae are cultivated on patches in the same tank. Even is the power of dispersal is low for the larvae, a mix between families is also pos- sible at this step. The adult growth conditions includes an opening of the cages and in most of the cases, some abalones can be taken out theses cages during the bringing of fresh sea- weeds. In such conditions, a mix between some adult individuals is also possible. The aim of the parental as- Table 3. Heritability (H²) and its standard deviation ( H²) for all traits studied. For Adult families: CF: Condition Factor, L/W: Length/Width, Cr: red color, Cb: blue color; H: Hue. For Juvenile families: SizeJuv: Size of the shell. Trait H² H² Length Width Weight CF L/W Cr Cb H Sizejuv 0.37 0.29 0.40 0.12 0.02 0.33 0.08 0.20 0.51 0.11 0.07 0.13 0.02 0.00 0.09 0.01 0.04 0.18 signment was to select only the real full-sib progeny of each family. But, as the genitors were not available, we had to determine their genotypes from those of their progeny. Whereas it was shown the presence of Haliotis tuber- culata coccinea COI signature in high proportion in French Brittany [24] the analysis of this marker indicated that all the analyzed individuals were of “tuberculata” sub-species. SSR analysis confirmed the absence of H. t. coccinea in French Brittany. This was important to high- light because nobody has tested the growth performances of the two sub-species and the putative role of mito- chondrial introgressed genomes in French Brittany. Microsatellite markers were evidenced to be efficient for parentage assignment in different species like com- mon carp [28] for which eight microsatellites were used to assign parentage in a full factorial cross of 10 dams and 24 sires, the shrimp Penaeus japonicus [29] for which eight microsatellites loci were used to discriminate 864 shrimp progeny originated from four ponds, abalone Haliotis asinina [30] for which five polymorphic loci were successfully used to assign parentage to young lar- vae, and for adult individuals [31]. In our case, as the parents genotypes were not known, the only solution to be sure of the common origin of the progenies was to choose only full-sib individuals. In fact, it the case of multi-parental progenies, the segregation of individuals would be impossible without the parental genotypes as start point. That is why the first step of the study was to select only bi-parental families where the number of alleles was reduced. The next steps corre- sponding to the elimination of foreign individuals with the study of allelic frequencies and genotype analysis permit to select only the effective progeny of the parents previously genotyped. This method is efficient to define parentage assignment when the genitors genotypes are not known but when the study of families without for- eign individuals is necessary, like in heritability studies. Copyright © 2013 SciRes. OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 230 Figure 5. Box plots of length and density for each juvenile family. Size was measured after 90 days on 120 individuals. Adult fami- lies from which they corresponded are indicated in Table 1. 4.2. Heritabilities All the studied traits present a moderate to high diversity. For instance, no markers assisted selection (MAS) pro- grams exist for European abalone. The first step of such programs, correspond to the identification of the traits which can be improved by selection, by studying their heritabilities. This work was done for several morpho- logical or color traits, which can be of interest in selec- tion. Our study was done on breeding conditions, by us- ing families selected in a farm. Higher the heritability is, and higher and easier the potential of improvement is. In general, we consider that traits which present an herita- bility value equal or superior to 0.2 can be useful for selection, and can be improved during a breeding pro- gram [32]. These results were obtained in a commercial environment, so they can be used directly for improve- ment programs. But according to this mating system, the calculation of heritability allow to obtain a broad-sense heritability, taking in account all the genetic parameters, contrarily to the narrow-sense heritability, taking in ac- count only the additive effects of the traits and leading to a finer estimate of the heritability for breeding purposes. Finer results could be obtained with another mating de- sign, like the “Full Factorial Design with Few Dams” which was evidenced to be superior to other ones [33] and could give access to the narrow-sense heritability. Heritabilities obtained for weight (0.40) appear to be comparables to those obtained for Haliotis asinina [31] which presents a weight heritability equal to 0.36. Shell length and width presents heritabilities of 0.37 and 0.29 respectively which is comparable to the results obtained with Haliotis discus hannai (0.36 and 0.32 respectively, [34]) or Haliotis rufescens for which an heritability of 0.34 was evidenced for shell length [35], but our result seems to be lower than the one obtained for Haliotis asinina shell length which was of 0.48 [31]. Concerning the shell morphology, it has been shown that heritability varies during animal development and becomes higher at the adult stage [35-37]. Consequently, for improving these traits, it would be better to use 24 months values. As the correlation between the three traits is very high, the weight can likely be improved by selecting for shell size. The ratio length/width and the Condition Factor pre- sents a low value of heritability, which can be linked to the high values of intra-family variability, and to the non normality of the characters. As the heritability calcula- tion takes into account the variability at the intra and inter-family level, when the first value is higher, the value of heritability is lowered (in this case, the in- tra-family variability is equal to 25% and 65% of the inter-family variability, for each of the two traits, respec- tively). Some previous studies concerning the shell color were done for Haliotis discus hannai [38,39] evidencing the monogenic genetic control of the shell color variation in this species presenting different colors (green, blue or orange). The Mendelian segregation of the progenies indicated that the color variant types were controlled by a single locus presenting a recessive and a dominant allele. As the shell color for Haliotis discus hannai is a mono- genic trait, the objective of the color study was to have information about genetic determinism of shell color in Haliotis tuberculata for witch some observations at the farm seemed to suspect the red shelled individuals to be taller than the others. Contrarily to what was suspected by observations, red ormers are not taller than the others. Consequently, this visual marker can not be used for an easy improvement of European abalones. As the shell color character presents a non normal distribution, it Copyright © 2013 SciRes. OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 231 seems to be highly influenced with environmental factors, so, for European abalone, the genetic control of the shell color seems to be different than the one previously ob- served for Haliotis discus hannai. It has been shown that the abalone diet could modify the shell color [39], that is why all the studied animals were feed with the same fresh algal diet, mostly Palmaria palmata, Ulva lactuca and Laminaria digitata, the ones which are usually con- sumed by the wild abalones during the study to avoid the expression of colors linked to the diet and thus, limit the environmental effect for the studied trait. Concerning Haliotis tuberculata, contrarily to other species, shell color present only different variations of red/brown, that is why we only studied the red color and the hue. Other colors were not representative, and inten- sity or saturation could not be representative because of absence of variability linked to the shell characteristics of our model. The high value of the correlation between the two traits indicates that the red color is the majority represented color in the shell panel. Contrarily to Haliotis discus hannai [38,39], no men- delian segregation could be evidenced in our progenies, but the observed distribution (Figure 4) evidenced a role of environmental conditions. If such a genetic control was not evidenced before in abalone species, it is not the case for another molluscan species like oyster [40,41]. These studies evidenced a polygenic control of the extern shell pigmentation like in Crassostrea gigas with the implication of one major gene in this trait. 4.3. Juvenile Im p ro ve ment For Rodriguez [42], settlement is a process comprising two phases: 1) a phase of searching for a suitable sub- stratum, and 2) a phase of attachment and metamorphosis. These parametres were excluded from our study because they were controled in the Hatchery. At the fixation stage, it was not possible to evidence a significant role of water temperature on fixation rate. In all cases (14 to 22˚C), this rate is always very variable and no temperature ap- pears to bring better results than others. At this stage, numerous environmental variables can be involved in fixation rate, like algal composition on the plates [43]. At the benthic stage, we observed a negative link be- tween the size of animals and the density, probably be- cause of the limited food on the plate. Moreover, due to the limited number of crossing adressed for this experi- ment, the asignment of the descendance was not neces- sary. The high heritability for the size (0.74) is probably due to 1) the elimination of the density parameter, and 2) the fact that for juvenile, we have less environmental parameters to take in account compared with adult ani- mals. It will be now important to study the correlation between juvenile size and adult size. If this parameter is high, it will be possible in the future, to select families at a juvenile stage without waiting the animals to be adults. 5. CONCLUSION Concerning shell color, contrarily to the Pacific abalone, European abalone does not present a monogenic control of this trait even if we could observe a major color (red). This trait is not related to growth parameters. This study brings other useful information about the European aba- lone selection, particularly for biological traits like weight and shell size, evidencing the high correlation between these traits and their high broad-sense heritabil- ity which indicates that a selection could be effective for such traits. This high heritability value was also evi- denced in juvenile families of 90 days and on adults. 6. ACKNOWLEDGEMENTS This work was supported by EC (SUDEVAB No. 222156 “Sustainable development of European SMEs engaged in abalone aquaculture”). The authors would like to thank Yves Barrière for its helpful and construc- tive comments to improve the manuscript. REFERENCES [1] Coleman, A.W. and Vacquier, V.D. (2002) Exploring the phylogenetic utility of ITS sequences for animals: A test case for abalone (Haliotis). Journal of Molecular Evolu- tion, 54, 246-257. doi:10.1007/s00239-001-0006-0 [2] Fleming, A.E. and Hone, P.W. (1996) Abalone aquacul- ture. Aquaculture, 140, 1-4. doi:10.1016/0044-8486(95)01183-8 [3] Hahn, K. (1989) Nutrition and growth of abalone. In: Handbook of Culture of Abalone and Other Marine Gas- tropods, CRC Press, Boca Raton, 135-180. [4] Britz, P.J., Hecht, T. and Mangold, S. (1997) Effect of temperature on growth, feed consumption and nutritional indices of Haliotis midae fed a formulated diet. Aquacul- ture, 152, 191-203. doi:10.1016/S0044-8486(97)00002-1 [5] Elliott, N.G. (2000) Genetic improvement programmes in abalone: what is the future? Aquaculture Research, 31, 51-59. doi:10.1046/j.1365-2109.2000.00386.x [6] Elliott, N.G., Evans, B., Conod, N., Bartlett, J., Officer, R. and Sweijd, N. (2000) Application of molecular genetics to the understanding of abalone population structure— Australian and South African case studies. Journal of Shellfish Research, 19, 510. [7] Grubert, M.A. and Ritar, A.J. (2005) The effect of tem- perature and conditioning interval on the spawning suc- cess of wild-caught blacklip (Haliotis rubra, Leach 1814) and greenlip (H. laevigata, Donovan 1808) abalone. Aqua- culture Research, 36, 654-665. doi:10.1111/j.1365-2109.2005.01269.x [8] Braid, B.A., Moore, J.D., Robbins, T.T., Hedrick, R.P., Tjeerdema, R.S. and Friedman, C.S. (2005) Health and survival of red abalone, Haliotis rufescens, under varying temperature, food supply, and exposure to the agent of Copyright © 2013 SciRes. OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 232 withering syndrome. Journal of Invertebrate Pathology, 89, 219-231. doi:10.1016/j.jip.2005.06.004 [9] Lymbery, A.J. (2000) Genetic improvement in the Aus- tralian aquaculture industry. Aquaculture Research, 31, 145-149. doi:10.1046/j.1365-2109.2000.00435.x [10] Roodt-Wilding, R. (2007) Abalone ranching: A review on genetic considerations. Aquaculture Research, 38, 1229- 1241. doi:10.1111/j.1365-2109.2007.01801.x [11] Liu, X., Liu, X., Guo, X., Gao, Q., Zhao, H. and Zhang, G. (2006) A preliminary genetic linkage map of the pa- cific abalone Haliotis discus hannai Ino. Marine Bio- technology, 8, 386-397. doi:10.1007/s10126-005-6133-7 [12] Qi, L., Yanhong, X., Ruihai, Y. and Akihiro, K. (2007) An AFLP genetic linkage map of pacific abalone (Ha- liotis discus hannai). Journal of Ocean University of China (English Edition), 6, 259-267. doi:10.1007/s11802-007-0259-3 [13] Sekino, M. and Hara, M. (2007) Linkage maps for the Pacific abalone (genus haliotis) based on microsatellite DNA markers. Genetics, 175, 945-958. doi:10.1534/genetics.106.065839 [14] Baranski, M., Loughnan, S., Austin, C.M. and Robinson, N. (2006) A microsatellite linkage map of the blacklip abalone, Haliotis rubra. Animal Genetics, 37, 563-570. doi:10.1111/j.1365-2052.2006.01531.x [15] Huchette, S. and Clavier, J. (2004) Status of the ormer (Haliotis tuberculata L.) industry in Europe. Journal of Shellfish Research, 23, 951-955. [16] Hayes, B., Baranski, M., Goddard, M.E. and Robinson, N. (2007) Optimisation of marker assisted selection for aba- lone breeding programs. Aquaculture, 265, 61-69. doi:10.1016/j.aquaculture.2007.02.016 [17] Wagner, R. and Abbott, R. (1967) Van Nostrand’s stan- dard catalog of shells. V.N. Company, Princeton. [18] Girard, A.(1972) La reproduction de l’ormeau Haliotis tuberculata L. Rev. Trav. Inst. Pêches Marit., 36, 163- 184. [19] Geiger, D.L. and Groves, L.T. (1999) Review of fossil abalone (Gastropoda, Vetigastropoda, Haliotidae) with comparison to recent species. Journal of Paleontology, 73, 872-885. [20] Daume, S., Huchette, S., Ryan, S. and Day, R.W. (2004) Nursery culture of Haliotis rubra: The effect of cultured algae and larval density on settlement and juvenile pro- duction. Aquaculture, 236, 221-239. doi:10.1016/j.aquaculture.2003.09.035 [21] Bentsen, H.B. and Olesen, I. (2002) Designing aquacul- ture mass selection programs to avoid high inbreeding rates. Aquaculture, 204, 349-359. doi:10.1016/S0044-8486(01)00846-8 [22] Roussel, V., Huchette, S. and Van Wormhoudt, A. (2010) Isolation and characterization of eight microsatellite loci in the European abalone, Haliotis tuberculata. Molecular Ecology Resources, 10, 751-754. [23] Doyle, J.J. and Doyle, J.L. (1987) A rapid isolation pro- cedure for small quantities of fresh leaf tissue. Phyto- chemistry Bulletin, 19, 11-15. [24] Van Wormhoudt, A., Le Bras, Y. and Huchette, S. (2009) Haliotis marmorata from Senegal; A sister species of Haliotis tuberculata: Morphological and molecular evi- dence. Biochemical Systematics and Ecology, 37, 747- 755. doi:10.1016/j.bse.2009.12.020 [25] Britz, P. J. (1996) The suitability of selected protein sources for inclusion in formulated diets for the South African abalone, Haliotis midae. Aquaculture, 140, 63-73. doi:10.1016/0044-8486(95)01197-8 [26] Otsu, N. (1979) A threshold selection method from gray- level histograms. IEEE Transactions on Systems, Man and Cybernetics, 9, 62-66. doi:10.1109/TSMC.1979.4310076 [27] Garenc, C., Silversides, F.G. and Guderley, H. (1998) Burst swimming and its enzymatic correlates in the threespine stickleback (Gasterosteus aculeatus): Full-sib heritabilities. Canadian Journal of Zoology, 76, 680-688. [28] Vandeputte, M., Kocour, M., Mauger, S., Dupont-Nivet, M., De Guerry, D., Rodina, M., Gela, D., Vallod, D., Chevassus, B. and Linhart, O. (2004) Heritability esti- mates for growth-related traits using microsatellite par- entage assignment in juvenile common carp (Cyprinus carpio L.). Aquaculture, 235, 223-236. doi:10.1016/j.aquaculture.2003.12.019 [29] Jerry, D.R., Preston, N.P., Crocos, P.J., Keys, S., Mead- ows, J.R.S. and Li, Y.(2006) Application of DNA par- entage analyses for determining relative growth rates of Penaeus japonicus families reared in commercial ponds. Aquaculture, 254, 171-181. doi:10.1016/j.aquaculture.2005.10.035 [30] Selvamani, M.J.P., Degnan, S.M. and Degnan, B.M. (2001) Microsatellite genotyping of individual abalone larvae: Parentage assignment in aquaculture. Marine Biotechnology, 3, 478-485. doi:10.1007/s1012601-0062-X [31] Lucas, T., Macbeth, M., Degnan, S.M., Knibb, W. and Degnan, B.M. (2006) Heritability estimates for growth in the tropical abalone Haliotis asinina using microsatellites to assign parentage. Aquaculture, 259, 146-152. doi:10.1016/j.aquaculture.2006.05.039 [32] Newkirk, G.F., Haley, L.E., Waugh, D.L. and Doyle, R. (1977) Genetics of larvae and spat growth rate in the oyster Crassostrea virginica. Marine Biology, 41, 49-52. doi:10.1007/BF00390580 [33] Dupont-Nivet, M., Vandeputte, M. and Chevassus, B. (2002) Optimization of factorial mating designs for in- ference on heritability in fish species. Aquaculture, 204, 361-370. doi:10.1016/S0044-8486(01)00839-0 [34] Deng, Y., Liu, X., Zhang, G. and Zhao, H. (2007) Genetic parameter estimates for growth traits at early stage of Pa- cific abalone, Haliotis discus hannai Ino. Acta Ocean- ologica Sinica, 26, 90-95. [35] Jonasson, S. and Stefansson, S.E. (1999) Genetic varia- tion for survival and shell length of cultured red abalone (Haliotis rufescens) in Iceland. Journal of Shellfish Re- search, 1, 621-625. [36] You, W.-W., Ke, C.-H., Luo, X. and Wang, D.-X. (2009) Divergent selection for shell length in two stocks of small Copyright © 2013 SciRes. OPEN ACCESS  V. Roussel et al. / Open Journal of Genetics 3 (2013) 224-233 Copyright © 2013 SciRes. 233 OPEN ACCESS abalone Haliotis diversicolor. Aquaculture Research, 41, 921-929. doi:10.1111/j.1365-2109.2009.02376.x [37] You, W., Ke, C., Luo, X. and Wang, D. (2010) Heritabil- ity of growth traits for small abalone Haliotis diversicolor estimated from sib matings. Journal of Shellfish Research, 29, 705-708. doi:10.2983/035.029.0322 [38] Kobayashi, T., Kawahara, I., Hasekura, O. and Kijima, A. (2007) Genetic control of bluish shell color variation in the Pacific abalone, Haliotis discus hannai. Journal of Shellfish Research, 23, 1153-1156. [39] Liu, X., Liu, X. and Zhang, G. (2007) Identification of quantitative trait loci for growth-related traits in the Pa- cific abalone Haliotis discus hannai Ino. Aquaculture Research, 38, 789-797. doi:10.1111/j.1365-2109.2006.01647.x [40] Evans, S., Camara, M.D. and Langdon, C.J. (2009) Heritability of shell pigmentation in the Pacific oyster, Crassostrea gigas. Aquaculture, 286, 211-216. doi:10.1016/j.aquaculture.2008.09.022 [41] Brake, J., Evans, F. and Langdon, C. (2004) Evidence for genetic control of pigmentation of shell and mantle edge in selected families of Pacific oysters, Crassostrea gigas. Aquaculture, 229, 89-98. doi:10.1016/S0044-8486(03)00325-9 [42] Rodriguez, S.R., Ojeda, F.P. and Inestrosa, N.C. (1993) Settlement of benthic marine invertebrates. Marine ecol- ogy progress series. Oldendorf, 97, 193-207. [43] Courtois de Viçose, G., Viera, M., Bilbao, A. and Iz- quierdo, M. (2010) Larval settlement of Haliotis tuber- culata coccinea in response to different inductive cues and the effect of larval density on settlement, early growth, and survival. Journal of Shellfish Research, 29, 587-591. doi:10.2983/035.029.0306
|