 Open Journal of Forestry 2013. Vol.3, No.3, 88-98 Published Online July 2013 in SciRes (http://www.scirp.org/journal/ojf) http://dx.doi.org/10.4236/ojf.2013.33015 Copyright © 2013 SciRes. 88 The Relative Importance of Natural Disturbances and Local Site Factors on Woody Vegetation Regeneration Diversity across a Large, Contiguous Forest Region Gerardo P. Reyes1,2*, Daniel Kneeshaw1, Louis de Grandpré1,3 1Department of Biological Sciences, Centre for Forest Research, University of Quebec in Montreal, Montreal, Canada 2Faculty of Interdisciplinary Studies, Lakehead University, Orillia, Canada 3Natural Resources Canada, Canadian Forest Service, Laurentian Forestry Centre, Ste-Foy, Canada Email: *gerardo1reyes@gmail.com, kneeshaw.daniel@uqam.ca, Louis.DeGrandpre@RNCan.gc.ca Received February 3rd, 2013; revised April 21st, 2013; accepted May 11th, 2013 Copyright © 2013 Gerardo P. Reyes et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Stand-level diversity after natural disturbance can potentially differ across a large, contiguous forest re- gion despite being dominated by the same canopy species throughout as differences in disturbance types and local site conditions can regulate species distribution. Our main objective was to examine the relative importance of natural disturbances (spruce budworm (Choristoneura fumiferana) outbreak, windthrow, and their interaction) and local site factors (climate, physiography, and stand structure and composition variables) on woody vegetation diversity among three, physiographically distinct locations across a large, contiguous forest region. Seventy-six Abies balsamea-Betula spp. stands affected by natural disturbance were compared and analysed using canonical ordination methods, diversity indices, and ANOVA. Dif- ferent combinations of factors were important for vegetation re-establishment at each location. Differ- ences in alpha (α), beta (β), gamma (γ), Shannon’s H’, and evenness (J) diversity indices were observed among locations across the study region. Our findings indicate that while certain processes are important for maintaining canopy dominance by Abies balsamea and Betula spp. throughout the region, different combinations of factors were important for creating variation in woody species diversity among locations that resulted in greater woody species diversity at the regional scale. Keywords: Regeneration; Natural Disturbance; Environmental Factors; Species Diversity; Eastern Boreal Mixedwood Region Introduction Climate determines the regional species pool (Dansereau, 1954; Neilson et al., 1992; Messaoud et al., 2007). However, woody species distribution and abundance across the landscape is largely determined by the disturbance regime (Gromtsev, 2002; Belote et al., 2009). Consequently, a distinct spatio-tem- poral arrangement of woody species is often observed at local scales due to variation in disturbance type, severity, and fre- quency across the landscape (Chen & Popadiouk, 2002; Reyes et al., 2010). Apart from natural disturbances, local site factors also play key roles in forest development, as they are capable of influencing the disturbance regime itself, and can directly affect species diversity after disturbance (Peterson, 2000; McGuire et al., 2002; Wang et al., 2006). Different types of disturbance influence regeneration patterns in distinctive ways depending on the severity of effects on for- est vegetation and stand structure, and the variation in the tem- poral release of newly available resources. Windthrow and spruce budworm (Choristoneura fumiferana) outbreak, for example, have been shown to result in alternative successional trajectories in boreal ecosystems due to differences in the de- gree of soil disturbance and in the timing of tree mortality (Krasny & Whitmore, 1992; Nagel & Diaci, 2006). In the case of windthrow, this disturbance type can substantially alter can- opy species dominance whereas budworm damage can simply delay or promote development towards late-succession condi- tions (Bergeron, 2000; Chen & Popadiouk, 2002). However, variation in forest regeneration among stands experiencing the same disturbance type can also occur because of differences in local site conditions (Turner et al., 1997; Reyes & Kneeshaw, 2008). For example, where advance regeneration is abundant, windthrow may cause only structural versus compositional changes in the tree canopy (Peterson, 2000; Reyes et al., 2010). Moreover, the sub-canopy tree and shrub community can dis- play variation in responses to natural disturbances depending on light availability and level of soil disturbance; and depending on the relative speed of development and growth after distur- bance, can ultimately impact the composition of the regenerat- ing tree canopy (Gray et al., 2005; Hart & Chen, 2006). Other local site factors such as pre-disturbance stand composition, physiography, and stand structural characteristics have been shown to affect woody vegetation diversity after natural distur- bance at the stand scale (Dupont et al., 1991; Osumi et al., 2003; *Corresponding author.  G. P. REYES ET AL. Rodriguez-Garcia et al., 2011). However, it is unclear if the same factors play key roles in determining re-vegetation pat- terns throughout an entire forest region. Factors can differ in importance among locations across a region, operate at local or regional scales, and can work synergistically to affect species diversity (Wimberly & Spies, 2001; Hillebrand & Blenckner, 2002; Huebner & Vankat, 2003). Examining woody vegetation distribution after natural disturbances at various locations across a large, contiguous forest region dominated by the same canopy species can thus help to resolve potential contrasts by determining the different factors responsible for any observed variation across the region, for differences in responses to the same factors, or conversely, potentially link fine and broad- scale vegetation recovery patterns to a few important factors that have consistent effects on woody species diversity across the region as a whole. The purpose of our study is to document the relative impor- tance of natural disturbances and local site factors on woody vegetation regeneration diversity among three, physiographi- cally distinct locations across a large, contiguous forest ecosys- tem dominated by the same canopy species throughout. We specifically addressed the following questions: is the canopy composition maintained after disturbance or are different suc- cessional trajectories occurring? Does the type of natural dis- turbance determine woody species diversity or do local site factors more strongly influence regeneration distribution? Al- ternatively, do natural disturbances work synergistically with local site factors in determining woody species diversity? Lastly, do different combinations of disturbances and local site factors influence woody species diversity for specific locations or can a few common factors explain species distribution across the region? We hypothesized that given the physiographical differences among locations across the region, that different combinations of factors would be responsible for woody spe- cies diversity after natural disturbance at the local level; and consequently, that if different factors influence regeneration dynamics at local scales, it is likely that species diversity will differ across the region. Thus, although woody species diversity should be the highest at the regional scale, diversity should also reflect the intra-regional differences in disturbance history and the unique local site characteristics of these forests. Study Area The Abies balsamea-Betula spp. boreal mixedwood zone spans east to west across southern Quebec, Canada from 46˚ to 50˚N and 64˚ to 80˚W (Figure 1). It encompasses 23.8 million ha and represents 18.6% of the forested land of the province (MRNQ, 2003). The physiography of the region is highly vari- able. The western portion is continental, and is relatively flat to rolling. Topography becomes increasingly hilly to montane towards the east, as the eastern portion of the region includes the northeastern limit of the Appalachian mountain chain that runs southwest into the United States. The eastern portion also borders the Gulf of St. Lawrence, and thus contrasts with the western portion of the region by having a maritime influence. Elevations of forested areas range from 80 to 400 m in the west, and from sea level to 900 m in the east. Surface deposits are mostly glacial till or of lacustrine origin (Robitaille & Saucier, 1998). The soil moisture regime is classified as xeric-mesic to mesic, while soil drainage ranges from imperfect to rapid. Numerous disturbances such as fire, insect pests, and wind- throw occur at varying frequency and severity in eastern boreal mixedwoods. The current return interval length for catastrophic Figure 1. Study locations within the boreal mixedwood region of Quebec, Can- ada. (A) continental—Abitibi-Temiskaming; (B) northern coastal—the North Shore; (C) southern coastal—the Gaspé Peninsula. fires across the region ranges from 170 to 645 years (Bergeron et al., 2006), suggesting that less severe, but more frequent disturbances now assert greater influence on post-disturbance boreal mixedwood dynamics than in the recent past. Thus, we focused our attention on three types of non-fire disturbances that are common to boreal mixedwood forests: spruce budworm (Choristoneura fumiferana) outbreak, windthrow, and their interaction (nb., where a stand affected by spruce budworm outbreak is then subjected to windthrow prior to canopy recov- ery). We sampled within three physiographically distinct, widely dispersed locations (between 200 and 500 km) across the boreal mixedwood region of Quebec, Canada (Figure 1). The western sites are located in the Abitibi and Temiskaming municipalities (77˚30' to 79˚10'W and 47˚30' to 48˚20'N). These sites have a flat to rolling topography and are influenced by continental climate conditions. The two eastern coastal locations both have hilly to rolling topography but differ mainly by latitude and elevation. The northern coastal sites are situated along the north shore of the St. Lawrence River, within the Haute Côte Nord and Manicouagan municipalities, and between 48˚30' to 50˚00' N and 68˚00' to 69˚50'W. Stands in this location are near the northern limit of the boreal mixedwood region. The southern coastal sites are situated within the southern part of the Gaspé Peninsula, along the Chaleur Bay area of the Atlantic Ocean (between 48˚10' to 48˚35'N and 65˚45' to 66˚15'W). Other cli- mate, physiographic, and forest structure and composition characteristics are compared among geographic locations in Table 1. Relative species composition of the forest canopy can be quite variable within this forest type. We limited sampling to stands having two of the more common relative species compo- sitions: stands were either conifer-dominated (≥75% density of conifers in canopy) or mixed coniferous-deciduous (50% - 74% density of conifers in canopy) prior to disturbance. In all cases, Abies balsamea, Betula papyrifera, and Betula alleghaniensis dominated the forest canopy prior to disturbance, and repre- sented at least 60% of the coniferous and deciduous compo- nents within each stand, respectively. Picea glauca, Picea mariana, and Acer rubrum were also abundant in some sites while Thuja occidentalis, Pinus resinosa, Pinus strobus, Pinus Copyright © 2013 SciRes. 89  G. P. REYES ET AL. Copyright © 2013 SciRes. 90 Table 1. Comparison of climate, physiographic, and forest structure and composition variables at each location using factorial ANOVA. Values in parentheses indicate ±1 Standard Error of the mean. Unlike letters along a row indicate values are significantly different among locations (p < 0.05). Location Variable Continental Northern coastal Southern coastal Annual rain (mm) 691.3 (5.2) a 691.0 (7.5) a 724.3 (2.0) b Annual snow (mm) 274.3 (6.2) a 303.4 (4.5) b 238.6 (0.6) c Annual precipitation (mm) 965.6 (11.4) ab 988.9 (10.3) b 963.1 (1.4) a Annual temperature (˚C) 2.1 (0.1) a 2.2 (0.1) a 3.6 (0.1) b Mean monthly temperature-summer (˚C) 16.3 (0.1) a 15.3 (0.1) a 16.6 (0.1) a Mean monthly temperature-winter (˚C) −10.4 (0.1) a −8.8 (0.1) b −7.3 (0.1) c Mean windspeed (km·h−1) 12.9 (0.1) a 15.6 (0.0) b 18.7 (0.1) c Maximum windspeed (km·h−1) 52.8 (0.7) a 75.6 (0.1) b 76.0 (0.2) b Maximum gust speed (km·h−1)† 98.4 105.2 no data Physiography Elevation (m) 311.2 (11.8) a 274.2 (25.4) a 421.4 (9.7) b Slope* (%) 1.0 (0.0) a 1.2 (0.2) a 2.1 (0.1) b Latitude (degrees) 47.3 (0.1) a 49.0 (0.1) b 48.5 (2.3) b Longitude (degrees) 78.4 (0.1) a 68.8 (0.1) b 66.1 (2.8) c Soil drainage† 3.5 (0.2) a 3.1 (0.2) a 2.3 (0.1) b Forest structure and composition Mean disturbance area (ha) 11.6 (1.6) ab 8.9 (1.7) a 13.2 (1.0) b Snag density (ha−1) 138.2 (17.8) a 294.1 (33.3) b 499.3 (29.4) c Course woody debris density (ha−1) 1176.4 (10.9) a 1729.1 (82.2) b 2159.9 (105.2) c Percentage of trees uprooted 28.0 (3.5) a 31.9 (3.4) a 37.2 (1.9) a Decay class average ‡ 11.2 (0.1) a 11.2 (0.1) a 10.4 (0.1) b Tree regeneration density (ha−1) 126 + 907.9 (1936.6) a 36825.6 (2593.6) b 15284.5 (1525.3) a Shrub regeneration density (ha−1) 11982.9 (2288.1) a 33,969.6 (3407.6) b 7 454.0 (732.1) a Total regeneration density (ha−1) 24890.7 (2043.0) a 70,895.2 (3279.9) b 22 738.6 (1463.2) a Coniferous legacy tree density (ha−1) 1.3 (.3) a 8.9 (1.2) b 0.1 (0.2) c Deciduous legacy tree density (ha−1) 0.1 (0.03) a 2.7 (0.6) b 0.2 (0.2) a Crown cover of coniferous legacy trees (%) 3.5 (0.8) a 3.5 (0.9) a 4.2 (1.0) a Crown cover of deciduous legacy trees (%) 1.6 (0.7) a 1.7 (0.6) a 6.8 (1.2) b Stand density prior to disturbance (ha−1) 926.6 (89.3) a 1474.0 (83.8) b 1718.7 (80.7) b Note: †Insufficient data to include in analyses. *1: 0˚ to 10˚, 2: >10˚ to 20˚, 3: >20˚ to 30˚, 4: >30˚ to 40˚, 5: >40˚; †2: good, 3: moderate; 4: imperfect; ‡See Imbeau & Des- rochers (2002). banksiana, Larix laricina, Acer saccharum, Fraxinus ameri- cana, Fraxinus nigra, Populus balsamifera, and Populus tremuloides were occasionally present. Sampling Methods Disturbance type, climate, and physiographical data were de- rived from digitised aerial photos, eco-forestry maps, or sourced from various provincial and federal government agen- cies (MNRQ, 2003; Environment Canada, 2004), with the ex- ception of slope and elevation, which were determined on site using a clinometer and GPS unit, respectively. A total of 76 sites affected by spruce budworm outbreak, windthrow, or their interaction were examined: 24, 22, and 30 sites for the conti- nental (Abitibi-Temiskaming), northern coastal (North Shore), and southern coastal (Gaspé Peninsula) locations, respectively.  G. P. REYES ET AL. Up to six 20 × 20 m quadrats were sampled within each dis- turbed site. We limited our sampling to mature stands (≥80 years old-verified within each site by counting annual rings of remnant trees or old stumps) having at least 0.2 ha of contigu- ous canopy mortality to reduce variation resulting from differ- ences in stand age, disturbance severity, and the spatial extent of disturbance. Further, all quadrats were at least 40 m from the nearest intact forest and 30 m from the nearest logging road to avoid edge effects. Density of tree and shrub regeneration was quantified within each quadrat for three size classes: (1) 1 to 2 m tall, (2) >2 m tall and <4 cm dbh (1.37 cm), and (3) between 4 and 8 cm dbh using a nested plot design. Different height classes were used to determine which individuals were more or less likely to be recruited into the canopy. Some species in the region such as Abies balsamea can produce 1000 s of regenerating seedlings per hectare, many of which remain suppressed even after can- opy disturbance (Reyes & Kneeshaw, 2008). Class 1 regenera- tion was sampled in a 2 × 10 m area, class 2 in a 5 × 10 m area, and class 3 within the entire quadrat. Density of snags (standing dead trees >10 cm dbh) and coarse woody debris (downed trees >10 cm dbh) were determined and classified using a modified decay classification scale developed by Imbeau & Desrochers (2002). Coarse woody debris was also categorised as uprooted or snapped when possible; n.b., past research has shown that certain boreal species establish better in exposed mineral soils versus other substrates (Kuuluvainen & Juntunen, 1998). Thus, if more coarse woody debris is uprooted versus snapped, then one could expect a greater proportion of these types of species as well. Crown cover (m2) of legacy trees (mature overstory trees that survived the natural disturbance) within each quadrat, and density of deciduous and coniferous legacy trees within a 35 m radius of the quadrat centre were also determined. This was done to provide some indication of the amount of shade available to the regeneration, which can thus potentially influ- ence what species regenerate in close proximity, and provide information on potential sources of seed rain, respectively. Analyses Various analyses were used to compare and contrast woody vegetation diversity after natural disturbance between the con- tinental, northern coastal, and southern coastal locations. Com- parisons of the climate, physiographic, and forest structure and composition characteristics that were quantified for each loca- tion were made using analysis of variance (ANOVA) (SPSS 10.0, 1999) followed by the Student-Newman-Keuls multiple range test when significant differences were observed (at p < 0.05) (Table 1). Direct Gradient Analyses We used a series of redundancy analyses (RDA) (van den Wollenberg, 1977) to examine the relationships between local site factors (i.e., the various disturbance types, climate, physic- ographic, and stand structure and composition variables) and woody vegetation species distribution for each of the three study locations. For each analysis, the forward selection option was implemented to both rank the importance of each site fac- tor variable and to exclude redundant and non-significant vari- ables from the model. The significance of the explanatory effect of a site factor variable was determined using a Monte Carlo permutation test (200 permutations, p < 0.05) prior to the addi- tion of the next best fitting variable. CANOCO 4.0 software (ter Braak & Smilauer, 1998) was used to run the analyses. Vari- ables were centred and standardized as the site factor variables were measured using different units. Species Diversity Estimates Five measures of species diversity were calculated at two spatial scales: for both the entire study region and for each of the three study locations (continental, northern coastal, southern coastal). Thus, comparisons between regional and local diver- sity levels, as well as among the three study locations could be made. Calculations were also made for the entire woody vege- tation community and for each of the woody vegetation layers separately; i.e., canopy tree regeneration versus sub-canopy tree & shrub regeneration. This was done as different disturbance types can affect the understory community differently (Veblen & Ashton, 1978; Hart & Chen, 2006). Thus, diversity of the canopy tree regeneration may be influenced by the sub-canopy tree and shrub community that survived the disturbance or by sub-canopy species that can quickly establish soon after (Gray et al., 2005). Regional level species diversity estimates were the following: alpha diversity (α) represented mean site level richness, beta diversity (β) represented differences in richness among sites across the study region; i.e., the differences in species composi- tion among spatial units, while gamma diversity (γ) represented the total richness across the study region (Whittaker, 1960; Novotny & Weiblen, 2005). Beta diversity (Whittaker, 1960) was computed as: βγα . (1) Shannon’s diversity index (Shannon & Weaver, 1949): 'lniip p (2) where pi = proportion of the total sample belonging to species i (in our case, the relative density of a species), and evenness (J): 'lnαJH (3) where J is an index of how relative abundances of species are distributed (Pielou, 1966), were determined for each regenera- tion layer for each site. Analysis of variance followed by the Student-Newman-Keuls multiple range tests were used to compare the various diversity estimates among study locations and vegetation layers (p < 0.05). Species richness is partly a function of spatial scale (Palmer & White 1994). We acknowledge that differences in richness could be an artifact of sampling effort among locations. The total disturbed area examined for the continental, northern coastal, and southern coastal sites were 405.2, 505.6, and 1238.3 ha, respectively. We sampled the three locations using 35, 57, and 95 quadrats, respectively, to make sampling effort more equitable among locations. However, bias in species richness may also occur due to the unequal number of quadrats. Sample rarefaction (Krebs, 1989) was used to compute a spe- cies accumulation curve as a function of the number of quadrats examined for each location. PAST version 1.94b (Hammer et al., 2001) was used to run analysis. Results show that differ- ences in species richness due to differences in the number of quadrats examined in each location was negligible (Figure 2). Thus, we felt that making comparisons of diversity estimates among the three locations using our sampling protocol was a valid undertaking. Copyright © 2013 SciRes. 91 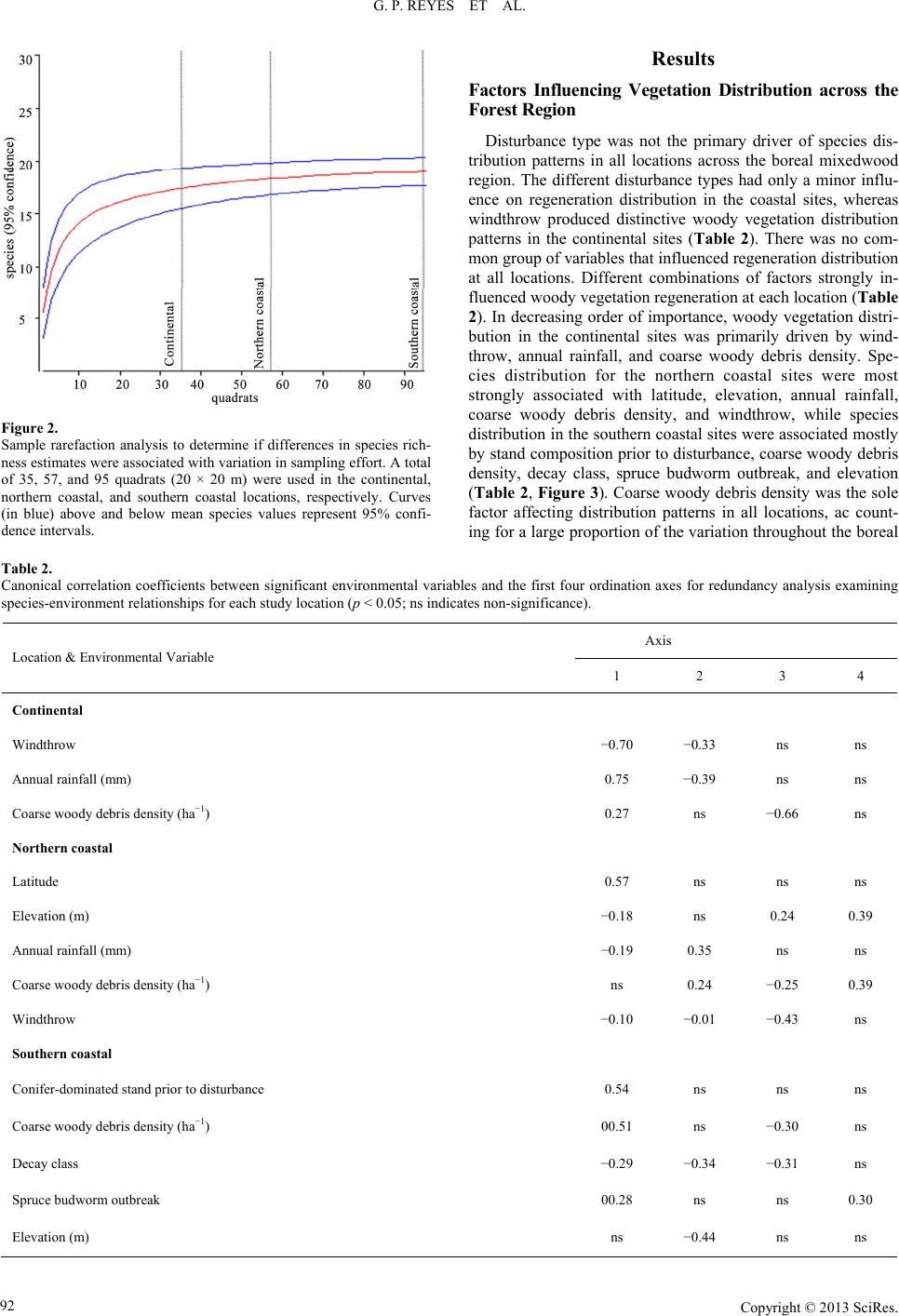 G. P. REYES ET AL. Copyright © 2013 SciRes. 92 Results Factors Influencing Vegetation Distribution across the Forest Region Disturbance type was not the primary driver of species dis- tribution patterns in all locations across the boreal mixedwood region. The different disturbance types had only a minor influ- ence on regeneration distribution in the coastal sites, whereas windthrow produced distinctive woody vegetation distribution patterns in the continental sites (Table 2). There was no com- mon group of variables that influenced regeneration distribution at all locations. Different combinations of factors strongly in- fluenced woody vegetation regeneration at each location (Table 2). In decreasing order of importance, woody vegetation distri- bution in the continental sites was primarily driven by wind- throw, annual rainfall, and coarse woody debris density. Spe- cies distribution for the northern coastal sites were most strongly associated with latitude, elevation, annual rainfall, coarse woody debris density, and windthrow, while species distribution in the southern coastal sites were associated mostly by stand composition prior to disturbance, coarse woody debris density, decay class, spruce budworm outbreak, and elevation (Table 2, Figure 3). Coarse woody debris density was the sole factor affecting distribution patterns in all locations, ac count- ing for a large proportion of the variation throughout the boreal Figure 2. Sample rarefaction analysis to determine if differences in species rich- ness estimates were associated with variation in sampling effort. A total of 35, 57, and 95 quadrats (20 × 20 m) were used in the continental, northern coastal, and southern coastal locations, respectively. Curves (in blue) above and below mean species values represent 95% confi- dence intervals. Table 2. Canonical correlation coefficients between significant environmental variables and the first four ordination axes for redundancy analysis examining species-environment relationships for each study location (p < 0.05; ns indicates non-significance). Axis Location & Environmental Variable 1 2 3 4 Continental Windthrow −0.70 −0.33 ns ns Annual rainfall (mm) 0.75 −0.39 ns ns Coarse woody debris density (ha−1) 0.27 ns −0.66 ns Northern coastal Latitude 0.57 ns ns ns Elevation (m) −0.18 ns 0.24 0.39 Annual rainfall (mm) −0.19 0.35 ns ns Coarse woody debris density (ha−1) ns 0.24 −0.25 0.39 Windthrow −0.10 −0.01 −0.43 ns Southern coastal Conifer-dominated stand prior to disturbance 0.54 ns ns ns Coarse woody debris density (ha−1) 00.51 ns −0.30 ns Decay class −0.29 −0.34 −0.31 ns Spruce budworm outbreak 00.28 ns ns 0.30 Elevation (m) ns −0.44 ns ns 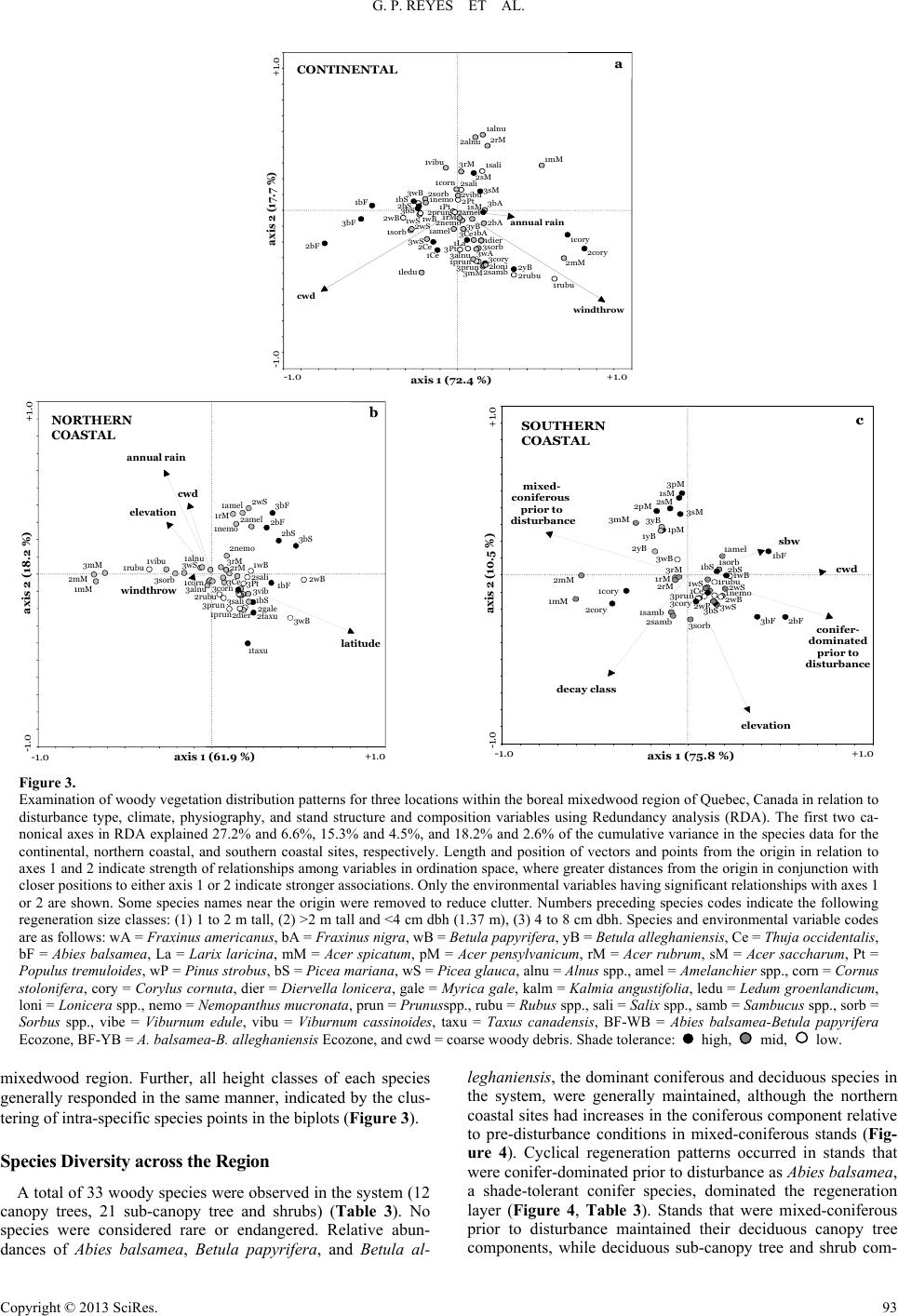 G. P. REYES ET AL. CONTINENTAL a axis 2 (17.7 %) axis 1 (72.4 %) -1.0 +1.0 -1.0 +1.0 windthrow cwd annual rain 2bF 2cory 1cory 2mM 1rubu 3bF 1bF 1mM 2yB 2rubu 2wB 1sorb 2wS 1bS 3bS 3wB 2bS 1wS 1wB 1ledu 2sorb 1nemo 3wS 2rM 3bA 2samb 2loni 2bA 1sM 1alnu 1sali 2Ce 3sorb 3wA 1bA 3sM 3mM 1La 1Ce 3cory 1pru n 2alnu 2sM 1dier 3prun 2amel 1vibu 3alnu 3Ce 3yB 2nemo 1Pt 1amel 2sali 3rM 2prun 3Pt 1rM 1corn 2Pt 2vibu -1.0 +1.0 -1.0 +1.0 latitude annual rain elevatio n cwd windthrow 2mM 1mM 3mM 2wB 3bS 3wB 2bS 1rubu 3bF 1bF 2bF 1vibu 2taxu 2gale 2amel 1wB 2wS 1bS 1taxu 3sorb 2sali 3Pt 1amel 3vib 3sali 2dier 3Ce 2rM 1nemo 1rM 1prun 3rM 2nemo 3corn 3prun 3wS 1alnu 2rubu 3alnu 1corn NORTHERN COASTAL axis 2 (18.2 %) axis 1 (61.9 %) b -1.0 +1.0 -1.0 +1.0 cwd sbw eleva tio n 1mM 2mM 2bF 1bF 2cory 3bF 1cory 3mM 1wB 2bS 2wS 1nemo 1amel 2wB 2yB 1sorb 2pM 3wS 1bS 1rubu 2wP 3bS 1pM 3yB 1yB 1Ce 1wS 2sM 1samb 2samb 3rM 2rM 3prun 3wB 1sM 3cory 1rM 3sM 3pM 3sorb SOUTHERN COASTAL axis 2 (10.5 %) axis 1 (75.8 %) c mixed- coniferous prior to disturba nce conifer- dominated prior to dist ur ba nc e decay class Figure 3. Examination of woody vegetation distribution patterns for three locations within the boreal mixedwood region of Quebec, Canada in relation to disturbance type, climate, physiography, and stand structure and composition variables using Redundancy analysis (RDA). The first two ca- nonical axes in RDA explained 27.2% and 6.6%, 15.3% and 4.5%, and 18.2% and 2.6% of the cumulative variance in the species data for the continental, northern coastal, and southern coastal sites, respectively. Length and position of vectors and points from the origin in relation to axes 1 and 2 indicate strength of relationships among variables in ordination space, where greater distances from the origin in conjunction with closer positions to either axis 1 or 2 indicate stronger associations. Only the environmental variables having significant relationships with axes 1 or 2 are shown. Some species names near the origin were removed to reduce clutter. Numbers preceding species codes indicate the following regeneration size classes: (1) 1 to 2 m tall, (2) >2 m tall and <4 cm dbh (1.37 m), (3) 4 to 8 cm dbh. Species and environmental variable codes are as follows: wA = Fraxinus americanus, bA = Fraxinus nigra, wB = Betula papyrifera, yB = Betula alleghaniensis, Ce = Thuja occidentalis, bF = Abies balsamea, La = Larix laricina, mM = Acer spicatum, pM = Acer pensylvanicum, rM = Acer rubrum, sM = Acer saccharum, Pt = Populus tremuloides, wP = Pinus strobus, bS = Picea mariana, wS = Picea glauca, alnu = Alnus spp., amel = Amelanchier spp., corn = Cornus stolonifera, cory = Corylus cornuta, dier = Diervella lonicera, gale = Myrica gale, kalm = Kalmia angustifolia, ledu = Ledum groenlandicum, loni = Lonicera spp., nemo = Nemopanthus mucronata, prun = Prunusspp., rubu = Rubus spp., sali = Salix spp., samb = Sambucus spp., sorb = Sorbus spp., vibe = Viburnum edule, vibu = Viburnum cassinoides, taxu = Taxus canadensis, BF-WB = Abies balsamea-Betula papyrifera Ecozone, BF-YB = A. balsamea-B. alleghaniensis Ecozone, and cwd = coarse woody debris. Shade tolerance: high, mid, low. mixedwood region. Further, all height classes of each species generally responded in the same manner, indicated by the clus- tering of intra-specific species points in the biplots (Figure 3). Species Diversity across the Region A total of 33 woody species were observed in the system (12 canopy trees, 21 sub-canopy tree and shrubs) (Table 3). No species were considered rare or endangered. Relative abun- dances of Abies balsamea, Betula papyrifera, and Betula al- leghaniensis, the dominant coniferous and deciduous species in the system, were generally maintained, although the northern coastal sites had increases in the coniferous component relative to pre-disturbance conditions in mixed-coniferous stands (Fig- ure 4). Cyclical regeneration patterns occurred in stands that were conifer-dominated prior to disturbance as Abies balsamea, a shade-tolerant conifer species, dominated the regeneration layer (Figure 4, Table 3). Stands that were mixed-coniferous prior to disturbance maintained their deciduous canopy tree components, while deciduous su-canopy tree and shrub com- b Copyright © 2013 SciRes. 93 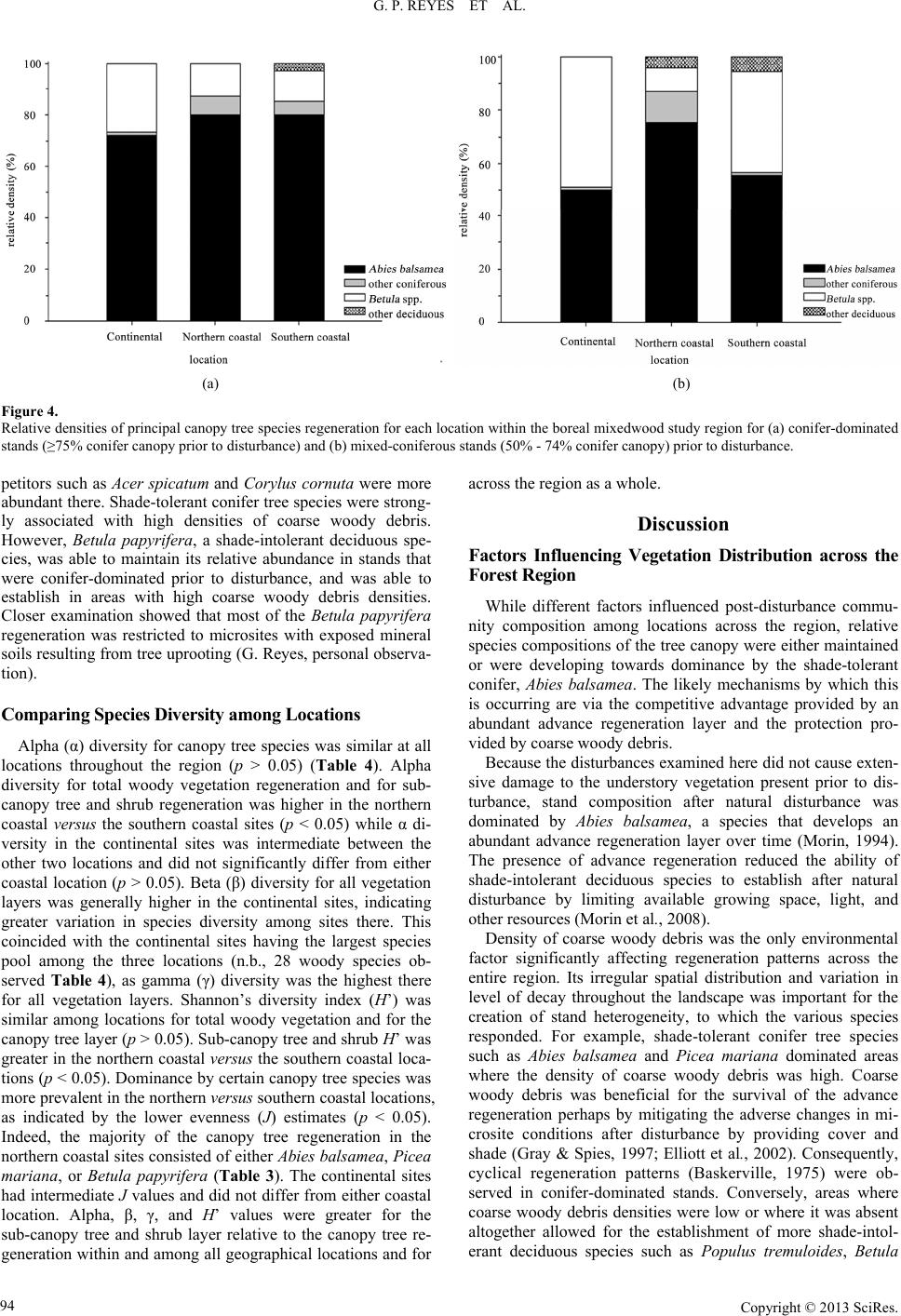 G. P. REYES ET AL. (a) (b) Figure 4. Relative densities of principal canopy tree species regeneration for each location within the boreal mixedwood study region for (a) conifer-dominated stands (≥75% conifer canopy prior to disturbance) and (b) mixed-coniferous stands (50% - 74% conifer canopy) prior to disturbance. petitors such as Acer spicatum and Corylus cornuta were more abundant there. Shade-tolerant conifer tree species were strong- ly associated with high densities of coarse woody debris. However, Betula papyrifera, a shade-intolerant deciduous spe- cies, was able to maintain its relative abundance in stands that were conifer-dominated prior to disturbance, and was able to establish in areas with high coarse woody debris densities. Closer examination showed that most of the Betula papyrifera regeneration was restricted to microsites with exposed mineral soils resulting from tree uprooting (G. Reyes, personal observa- tion). Comparing Species Diversity among Locations Alpha (α) diversity for canopy tree species was similar at all locations throughout the region (p > 0.05) (Table 4). Alpha diversity for total woody vegetation regeneration and for sub- canopy tree and shrub regeneration was higher in the northern coastal versus the southern coastal sites (p < 0.05) while α di- versity in the continental sites was intermediate between the other two locations and did not significantly differ from either coastal location (p > 0.05). Beta (β) diversity for all vegetation layers was generally higher in the continental sites, indicating greater variation in species diversity among sites there. This coincided with the continental sites having the largest species pool among the three locations (n.b., 28 woody species ob- served Table 4), as gamma (γ) diversity was the highest there for all vegetation layers. Shannon’s diversity index (H’) was similar among locations for total woody vegetation and for the canopy tree layer (p > 0.05). Sub-canopy tree and shrub H’ was greater in the northern coastal versus the southern coastal loca- tions (p < 0.05). Dominance by certain canopy tree species was more prevalent in the northern versus southern coastal locations, as indicated by the lower evenness (J) estimates (p < 0.05). Indeed, the majority of the canopy tree regeneration in the northern coastal sites consisted of either Abies balsamea, Picea mariana, or Betula papyrifera (Table 3). The continental sites had intermediate J values and did not differ from either coastal location. Alpha, β, γ, and H’ values were greater for the sub-canopy tree and shrub layer relative to the canopy tree re- generation within and among all geographical locations and for across the region as a whole. Discussion Factors Influencing Vegetation Distribution across the Forest Region While different factors influenced post-disturbance commu- nity composition among locations across the region, relative species compositions of the tree canopy were either maintained or were developing towards dominance by the shade-tolerant conifer, Abies balsamea. The likely mechanisms by which this is occurring are via the competitive advantage provided by an abundant advance regeneration layer and the protection pro- vided by coarse woody debris. Because the disturbances examined here did not cause exten- sive damage to the understory vegetation present prior to dis- turbance, stand composition after natural disturbance was dominated by Abies balsamea, a species that develops an abundant advance regeneration layer over time (Morin, 1994). The presence of advance regeneration reduced the ability of shade-intolerant deciduous species to establish after natural disturbance by limiting available growing space, light, and other resources (Morin et al., 2008). Density of coarse woody debris was the only environmental factor significantly affecting regeneration patterns across the entire region. Its irregular spatial distribution and variation in level of decay throughout the landscape was important for the creation of stand heterogeneity, to which the various species responded. For example, shade-tolerant conifer tree species such as Abies balsamea and Picea mariana dominated areas where the density of coarse woody debris was high. Coarse woody debris was beneficial for the survival of the advance regeneration perhaps by mitigating the adverse changes in mi- crosite conditions after disturbance by providing cover and shade (Gray & Spies, 1997; Elliott et al., 2002). Consequently, cyclical regeneration patterns (Baskerville, 1975) were ob- served in conifer-dominated stands. Conversely, areas where coarse woody debris densities were low or where it was absent altogether allowed for the establishment of more shade-intol- erant deciduous species such as Populus tremuloides, Betula Copyright © 2013 SciRes. 94 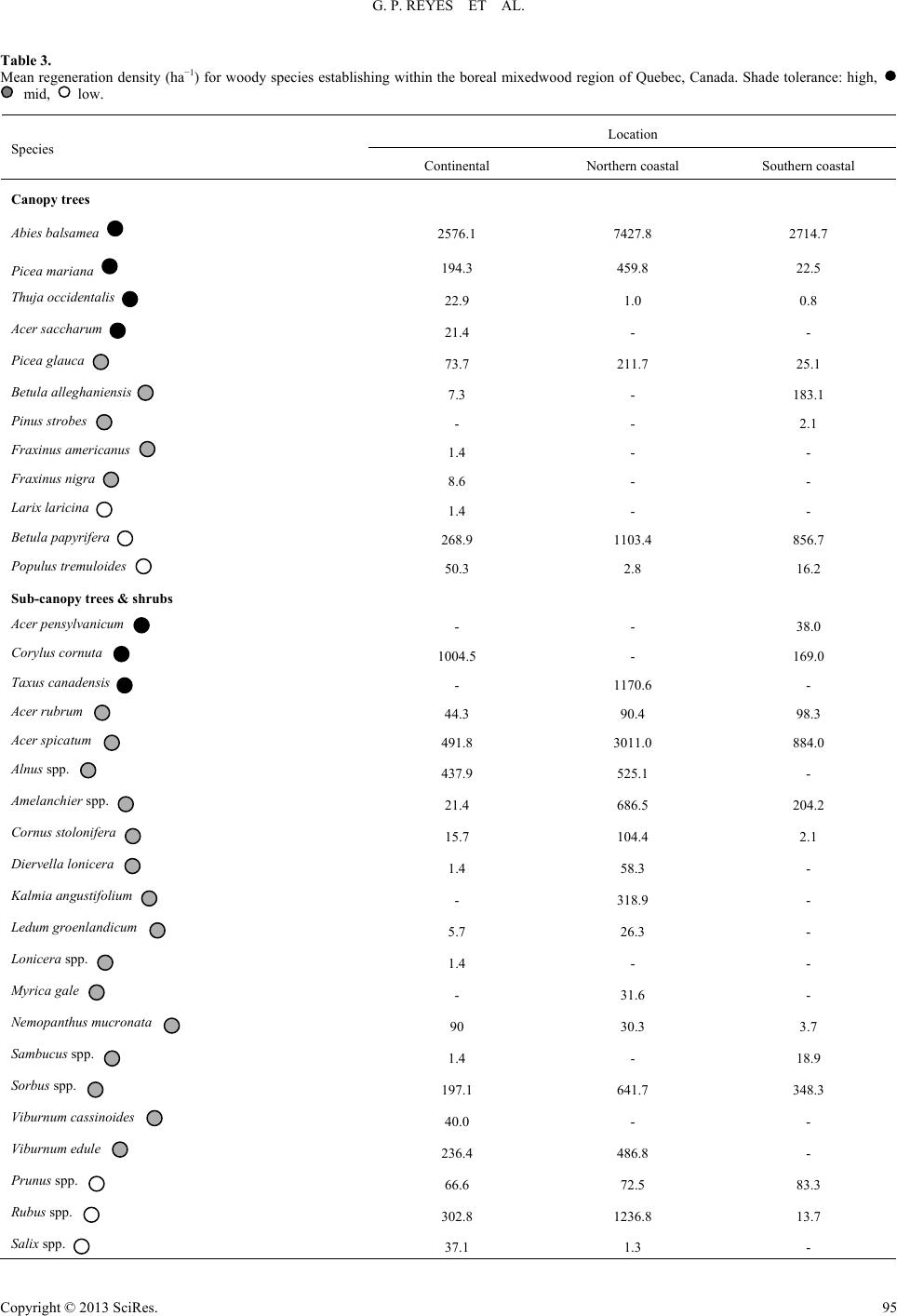 G. P. REYES ET AL. Table 3. Mean regeneration density (ha−1) for woody species establishing within the boreal mixedwood region of Quebec, Canada. Shade tolerance: high, mid, low. Location Species Continental Northern coastal Southern coastal Canopy trees Abies balsamea 2576.1 7427.8 2714.7 Picea marian a 194.3 459.8 22.5 Thuja occidentalis 22.9 1.0 0.8 Acer saccharum 21.4 - - Picea glauca 73.7 211.7 25.1 Betula alleghaniensis 7.3 - 183.1 Pinus strobes - - 2.1 Fraxinus americanus 1.4 - - Fraxinus ni gra 8.6 - - Larix laricina 1.4 - - Betula papyrifera 268.9 1103.4 856.7 Populus tremuloides 50.3 2.8 16.2 Sub-canopy trees & shrubs Acer pensylvanicum - - 38.0 Corylus cornuta 1004.5 - 169.0 Taxus cana densis - 1170.6 - Acer rubrum 44.3 90.4 98.3 Acer spicatum 491.8 3011.0 884.0 Alnus spp. 437.9 525.1 - Amelanchier spp. 21.4 686.5 204.2 Cornus stolo nifera 15.7 104.4 2.1 Diervella lonicera 1.4 58.3 - Kalmia angustifolium - 318.9 - Ledum groenlandicum 5.7 26.3 - Lonicera spp. 1.4 - - Myrica gale - 31.6 - Nemopant hus mucronat a 90 30.3 3.7 Sambucus spp. 1.4 - 18.9 Sorbus spp. 197.1 641.7 348.3 Viburnum cassinoides 40.0 - - Viburnum edule 236.4 486.8 - Prunus spp. 66.6 72.5 83.3 Rubus spp. 302.8 1236.8 13.7 Salix spp. 37.1 1.3 - Copyright © 2013 SciRes. 95 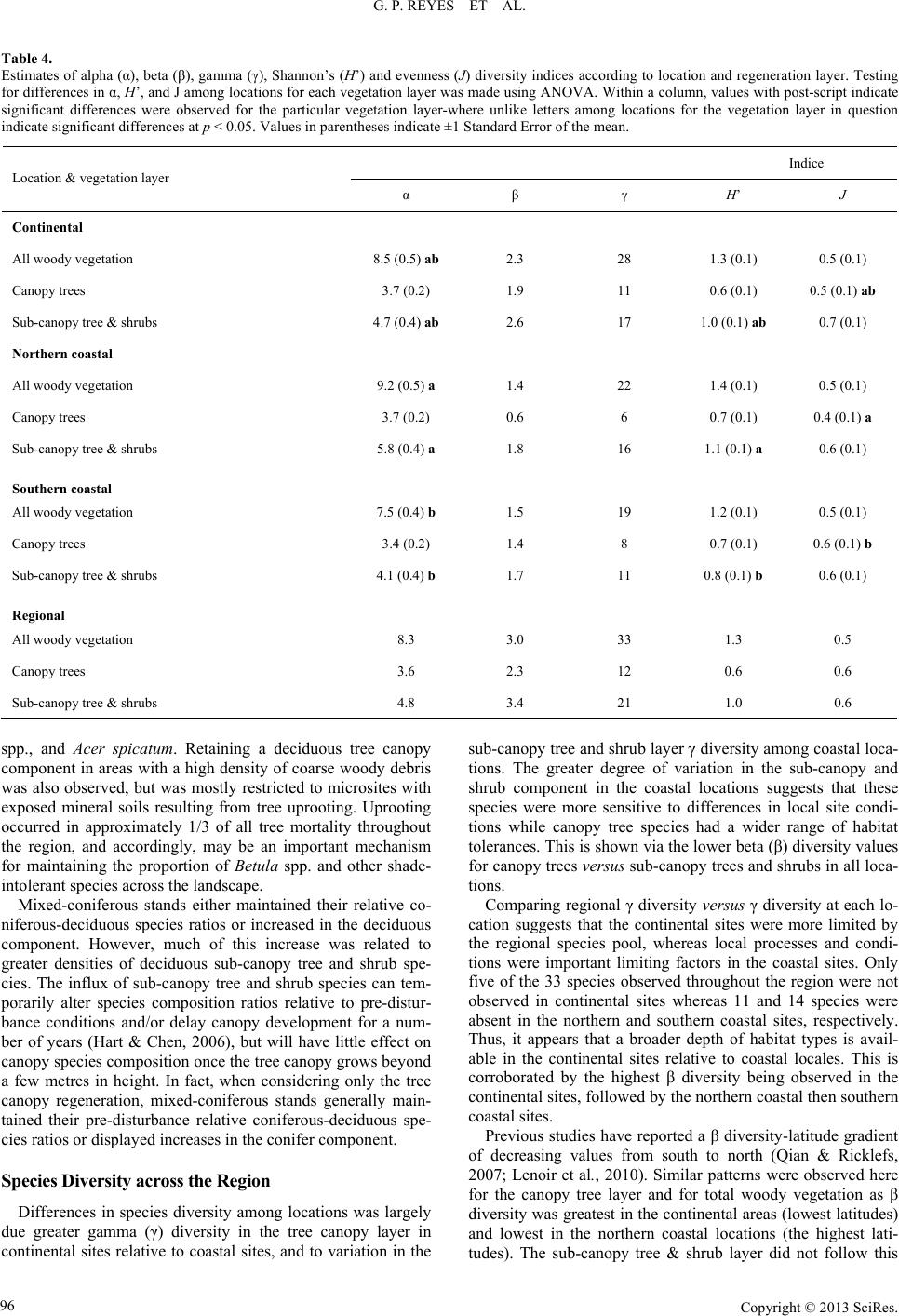 G. P. REYES ET AL. Table 4. Estimates of alpha (α), beta (β), gamma (γ), Shannon’s (H’) and evenness (J) diversity indices according to location and regeneration layer. Testing for differences in α, H’, and J among locations for each vegetation layer was made using ANOVA. Within a column, values with post-script indicate significant differences were observed for the particular vegetation layer-where unlike letters among locations for the vegetation layer in question indicate significant differences at p < 0.05. Values in parentheses indicate ±1 Standard Error of the mean. Indice Location & vegetation layer α β γ H’ J Continental All woody vegetation 8.5 (0.5) ab 2.3 28 1.3 (0.1) 0.5 (0.1) Canopy trees 3.7 (0.2) 1.9 11 0.6 (0.1) 0.5 (0.1) ab Sub-canopy tree & shrubs 4.7 (0.4) ab 2.6 17 1.0 (0.1) ab 0.7 (0.1) Northern coastal All woody vegetation 9.2 (0.5) a 1.4 22 1.4 (0.1) 0.5 (0.1) Canopy trees 3.7 (0.2) 0.6 6 0.7 (0.1) 0.4 (0.1) a Sub-canopy tree & shrubs 5.8 (0.4) a 1.8 16 1.1 (0.1) a 0.6 (0.1) Southern coastal All woody vegetation 7.5 (0.4) b 1.5 19 1.2 (0.1) 0.5 (0.1) Canopy trees 3.4 (0.2) 1.4 8 0.7 (0.1) 0.6 (0.1) b Sub-canopy tree & shrubs 4.1 (0.4) b 1.7 11 0.8 (0.1) b 0.6 (0.1) Regional All woody vegetation 8.3 3.0 33 1.3 0.5 Canopy trees 3.6 2.3 12 0.6 0.6 Sub-canopy tree & shrubs 4.8 3.4 21 1.0 0.6 spp., and Acer spicatum. Retaining a deciduous tree canopy component in areas with a high density of coarse woody debris was also observed, but was mostly restricted to microsites with exposed mineral soils resulting from tree uprooting. Uprooting occurred in approximately 1/3 of all tree mortality throughout the region, and accordingly, may be an important mechanism for maintaining the proportion of Betula spp. and other shade- intolerant species across the landscape. Mixed-coniferous stands either maintained their relative co- niferous-deciduous species ratios or increased in the deciduous component. However, much of this increase was related to greater densities of deciduous sub-canopy tree and shrub spe- cies. The influx of sub-canopy tree and shrub species can tem- porarily alter species composition ratios relative to pre-distur- bance conditions and/or delay canopy development for a num- ber of years (Hart & Chen, 2006), but will have little effect on canopy species composition once the tree canopy grows beyond a few metres in height. In fact, when considering only the tree canopy regeneration, mixed-coniferous stands generally main- tained their pre-disturbance relative coniferous-deciduous spe- cies ratios or displayed increases in the conifer component. Species Diversity across the Region Differences in species diversity among locations was largely due greater gamma (γ) diversity in the tree canopy layer in continental sites relative to coastal sites, and to variation in the sub-canopy tree and shrub layer γ diversity among coastal loca- tions. The greater degree of variation in the sub-canopy and shrub component in the coastal locations suggests that these species were more sensitive to differences in local site condi- tions while canopy tree species had a wider range of habitat tolerances. This is shown via the lower beta (β) diversity values for canopy trees versus sub-canopy trees and shrubs in all loca- tions. Comparing regional γ diversity versus γ diversity at each lo- cation suggests that the continental sites were more limited by the regional species pool, whereas local processes and condi- tions were important limiting factors in the coastal sites. Only five of the 33 species observed throughout the region were not observed in continental sites whereas 11 and 14 species were absent in the northern and southern coastal sites, respectively. Thus, it appears that a broader depth of habitat types is avail- able in the continental sites relative to coastal locales. This is corroborated by the highest β diversity being observed in the continental sites, followed by the northern coastal then southern coastal sites. Previous studies have reported a β diversity-latitude gradient of decreasing values from south to north (Qian & Ricklefs, 2007; Lenoir et al., 2010). Similar patterns were observed here for the canopy tree layer and for total woody vegetation as β diversity was greatest in the continental areas (lowest latitudes) and lowest in the northern coastal locations (the highest lati- tudes). The sub-canopy tree & shrub layer did not follow this Copyright © 2013 SciRes. 96  G. P. REYES ET AL. pattern as β diversity was higher in the northern versus southern coastal sites. Further, α and γ diversity for sub-canopy tree & shrubs did not follow this latitudinal pattern along the coast either, as these diversity components were also higher in north- ern coastal sites. Other site-level factors, such as density of coarse woody debris may have been important in influencing regeneration distribution. Differences in β diversity among locations coincided with coarse woody debris and snag densi- ties being lowest in the continental sites and the highest in the southern coastal sites, which would suggest greater availability of growing space in the continental areas and lowest available space in the southern coastal sites. Conclusion The effects of windthrow, spruce budworm, and interaction natural disturbances on local site conditions were not severe enough to alter canopy species dominance. Dominance by the main canopy species, Abies balsamea and Betula spp. was maintained throughout the region, suggesting that these species had a wider tolerance range and wider niche overlap relative to other woody species, and/or that the severity of these natural disturbances wasn’t sufficient to alter succession trajectories. Moreover, differences in the importance of factors influencing responses of the woody vegetation among locations across the boreal mixedwood region reinforces the concept that species distribution is not controlled by natural disturbance alone, but that local environmental characteristics and constraints on plant biology also dictate re-establishment success. Each location across the region had different and unique site characteristics, and accordingly, species composition and abundance varied from one location to the next. Thus, while the specific distur- bance types did not considerably alter canopy tree composition, as development towards old-growth, conifer-dominated condi- tions was maintained, these natural disturbances were important for creating a mosaic of structural legacies that promote stand heterogeneity, which can ultimately help to maintain biodiver- sity in the region. Acknowledgements Many thanks to all my field assistants (J.-F. Mnudles-Gagnon, Maude Beauregard, David Saucier, Julie Messierer, and Isabelle Nault). Your various personalities, quirks, and habits that were amplified during long, buggy, hot and/or rainy days in the field made data collection quite memorable and fun! Additionally, this study would not have been possible without financial and/or technical support from TEMREX, NSERC- CFS and the SFMN. REFERENCES Baskerville, G. L. (1975). Spruce budworm: Super silviculturist. The Forestry Chronicle, 51, 138-140. Belote, R. T., Sanders, N. J., & Jones, R. H. (2009). Disturbance alters local-regional richness relationships in Appalachian forests. Ecology, 90, 2940-2947. doi:10.1890/08-1908.1 Bergeron, Y. (2000). Species and stand dynamics in the mixed woods of Quebec’s southern boreal forest. Ecology, 81, 1500-1516. doi:10.1890/0012-9658(2000)081[1500:SASDIT]2.0.CO;2 Bergeron, Y., Cyr, D., Drever, R., Flannigan, M., Gauthier, S., Knee- shaw, D., Lauzon, E., Leduc, A., Le Goff, H., Lesieur, D., & Logan, K. (2006). Past, current, and future fire frequencies in Quebec’s commercial forests: Implications for the cumulative effects of har- vesting and fire on age-class structure and natural disturbance-based management. Canadian Journal of F ores t Research, 36, 2737-2744. doi:10.1139/x06-177 Chen, H. Y. H., & Popadiouk, R. V. (2002). Dynamics of North Ame- rican boreal mixedwoods. Environmental Rev i ews, 10, 137-166. doi:10.1139/a02-007 Dansereau, P. (1954). Climax vegetation and the regional shift of con- trols. Ecology, 35, 575-579. doi:10.2307/1931048 Dupont, A., Belanger, L., & Bousquet, J. (1991). Relationships between balsam fir vulnerability to spruce budworm and ecological site con- ditions of fir stands in central Quebec. Canadian Journal of Forest Research, 21, 1752-1759. doi:10.1139/x91-242 Elliott, K. J., Hitchcock, H. L., & Krueger, L. (2002). Vegetation re- sponse to large scale disturbance in a southern Appalachian forest: Hurricane Opal and salvage logging. Journal of the Torrey Botanical Society, 129, 48-59. doi:10.2307/3088682 Environment Canada (2004). Canadian climate normals or averages 1971-2000. URL (last checked 24 July 2006). http://climate.weatheroffice.ec.gc.ca/climate_normals/index_e.html Gray, A. N., & Spies, T. A. (1997). Microsite controls on tree seedling establishment in conifer forest canopy gaps. Ecology, 78, 2458-2473. doi:10.1890/0012-9658(1997)078[2458:MCOTSE]2.0.CO;2 Gray, A. N., Zald, H. S. J., Kern, R. A., & North, M. (2005). Stand conditions associated with tree regeneration in sierran mixed-conifer forests. Forest Science, 51, 198-210. Gromtsev, A. (2002). Natural disturbance dynamics in the boreal for- ests of European Russia: A review. Silva Fennica, 36, 41-55. Hammer, O., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleon- tological statistics software package for education and data analysis. Palaeontologia Electronica, 4, 1-9. Hart, S. A., & Chen, H. Y. H. (2006). Understory vegetation dynamics of North American boreal forests. Critical Reviews in Plant Sciences, 25, 381-397. doi:10.1080/07352680600819286 Hillebrand, H., & Blenckner, T. (2002). Regional and local impact on species diversity—From pattern to process. Oecologia, 132, 479-491. doi:10.1007/s00442-002-0988-3 Huebner, C. D., & Vankat, J. L. (2003). The importance of environment vs. disturbance in the vegetation mosaic of Central Arizona. Journal of Vegetation Science, 14, 25-34. doi:10.1111/j.1654-1103.2003.tb02124.x Imbeau, L., & Desrochers, A. (2002). Foraging ecology and use of drumming trees by three-toed woodpeckers. Journal of Wildlife Ma- nagement, 66, 222-231. doi:10.2307/3802888 Krasny, M. E., & Whitmore, M. C. (1992). Gradual and sudden forest canopy gaps in Allegheny northern hardwood forests. Canadian Journal of Forest Research, 22, 139-143. doi:10.1139/x92-019 Krebs, C. J. (1989). Ecological methodology. New York, NY: Harper and Row Publishers Inc., 654 p. Kuuluvainen, T., & Juntunen, P. (1998). Seedling establishment in rela- tion to microhabitat variation in a windthrow gap in a boreal Pinus sylvestris forest. Journal of Vegetation Science, 9, 551-562. doi:10.2307/3237271 Lenoir, J., Gégout, J.-C., Guisan, A., Vittoz, P., Wohlgemuth, T., Zim- mermann, N. E., Dullinger, S., Pauli, H., Willner, W., Grytnes, J.-A., Virtanen, R., & Svenning, J.-C. (2010) Crossscale analysis of the re- gion effect on vascular plant species diversity in southern and north- ern European mountain ranges. PLoS ONE, 5, e15734. doi:10.1371/journal.pone.0015734 McGuire, A. D., Wirth, C., Apps, M., Beringer, J., Clein, J., Epstein, H., Kicklighter, D. W., Bhatti, J., Chapin III, F. S., de Groot, B., Efre- mov, D., Eugster, W., Fukuda, M., Gower, T., Hinzman, L., Huntley, B., Jia, G. J., Kasischke, E., Melillo, J., Romanovsky, V., Shvidenko, A., Vaganov, E., & Walker, D. (2002). Environmental variation, ve- getation distribution, carbon dynamics and water/energy exchange at high latitudes. Journal of Vegetation Science, 13, 301-314. doi:10.1111/j.1654-1103.2002.tb02055.x Messaoud, Y., Bergeron, Y., & Leduc, A. (2007). Ecological factors explaining the location of the boundary between the mixedwood and coniferous bioclimatic zones in the boreal biome of eastern North America. Global Ecology and Biogeography, 16, 90-102. doi:10.1111/j.1466-8238.2006.00277.x Copyright © 2013 SciRes. 97  G. P. REYES ET AL. Copyright © 2013 SciRes. 98 Morin, H. (1994). Dynamics of balsam fir forests in relation to spruce budworm outbreaks in the boreal zone of Quebec. Canadian Journal of Forest Research, 24, 730-741. doi:10.1139/x94-097 Morin, H., Laprise, D., Simard, A.-A., & Amouch, S. (2008). Régime des épidémies de la Tordeuse des bourgeons de l’épinette dans l’Est de l’Amérique du Nord. In Aménagement écosystemique en forêt boréale (pp. 165-192). Quebec: University of Quebec Press. MRNQ. (2003). Ministère des resources naturelles du Quebec. URL (last checked 24 July 2006). http://www.mrn.gouv.qc.ca Nagel, T. A., & Diaci, J. (2006). Intermediate wind disturbance in an old-growth beech-fir forest in southwestern Slovenia. Canadian Journal of Forest Research, 36, 629-638. doi:10.1139/x05-263 Neilson, R. P., King, G. A., De Velice, R. L., & Lenihan, J. M. (1992). Regional and local vegetation patterns: The responses of vegetation diversity to subcontinental air masses. In Ecological Studies 92: Landscape boundaries: consequences for biotic diversity and eco- logical flows (pp. 129-149). New York: Springer-Verlag. Novotny, V., & Weiblen, D. B. (2005). From communities to conti- nents: beta diversity of herbivorous insects. Annales Zoologici Fen- nici, 42, 463-475. Osumi, K., Ikeda, S., & Okamoto, T. (2003). Vegetation patterns and their dependency on site conditions in the pre-industrial landscape on north-eastern Japan. Ecological Research, 18, 753-765. doi:10.1111/j.1440-1703.2003.00593.x Palmer, M., & White, P. S. (1994). Scale dependence and the species- area relationship. American Naturalist, 144, 717-740. doi:10.1086/285704 Peterson, C. J. (2000). Catastrophic wind damage to North American forests and the potential impact of climate change. The Science of the Total Environment, 262, 287-311. doi:10.1016/S0048-9697(00)00529-5 Pielou, E. C. (1966). The measurement of diversity in different types of biological collections. Journal of Theoretical Biology, 13, 131-144. doi:10.1016/0022-5193(66)90013-0 Qian, H., & Ricklefs, R. E. (2007). A latitudinal gradient in large-scale beta diversity for vascular plants in North America. Ecology Letters, 10, 737-744. doi:10.1111/j.1461-0248.2007.01066.x Reyes, G., & Kneeshaw, D. (2008). Moderate-severity disturbance dynamics in Abies balsamea-Betula spp. forests: The relative impor- tance of disturbance type and local stand and site characteristics on woody vegetation response. Ecoscie nce, 15, 241-249. doi:10.2980/15-2-3082 Reyes, G., Kneeshaw, D., De Grandpré, L., & Leduc, A. (2010). Chan- ges in woody vegetation abundance and diversity after natural dis- turbances causing different levels of mortality. Journal of Vegetation Science, 21, 406-417. doi:10.1111/j.1654-1103.2009.01152.x Robitaille, A., & Saucier, J. P. (1998). Paysages régionaux du Québec méridional. Québec: Les Publications du Québec. Rodriguez-Garcia, E., Gratzer, G., & Bravo, F. (2011). Climatic vari- ability and other site factor influences on natural regeneration of Pinus pinaster Ait. in Mediterranean forests. Annals of Forest Sci- ence, 68, 811-823. doi:10.1007/s13595-011-0078-y Shannon, C. E., & Weaver, W. (1949). The mathematical theory of communication. Urbana, IL: University of Illinois Press. SPSS Inc. (1999). Professional base system software for statistical analysis (v.10.0). Chicago, Illinois: SPSS Inc. terBraak, C. J. F. & Smilauer, P. (1998). CANOCO reference manual and user’s guide to CANOCO for windows: Software for canonical community ordination (Version 4.02). New York: Microcomputer Power, Ithaca. Turner, M. G., Dale, V. H., & Everham III, E. H. (1997). Crown fires, hurricanes, and volcanoes: A comparison among large-scale distur- bances. BioScience, 47, 758-768. doi:10.2307/1313098 van den Wollenberg, A. L. (1977). Redundancy analysis: An alternative for canonical correlation analysis. Psychometrika, 42, 207-219. doi:10.1007/BF02294050 Veblen, T. T., & Ashton, D. H. (1978). Catastrophic influences on the vegetation of the Valdivian Andes, Chile. Vegetatio, 36, 149-167. doi:10.1007/BF02342598 Wang, X.-P., Tang, Z.-Y., & Fang, J. Y. (2006). Climatic control on forests and tree species distribution in the forest region of northeast China. Journal of Integrative Plant Biology, 48, 778-789. doi:10.1111/j.1744-7909.2006.00294.x Whittaker, R. H. (1960). Vegetation of the Siskiyou Mountains, Oregon and California. Ecological Monogra phs, 30, 279-338. doi:10.2307/1943563 Wimberly, M. C., & Spies, T. A. (2001). Influences of environment and disturbance on forest patterns in coastal Oregon watersheds. Ecology, 82, 1443-1459. doi:10.1890/0012-9658(2001)082[1443:IOEADO]2.0.CO;2
|