N. Tabti, K. Abdellaoui-Hassaïne / Open Journal of Ecology 3 (2013) 265-271
266
to the method recommended by the WHO1 [9] in order to
demonstrate that the bacteriological and physicoche-
mical conditions may play a role in the effectiveness of
Bti.
Moreover, if the evaluation of the effect of insecticide
treatment on larval populations is usually done in the
field, in natural environments, this study analyzes the ef-
fects on populations from breeding after purification.
2. MATERIALS AND METHODS
Toxicological tests are conducted in two ways:
- Populations from livestock breeding: the larval hat-
chlings of hypogean deposits are grouped in lots of
20 larvae at stage 4 placed in cages. Laid eggs are
retrieved in 200 ml crystallizers. Adults (30 males
and 10 females) are evenly distributed in five diffe-
rent cages. The food, provided daily, of the larvae is
composed of a mixture of biscuit and dried yeast
(75/25). This is done in May at an ambient tempera-
ture of 25˚C and a photoperiod 14/10 hours [10].
- Populations sampled from deposits: the larvae from
the four stages are taken directly from three hypogean
deposits (crawl spaces) which are located in different
neighborhoods of Tlemcen urban group. Water intake
is putrid, nauseating and heavily loaded with organic
matter from the leakage of defective pipes (rate of
conductivity).
The bio insecticide used is the standard commercial
formula Vectobac at 200 IUT/mg, granules. The tests were
repeated three times on the same larval stage and were
performed on the same number of larvae (50 larvae of the
same stage), placed in the same volume of water in 1000
ml test tubes, at an ambient temperature between 20 and
25˚C.
The water used is distilled and seven do ses decreasing
from 35 to 2 mg/L were tested on each stage of larvae
from breeding and from 100 to 10 mg/L for those taken
from the deposits. For each test a control group (50 larvae
of the same stage) was placed.
Dead larvae are removed at regular time intervals (30
minutes) until the death of all larvae (100% mortality).
The results are analyzed by a statistical treatment, de-
veloped by the software Minitab12. Mortalities are ex-
pressed as m ean a n d st an dard deviatio n, calculated on t he
percentage mortalities of the three tests and corrected [1 1],
this allows to eliminate th e natural mortality an d to know
the actual larvicide toxicity. Variance analyses with con-
trolled factor were used to demonstrate the effect of time,
the effect of dose and the effect of larval stage. The re-
gression lines are based on Swaroop & Uemera (1966)
method to determine the LD50 and LD90 of the different
larval stages, with a probability of 95%.
3. RESULTS
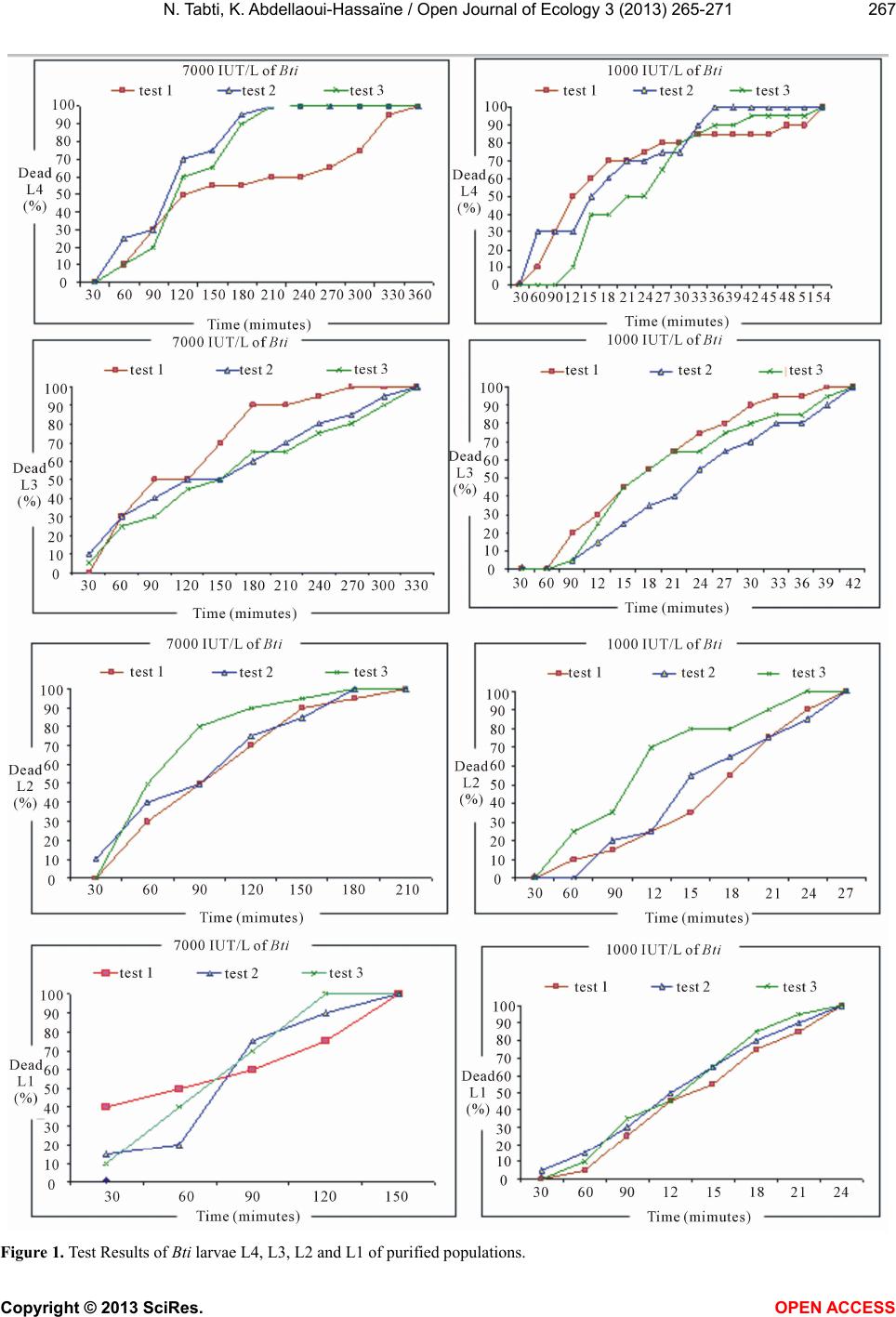
3.1. Determination of the Efficiency of Bti
For all doses tested, the bio insecticide treatment caus-
es a significant lengthening of the duration effect (p <
0.001). It goes from 330 minutes in high doses to 510
minutes for the dose of 50 mg/L.
The Bti time effect is shorter on populations from
breeding than on populations taken directly from depo-
sits (Figur es 1 and 2).
The analysis of variance shows that under the effect of
the same doses, the mortality rates recorded 360 minutes
after treatment of larvae are high (Figures 1 and 2). In
the four larval stages (p < 0.005), there wa s a highly sig-
nificant difference between the average mortality for all
instars.
Taking into account the statistical analysis, it is clear
that the Bti has effects on larval mortality, especially for
higher doses, where the larvae are all dead.
The first stage treated larvae, appear to be more sen-
sitive to Bti, due to mortality rates recorded for the dif-
ferent concentrations. For larval stages L2, L3 and L4,
doses act the same way on the mortality of larvae as well
as purified populations of artificial deposits.
The effectiveness of the two factors “dose” and “larval
stage” is demonstrated through an analysis of variance
with two controlled factors, encompassing the four larval
stages and the different doses tested. This analysis shows
the impact of each of the two factors; the larval stage and
the different doses used, significantly, affect larval mor-
tality of Cx. pipiens: probability p < 0.005 (Table 1).
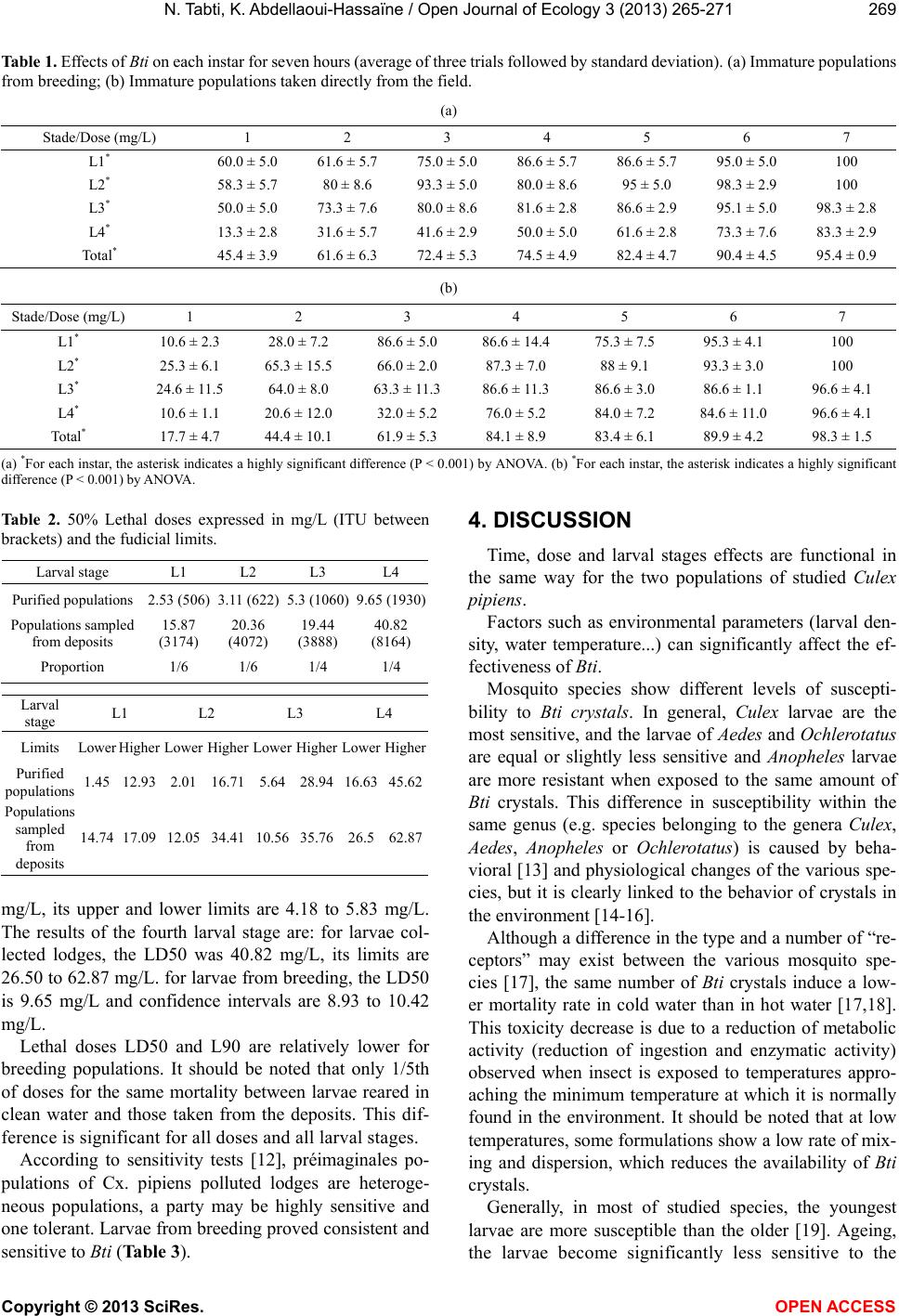
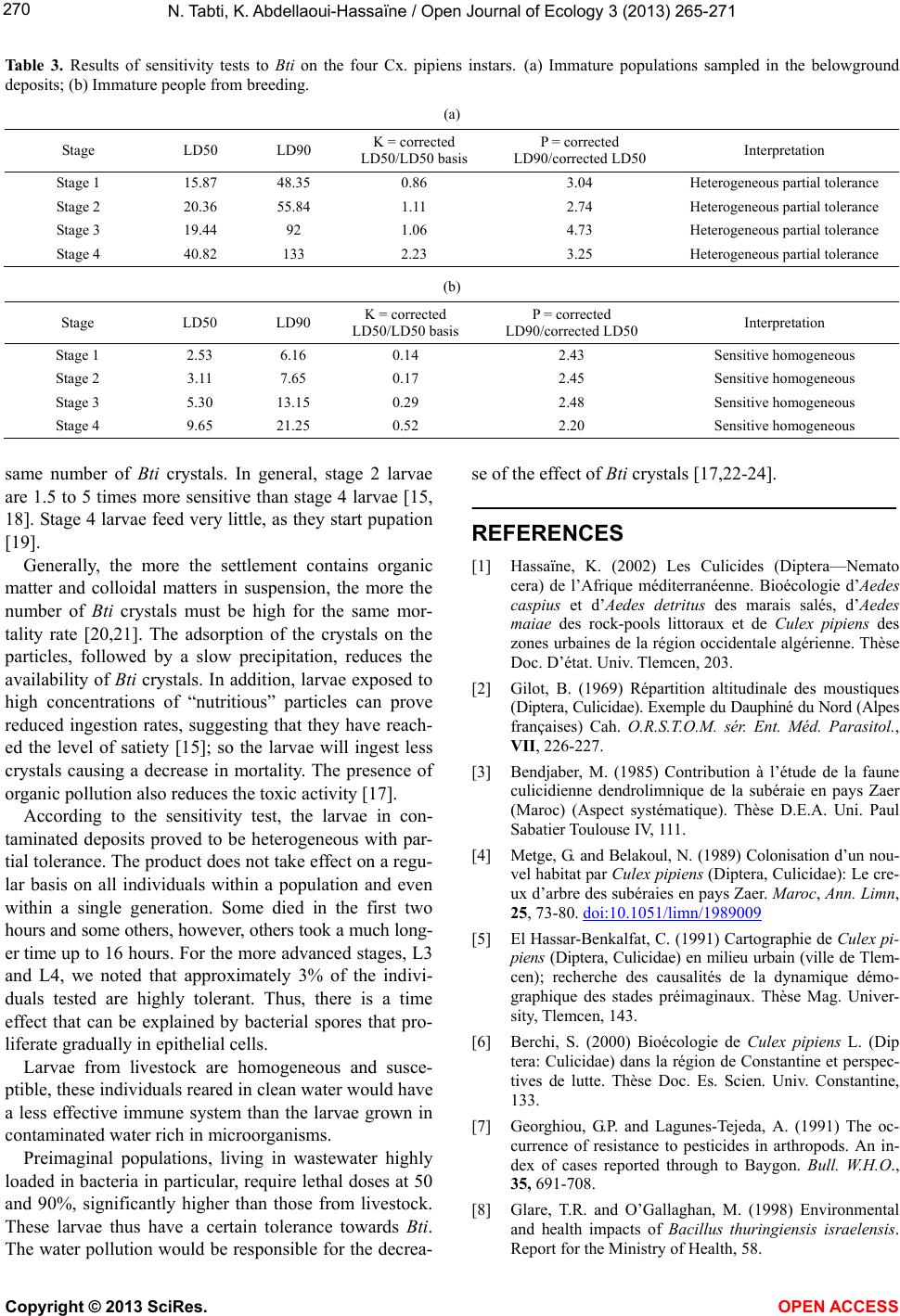
3.2. Determination of Lethal Dose (Table 2)
The coefficients of the carried out eight regression
lines show that there is a significant relation ship between
the dead larvae and the Bti different dosages.
The probit analysis allowed us to retain 50% lethal
dose (LD50) of Bti which is of the order of 15.87 mg/L
for the first larval stage of the breeding population col-
lected from polluted deposits, its upper and lower limits
are respectively upper 14.74 and 17.09 m g/L. for t his same
level of rearing larvae from the LD50 is 2.53 mg/L, its
upper and lower limits are 2.26 to 2.83 mg/L.
For larvae of the second stage, the LD50 of larvicide is
20.36 mg/L. Confidence intervals of LD50 are from 12.05
to 34.41 mg/L (larvae from polluted deposits). For larvae
from breeding, the LD50 is 3.11 mg/L and confidence
intervals are from 2.8 to 3.45 mg/L.
The lethal dose for 50% mortality for larvae stage 3 is
19.44 mg/L. The lower limit is 10.56 mg/L and the upper
limit is 35.76 mg/L, this for individuals collected directly
from shelters. LD50 populations from livestock is 5.3
1international toxicity units.
2lethal dose for 50% mortality.
3World Health Organization.
Copyright © 2013 SciRes. OPEN A CCESS