 Vol.3, No.3, 215-223 (2013) Open Journal of Ecology http://dx.doi.org/10.4236/oje.2013.33025 Stand age structural dynamics of conifer, mixedwood, and hardwood stands in the boreal forest of central Canada Jennifer M. Fricker1, Jian R. Wang1*, H. Y. H. Chen1, Peter N. Duinker2 1Faculty of Natural Resources Management, Lakehead University, Thunder Bay, Canada; *Corresponding Author: jian.wang@lakeheadu.ca 2School for Resource and Environmental Studies, Dalhousie University, Halifax, Canada Received 18 April 2013; revised 26 May 2013; accepted 28 June 2013 Copyright © 2013 Jennifer M. Fricker et al. This is an open access article distributed under the Creative Commons Attribution Li- cense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT To study the effects of stand development and overstory composition on stand age structure, we sampled 32 stands representing conifer, mix- edwood, and hardwood stand types, ranging in ages from 72 to 201 years on upland mesic sites in northwestern Ontario. We defined the stages of stand development as: stem exclusion/cano- py transition, canopy transition, canopy transi- tion/gap dynamics, and g ap d ynamics. Stand age structure of conifer stands changed from bimo- dal, bimodal, reverse-J, and bimodal, respe ctive- ly, through the stages of stand development. Mixedwood and hardwood stands revealed simi- lar trends, with the exception of missing the ca- nopy transition/gap dynamic stage in mixedwoods. Canopy transition/gap dynamic stage in hard- woods showed a weaker reverse-J distribution than their conifer counterparts. The results sug- gest that forest management activities such as partial and selection harvesting and seed-tree systems may diversify standard landscape-level age structures and benefit wildlife, hasten the onset of old-growth, and create desired stand age structures. We also recommend that the de- termination of old-growth using the following criteria in the boreal forest: 1) canopy break- down of pioneering coh ort is comp le te and stand is dominated by later successional tree species, and 2) stand age structure is bimodal, with do- minating canopy trees that fall w ithin a relatively narrow range of age and height classes and a sig- nificant amount of understory regeneration. Keywords: Time Since Fire (TSF); Stand Development; Old-Growth F orest; Conife rs; Hardwoods; Mixedwood; Boreal Forests 1. INTRODUCTION Forest stands have long been described by their age structure and diameter distribution. Ecosystem dynamics in the Canadian boreal forest are closely tied to natural fire regimes. Fire types, intensity and time since fire (TSF) are fundamental to forest species composition, age structure and forest succession [1-3]. Forest managers are challenged and mandated by law in some jurisdic- tions to preserve and emulate natural disturbances such as fire in the boreal forest region. Therefore the dynamic patterns of age structure are the basis for landscape level planning in Northwest Ontario. Age structures of natural forest stands change over time [4,5]. Research has found that forest stands change from an even-aged, relatively homogeneous tree height structure to a two-cohorts, bimodal height structure to where tree heights are relatively heterogeneous as TSF increases [5-8]. Young stands are primarily composed of early successional species that grow quickly in open ar- eas in full light [9]. However, as stand age increases, stands become increasingly composed of later succes- sional species that can establish under a closed canopy with limited light. Age structure has been examined for conifer [10], mix- edwood [9], and hardwood forest types [11,12] in a spe- cific successional stage. However, few studies thus far have compared age structure for different boreal forest cover types with similar environmental characteristics (i.e., soils, topography and climate) along a successional gradient. As many stand characteristics have been found to differ with stand composition (i.e., productivity, coarse woody debris) [13-16], we hypothesized that the differ- Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 2 15-223 216 ent stand cover types would vary in stand age structural dynamics. Old-growth forests have potential wildlife habitat and biodiversity benefits that link to unique structural char- acteristics i.e., larger trees, a multi-layered canopy, cano- py gaps, and higher tree species richness [17]. However, few definitions exist to identify old-growth in the boreal forest, and this lack of information and confusion sur- rounding the old-growth condition is hindering manage- ment planning and implementation [18]. The goal of this study is to determine how stand age structure changes in the boreal forest over time. Specifi- cally, we addressed how stand age structure varies with forest cover type, conifer, mixedwood, and hardwood and developmental stages, stem exclusion/canopy transition, canopy transition, canopy transition/gap dynamics, and gap dynamics [1]. 2. MATERIALS AND METHODS 2.1. Study Area We conducted this study in northwestern Ontario bo- real forests north of Lake Superior in the Superior (B.9) Forest Region [19], Ontario (48˚ 22’ N, 89˚ 19’ W, 199 m altitude) [20] in the Spruce River Forest. Climatically, the area is influenced by Lake Superior and has a moder- ately dry, cool climate with a short summer. The average annual precipitation for Thunder Bay is 712 mm with an average annual temperature of 2.5˚C. Topographical fea- tures were formed during the retreat of the Laurentide Ice Sheet approximately 10,000 years ago [20]. The area is characterized as containing tree species of paper birch (Betula papyrifera Marsh.), trembling aspen (Populus tremuloides Michx.), balsam fir (Abies bal- samea (L.) Mill.), white spruce (Picea glauca (Moench) Voss), black spruce (Picea mariana (Mill.) BSP), jack pine (Pinus banksiana Lamb), and eastern white cedar (Thuja occidentalis L.) on sites [19]. Common shrubs and herbs found in this area were mountain maple (Acer spicatum Lam.), beaked hazel (Corylus cornuta Marsh.), labrador tea (Ledum groenlandicum Jacq.), Canada fly honeysuckle (Lonicera canadensis Bart. Ex Marsh.), nor- thern star flower (Trientalis borealis Raf.), rose twisted stalk (Streptopus roseus Michx.), bunchberry (Cornus ca- nadensis L.), and wild lily of the valley (Maianthemum canadense Desf.). The natural stand-initiating distur- bance in the area is predominately stand-replacing fire with a fire return interval of approximately 100 years [21]. 2.2. Sampling Design Thirty two stands were sampled throughout the study area in a stratified (by forest type) random manner. We sampled three overstory types: 1) conifer dominated by jack pine at early stages of development with a mixture of black spruce, white spruce, and balsam fir at later stages of stand development; 2) hardwood dominated by trembling aspen at early stages of stand development and paper birch at later stages; and 3) mixedwood dominated by a mixture of jack pine and trembling aspen in early stages of stand development and a mixture of black spruce, white spruce, balsam fir, and paper birch in later stages. The sampled stands aged from 72 to 201 years TSF (Table 1). Stand type was determined through a modification of methods used by [22]. Stands were assessed as belonging to a specific stand type based on the density (stems/plot) of conifer trees that dominated the overstory of the stand. Stands with a greater than 75% conifer component were classified as “conifer type”, stands with a 25% - 75% conifer component were classified as “mixedwood type”, and stands with less than a 25% conifer component were classified as “hardwood type”. All sampled stands were fire-origin on prevailing me- sic, upland sites in order to represent the forests in the re- gion and limit soil variability. Soil order and texture were determined by excavating three soil pits using methods outlined by British Columbia Ministry of Environment & British Columbia Ministry of Forests [23]. Soil assess- ment followed [24,25] to ensure that sites met the selec- tion criteria described previously. For all sites, soil order was Brunisol while soil texture was sandy loam, sandy clay loam, or clay loam. 2.3. Field Measurements Within each stand, a 400 m2 circular plot was estab- Table 1. Description of 32 sampled stands in northwestern On- tario. Density (trees/ha) Number of stands Stand type*Stage† TSF (year) Canopy trees‡ Regeneration‡ 3 C 2 - 372 1217 (253) § 3955 (1142) 3 C 3 90 1208 (158) 3022 (387) 2 C 3 - 4139 487 (62) 2666 (133) 3 C 4 201 908 (260) 1288 (437) 3 M 2 - 372 1083 (72) 2311 (823) 3 M 3 90 1108 (144) 2222 (898) 3 M 4 201 1066 (96) 3777 (1646) 3 H 2 - 372 1158 (375) 1111 (512) 3 H 3 90 675 (14) 177 (117) 3 H 3 - 4139 716 (41) 2088 (270) 3 H 4 201 1025 (203) 2866 (1404) *Stand type: C = conifer, M = mixedwood, H = hardwood; †Stand develop- mental stage: 2 - 3 = stem exclusion/canopy transition, 3 = canopy transition, 3 - 4 = canopy transition/gap dynamics, 4 = gap dynamics; ‡Canopy trees are ≥10 cm diameter at breast height (DBH), regeneration are trees <10 cm DBH; §Numbers in brackets equal one standard error of the mean. Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 215-223 217 lished to represent the stand. Within each plot, the di- ameter at breast height (DBH) (1.3 m above the root col- lar) and species of all trees (DBH ≥10 cm) were meas- ured and recorded. The DBH measurements were then grouped into DBH classes (10 - 14.9 cm, 15 - 19.9 cm, 20 - 24.5 cm, 25 - 29.9 cm, and ≥ 30 cm) to facilitate tree selection for height and age measurement. Five trees (if available) were randomly selected from each 5-cm DBH class for each species. For each sample tree, its height was measured using a clinometer, and an increment core was taken at breast height. Increment cores were stored in a freezer until they processed. Three circular 25 m2 subplots were randomly selected within each 400 m2 plot to evaluate natural regeneration (<10 cm DBH). This would also include trees that had not yet reached breast height. Within each subplot, di- ameter at the root collar, height, and species of all trees were measured and recorded. Further, trees were grouped into height classes (<0.29 m, 0.3 - 0.99 m, 1.0 - 1.9 m, 2.0 - 4.9 m, and ≥5 m), and 5 trees were randomly se- lected from each height class, and a disk was taken at the root collar for accurate aging of each sapling and seed- ling. 2.4. Tree Ring Counting In the lab, increment cores from trees DBH ≥10 cm were each mounted on a wood strip and sanded with grit sandpaper until the rings were visible. Growth rings were then counted using a hand-held magnifier. To estimate the age of each tree from root collar to breast height, a species-specific number of years was added to each tree’s growth ring count as outlined by [26] (jack pine = 8 years, trembling aspen and paper birch = 7 years, black and white spruce and balsam fir = 18 years). Balsam fir and white spruce were based on a conservative estimate of the ages of trees using black spruce because it is more shade-tolerant. For trees DBH <10 cm, the growth rings of each disk were counted using a hand-held magnifier or under a microscope. As the disk was taken at the root collar, no ages had to be added. 2.5. Data Analysis Ages for the remaining trees were estimated using spe- cies-specific non-linear regression models as outlined by [27] developed from the paired age and diameter meas- urements, 100110 LogLog diameterAa a where A is tree age (years), a0 and a1 are parameters, and diameter is diameter at (a) breast height (if tree DBH ≥10 cm) or (b) root collar (if tree DBH <10 cm). The para- meter estimates for the age-diameter models were pre- sented in Table 2. Table 2. Species-specific parameter estimates of non-linear age-diameter at breast height models using Daniels et al. (1995) ( 01 10^ (* Log10diameterAaa ) where A is tree age (years), a0 and a1 are parameters, and diameter is diameter at (a) breast height (if tree DBH ≥10 cm) or (b) root collar (if tree DBH <10 cm). For trees DBH <10 cm, no jack pine were sam- pled, while the sample size for trembling aspen was very small. = Therefore, no age-diameter models for those species were developed. Parameter Species a0 a1 MS R2 Trees (DBH ≥10 cm) Bf 1.7265 0.0958 71.6835 0.0496 Bw 1.4499 0.3034 101.1264 0.2529 Pj 1.4472 0.309 178.5015 0.1715 Po 1.2813 0.4364 156.3646 0.3833 Sb 1.6816 0.1729 153.9571 0.1015 Sw 1.5596 0.2406 116.9068 0.3256 Trees (DBH <10 cm) Bf 1.2965 0.459 48.4667 0.5297 Bw 1.1645 0.2477 57.9858 0.2668 Sb 1.3301 0.5539 24.2526 0.6614 Sw 1.2944 0.4908 11.235 0.6537 Trees by species were then grouped into age classes as follows: 1) 0 - 9 years, 2) 10 - 19 years, 3) 20 - 29 years, 4) 30 - 39 years, 5) 40 - 49 years, 6) 50 - 59 years, 7) 60 - 69 years, 8) 70 - 79 years, 9) 80 - 89 years, 10) 90 - 99 years, 11) 100 - 109 years, 12) 110 - 119 years, and 13) ≥ 120 years and scaled up to stems per hectare. Bar charts were constructed to show the density of trees (trees/ha) by age class and species in each stand developmental stage and forest cover type. 3. RESULTS 3.1. Conifer Stands Stand age structure in the stem exclusion/transition stage was largely bimodal, having a younger cohort in the 2 and 3 age classes and an older cohort in the 7 and 8 age classes (Figure 1(a)). The younger cohort was com- posed of mainly balsam fir and black spruce which re- cruited in the understory at various times after the stand- replacing fire. The older cohort was composed of mainly jack pine canopy trees with some black spruce, balsam fir and paper birch that established shortly after fire, therefore falling within a relatively narrow range of age classes. Canopy tree density in this stage was 1217 trees/ ha, while regeneration density was 3956 trees/ha (Table 1). The age structure in the canopy transition stage of stand development was similar to the stem exclusion/ca- nopy transition stage with a bimodal age structure as well (Figure 1(b)). The older cohort of canopy trees was com- Copyright © 2013 SciRes. OPEN A CCESS 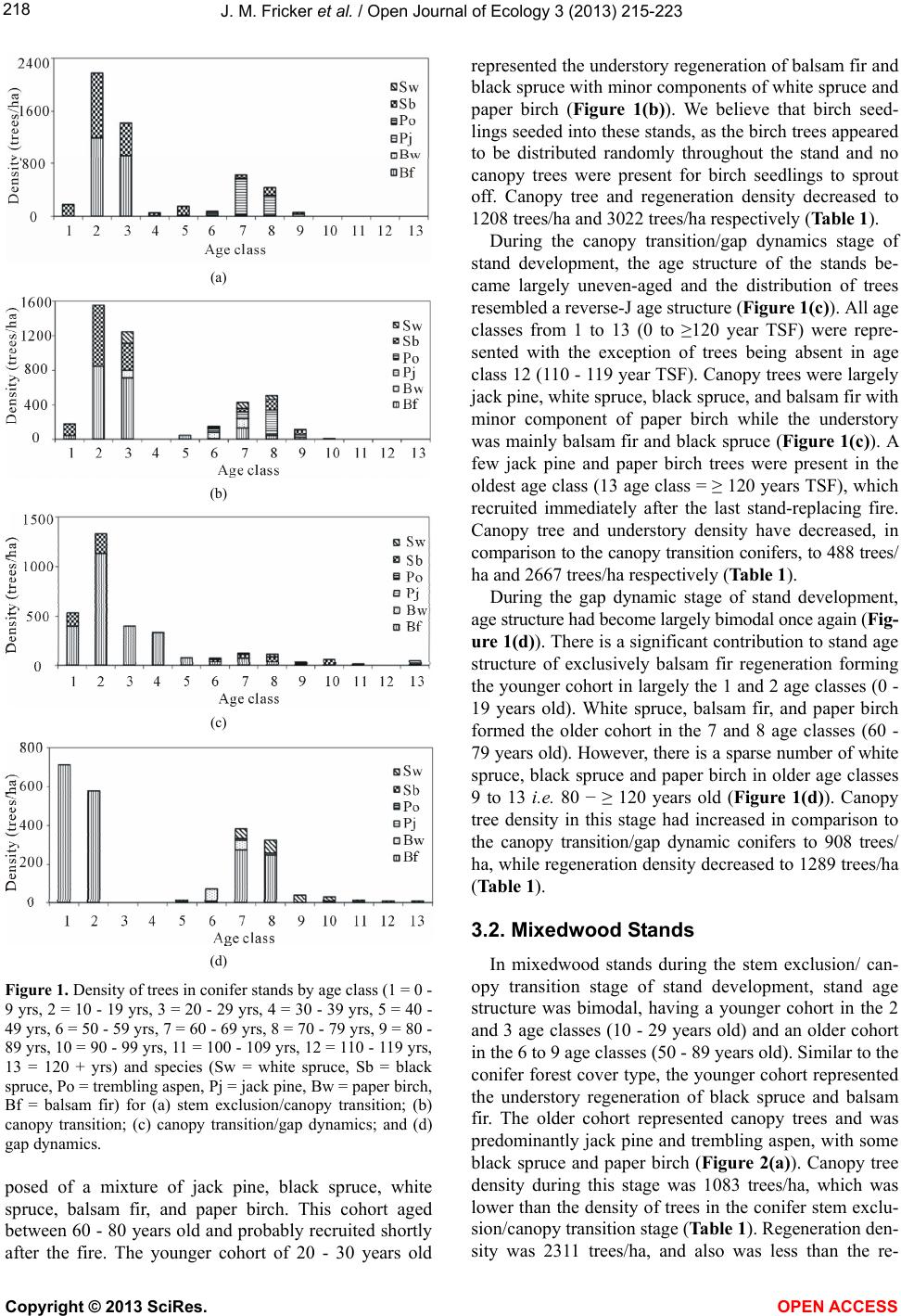 J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 2 15-223 218 (a) (b) (c) (d) Figure 1. Density of trees in conifer stands by age class (1 = 0 - 9 yrs, 2 = 10 - 19 yrs, 3 = 20 - 29 yrs, 4 = 30 - 39 yrs, 5 = 40 - 49 yrs, 6 = 50 - 59 yrs, 7 = 60 - 69 yrs, 8 = 70 - 79 yrs, 9 = 80 - 89 yrs, 10 = 90 - 99 yrs, 11 = 100 - 109 yrs, 12 = 110 - 119 yrs, 13 = 120 + yrs) and species (Sw = white spruce, Sb = black spruce, Po = trembling aspen, Pj = jack pine, Bw = paper birch, Bf = balsam fir) for (a) stem exclusion/canopy transition; (b) canopy transition; (c) canopy transition/gap dynamics; and (d) gap dynamics. posed of a mixture of jack pine, black spruce, white spruce, balsam fir, and paper birch. This cohort aged between 60 - 80 years old and probably recruited shortly after the fire. The younger cohort of 20 - 30 years old represented the understory regeneration of balsam fir and black spruce with minor components of white spruce and paper birch (Figure 1(b)). We believe that birch seed- lings seeded into these stands, as the birch trees appeared to be distributed randomly throughout the stand and no canopy trees were present for birch seedlings to sprout off. Canopy tree and regeneration density decreased to 1208 trees/ha and 3022 trees/ha respectively (Table 1). During the canopy transition/gap dynamics stage of stand development, the age structure of the stands be- came largely uneven-aged and the distribution of trees resembled a reverse-J age structure (Figure 1(c)). All age classes from 1 to 13 (0 to ≥120 year TSF) were repre- sented with the exception of trees being absent in age class 12 (110 - 119 year TSF). Canopy trees were largely jack pine, white spruce, black spruce, and balsam fir with minor component of paper birch while the understory was mainly balsam fir and black spruce (Figure 1(c)). A few jack pine and paper birch trees were present in the oldest age class (13 age class = ≥ 120 years TSF), which recruited immediately after the last stand-replacing fire. Canopy tree and understory density have decreased, in comparison to the canopy transition conifers, to 488 trees/ ha and 2667 trees/ha respectively (Table 1). During the gap dynamic stage of stand development, age structure had become largely bimodal once again (Fig- ure 1(d)). There is a significant contribution to stand age structure of exclusively balsam fir regeneration forming the younger cohort in largely the 1 and 2 age classes (0 - 19 years old). White spruce, balsam fir, and paper birch formed the older cohort in the 7 and 8 age classes (60 - 79 years old). However, there is a sparse number of white spruce, black spruce and paper birch in older age classes 9 to 13 i.e. 80 − ≥ 120 years old (Figure 1(d)). Canopy tree density in this stage had increased in comparison to the canopy transition/gap dynamic conifers to 908 trees/ ha, while regeneration density decreased to 1289 trees/ha (Table 1). 3.2. Mixedwood Stands In mixedwood stands during the stem exclusion/ can- opy transition stage of stand development, stand age structure was bimodal, having a younger cohort in the 2 and 3 age classes (10 - 29 years old) and an older cohort in the 6 to 9 age classes (50 - 89 years old). Similar to the conifer forest cover type, the younger cohort represented the understory regeneration of black spruce and balsam fir. The older cohort represented canopy trees and was predominantly jack pine and trembling aspen, with some black spruce and paper birch (Figure 2(a)). Canopy tree density during this stage was 1083 trees/ha, which was lower than the density of trees in the conifer stem exclu- sion/canopy transition stage (Table 1). Regeneration den- sity was 2311 trees/ha, and also was less than the re- Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 215-223 219 (a) (b) (c) Figure 2. Density of trees in mixedwood stands by age class (as in Figure 1) and species (Sw = white spruce, Sb = black spruce, Po = trembling aspen, Pj = jack pine, Bw = paper birch, Bf = balsam fir) for (a) stem exclusion/canopy transition; (b) canopy transition; and (c) gap dynamics. No data is available for the canopy transition/gap dynamic mixedwoods. generation occurring in the stem exclusion/canopy tran- sition conifers (Table 1). The age structure in the canopy transition stage of stand development was similar to the stem exclusion/ canopy transition stage (Figure 2(b)). It was character- ized by a strong bimodal age structure. A younger cohort represented balsam fir and black spruce regeneration with minor components of white spruce and paper birch that were in the 2 and 3 age classes (10 - 29 years old). The older cohort was composed of jack pine, trembling aspen, black spruce, white spruce, balsam fir, and paper birch trees that were within the 6 to 9 age classes i.e. 50 - 89 years old (Figure 2(b)). The density of canopy trees (1108 trees/ha) was higher than that occurring in the stem exclusion/canopy transition mixedwoods, which was low- er than that in the canopy transition conifers. As well, the regeneration density was lower (2222 trees/ha) compared to both the stem exclusion/canopy transition mixedwoods and the conifer canopy transitions (Table 1). We were unable to locate stands of the mixedwood co- ver type in the canopy transition/gap dynamics stage to sample. Therefore only three development stages for mixedwood cover types are presented. During the gap dynamic stage of stand development, age structure was somewhat bimodal (Figure 2(c)). There was a significant contribution to stand age structure of regeneration form- ing the younger cohort in the 1 and 2 age classes i.e. 0 - 29 years old. Canopy trees that formed the second small peak in the 7 and 8 age classes i.e. 60 - 79 years old were a mixture of paper birch, balsam fir, and white and black spruce (Figure 2(c)). The understory regeneration was pre- dominantly balsam fir with a small component of white spruce. Canopy tree density has decreased in comparison to canopy transition mixedwoods to 1067 trees/ha, which was higher than that in the gap dynamic conifers (Table 1). Regeneration was 3778 trees/ha, higher than the den- sity of regeneration in both the canopy transition mixed- woods and the gap dynamic conifers. In general, when present in the stands, balsam fir, white spruce and black spruce were found in most age classes. 3.3. Hardwood Stands During the stem exclusion/canopy transition stage of stand development in the hardwoods, stand age structure was typical bimodal, having a younger cohort in the 1, 2 and 3 age classes (0 - 30 years old) and an older cohort in the 6 to 9 age classes (50 - 90 years old). The older co- hort was composed of largely trembling aspen and black spruce, with some paper birch and balsam fir, while the younger cohort was the understory regeneration of black spruce and balsam fir (Figure 3(a)). Canopy tree density was 1158 trees/ha, and was similar to the canopy tree den- sity in the stem exclusion/canopy transition mixedwoods but lower than the conifers of this stage (Table 1). Rege- neration density was 1111 trees/ha, and was lower than both the conifer and mixedwood stem exclusion/canopy transition stands. Unlike the age structure in conifer and mixedwood co- ver types, the age structure in the canopy transition hard- woods was weakly unimodal. The major canopy tree co- hort was composed of largely trembling aspen within the 7 to 10 age classes (60 - 100 years old) regenerated shortly after the fire (Figure 3(b)). Understory regenera- tion density was very low (178 trees/ha) and spread across the 1 to 4 age classes (Table 1, Figure 3(b)). Canopy tree density was 675 trees/ha lower than the density of cano- py trees in the stem exclusion/canopy transition hardwoods and the canopy transition conifers and mixedwoods. During the canopy transition/gap dynamics stage of stand development, the age structure of the stand appears Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 2 15-223 220 (a) (b) (c) (d) Figure 3. Density of trees in hardwood stands by age class (as in Figure 1) and species (Sw = white spruce, Sb = black spruce, Po = trembling aspen, Pj = jack pine, Bw = paper birch, Bf = balsam fir) for (a) stem exclusion/canopy transition; (b) canopy transition; (c) canopy transition/gap dynamics; and (d) gap dy- namics. to be weakly reverse-J to weakly bimodal, with majority of trees in the younger cohort in the 2 and 3 age classes (10 - 20 years old). There were trees in every age class from 1 to 13 (0 to ≥120 years old) (Figure 3(c)). The older canopy trees were mainly trembling aspen and black spruce, while the younger canopy trees were large- ly balsam fir with some white and black spruce, paper birch, and trembling aspen. Regeneration cohort was lar- gely balsam fir and black spruce (Figure 3(c)). Canopy tree density increased compared to the canopy transition hardwoods to 717 trees/ha, while being higher than their conifer counterparts (Table 1). Regeneration density also increased compared to the canopy transition hardwoods to 2089 trees/ha, but was lower than their conifer coun- terparts. Age structure has become largely bimodal once again for the gap dynamic stage of hardwood stand develop- ment. There was a significant understory cohort of rege- neration dominated by white spruce and balsam fir and paper birch in the 1 to 3 age classes (0 - 30 years old). Canopy trees were largely paper birch with minor com- ponents of balsam fir, white spruce, and trembling aspen forming the older cohort in the 6 to 8 age classes (50 - 79 years old) (Figure 3(d)). Canopy tree and regeneration density both increased (1025 trees/ha and 2867 trees/ha, respectively) compared to the canopy transition/gap dy- namics hardwoods (Ta bl e 1 ). Canopy tree and regenera- tion density were both lower than their mixedwood but higher than their conifer respective counterparts. 4. DISCUSSION After Large, mature white birch were dispersed throu- ghout the hardwood stands comprising less than 1% of the total stem density and 5% of the stem basal area. Even though white birch is traditionally considered an “early successional” species, this species can persist wi- thin old-growth boreal coniferous stands [9]. Its prolific seed-producing ability, combined with the availability of suitable microsites within sufficiently larger gaps, proba- bly maintains the presence of white birch. Different stand types should also be managed differ- ently. In hardwood stands, clear-cut logging appears to emulate natural disturbance processes quite well, since aspen and white birch successfully sprout back immedi- ately following both fire and logging. In contrast, clear- cut harvesting of jack pine and black spruce is more pro- blematic, since the seed source is largely removed espe- cially if whole-tree harvesting is followed by roadside chipping. Stand age structural development in conifer stands pro- ceeded from a bimodal structure in the stem exclusion/ canopy transition and canopy transition stages to a re- verse-J age structure in the canopy transition/gap dyna- mics stage to a bimodal structure once again in the gap dynamics stage. In the stem exclusion/canopy transition and canopy transition stages, the canopy was dominated by jack pine established immediately after the stand-re- placing fire, as significant age-related mortality yet oc- curred. In turn, self-thinning that occurred in earlier stages of development would have opened up growing space Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 215-223 221 and freed up nutrients [5,27], thus contributing to the sig- nificant regeneration of conifers such as black spruce and balsam fir. By the canopy transition/gap dynamics stage of stand development, most of the pioneering cohort had died off, as only a few jack pine trees remain living in the stands (average 42 trees per hectare) because the age of these stands (139 years TSF) is beyond jack pine’s average life span [28]. As well, trees that were suppressed in earlier stages of stand development were released to undergo ra- pid growth and moved into the canopy and subcanopy positions. Mortality of the pioneering cohort of jack pine would free up additional space for further understory re- generation to establish. This resulted in a reverse-J age structure in these stands, as observed by [29], who found that old (>120 year-old) spruce stands in sub-boreal British Columbia show either a reverse-J or bimodal age structure. During the gap dynamic stage of stand development, pioneering cohort of jack pine had completely died off, and the canopy was dominated by late successional coni- fers. These conifers were younger than the jack pine that dominated in the preceding stage, causing the oldest age classes to disappear. As such, the age structure became bimodal, with a canopy dominated by later successional species and an understory with young regeneration of co- nifers, largely balsam fir. The very low density of balsam fir in age classes 3 to 6 (20 - 59 years) may have been caused by periodic spruce budworm (Choristoneura fu- miferana (Clem.)) outbreaks. The most recent one peak- ed in 1986 and had collapsed approximately ten years la- ter [30]. These outbreaks would have killed a significant amount of host-specific balsam fir and, to a lesser extent, white and black spruce trees [31-34]. While differing from conifer stands, hardwood stands were dominated by trembling aspen, hardwood stands in the stem exclusion/canopy transition stage of stand deve- lopment with a bimodal age structure with a small black and white spruce component established shortly after the stand-replacing fire. Understory contained largely conifer regeneration dominated by balsam fir. However, hard- woods in the canopy transition stage had a unimodal stand age structure with largely a single cohort of trem- bling aspen trees established after the disturbance and marginal amounts of conifer regeneration. Stands domi- nated by trembling aspen often developed a dense shrub layer of mountain maple (Acer spicatum) [35] and beak- ed hazel (Corylus cornuta) [36]. This was observed in all sampled hardwood stands in the canopy transition stage. Furthermore, dense shrub layers have been found to hin- der understory conifer regeneration [35-37], thus contri- buting to the unimodal age structure in the canopy transi- tion hardwoods. In contrast to the conifer stands, hardwood stands had many more trembling aspen trees (200 trees·ha−1) from the pioneering cohort still living in the canopy transi- tion/gap dynamic stage of stand development even thou- gh jack pine is generally a longer-lived tree species com- pared to trembling aspen [28]. This may be due to the hardwood sites in the canopy transition/gap dynamic stage of stand development being more productive than the conifer sites that were sampled. However, as in the coni- fer stands, hardwoods had a relatively reverse-J stand age structure. Paper birch dominated the hardwoods in the gap dy- namic stage of development. Paper birch has been shown to be able to live for well over 200 years [9], and is the only hardwood species in this area of the boreal forest that could form dominant stands by this stand age. Fur- ther, the ability of paper birch to allow light to pass through to the forest floor, and the sparse shrub layer that was found, would allow for a significant amount of un- derstory regeneration to establish. Therefore, the age struc- ture of gap dynamic hardwoods were similar to that of the gap dynamic conifers with a bimodal age stand struc- ture caused by a canopy of paper birch trees falling into the older age classes and regeneration of conifers form- ing the younger age classes. With the exception of mixedwood stands having cano- pies composed of a mixture of conifers and hardwoods that met the sampling criteria (25% - 75% conifer com- ponent), stand age structure of mixedwood stands devel- oped similarly to that of conifer stands. This was caused by the conifer component limiting light to the forest floor and preventing a dense shrub layer from developing [35]. Without this dense shrub layer, regeneration would have responded similarly to what was occurring in conifer stands, thus causing a similar age structure to develop. We believe that successional trajectories in mixedwood stands may be headed towards conifer dominance due to the composition, as indicated by the composition of rege- neration. However, budworm outbreaks tend to occur about every 20 years [30], therefore making this uncertain. Regardless of stand cover type or developmental stage, regeneration in all the stands was almost exclusively bal- sam fir and spruce, indicating that successional trajecto- ries in the study area are likely proceeding towards coni- fer dominance on most sites. This is likely a consequence of the silvics of these species and the availability of near- by seed sources. Black spruce is moderately shade-tole- rant, allowing it to establish under the cover of other trees, while also being able to reproduce by layering [38]. Bal- sam fir is a shade-tolerant species, and has seeds that are readily dispersed by wind [39] and enter a stand from nearby areas. 5. IMPLICATION FOR OLD-GROWTH MANAGEMENT Old-growth forests have been found to provide many Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 2 15-223 222 values from an ecological, aesthetic/recreational, and eco- nomic perspective [8,40]. However, management deci- sions surrounding old-growth are hampered by the lack of a clear definition on what old-growth is in the boreal forest [40,41]. While some studies including this study use the disappearance of the pioneering cohort as the point at which an old-growth structure is reached [1,8] defini- tions of old-growth vary depending on the study [42]. We recommend that old-growth in this region of the boreal forest be considered when the following criteria are met: 1) canopy breakdown of the pioneering cohort is com- plete and the stand is dominated by later successional tree species such as balsam fir and spruce, and 2) the age structure of the stand is bimodal, with dominating can- opy trees that fall within a relatively narrow range of age classes and a significant amount of understory regenera- tion. Selection harvesting could be used to hasten the onset of old-growth and/or create a reverse-J stand age struc- ture if applied to stands that are in approximately the stem exclusion/canopy transition stage of stand develop- ment or even earlier in the stem exclusion stage. We sug- gest that selectively removing canopy trees would (a) re- lease suppressed trees, (b) allow canopy trees to grow even faster, and (c) allow trees to establish in gaps cre- ated by the removal of canopy trees. This would promote the movement of a unimodal or bimodal age structure into a reverse-J age structure while increasing the later successional species in the stand thereby hastening old- growth onset. Sustained efforts must be made to ensure that harvest- ing rotations are such that various age and size structure of aspen stands are maintained at the landscape level. By age 100, jack pine stands in this area consist of an uneven- aged mixture of many species, making them less desir- able for logging. The harvest of large patches of aspen in the complex canopy matrix could help perpetuate the hard- wood component and thereby promote landscape-level biodiversity. 6. ACKNOWLEDGEMENTS The work was supported financially by the Natural Sciences and En- gineering Research Council (238891-01) and the Sustainable Forest Management Network of Centers of Excellence. We thank the review- ers for their constructive comments. REFERENCES [1] Chen, H.Y.H. and Popadiouk, R.V. (2002) Dynamics of North American boreal mixedwoods. Environmental Re- views, 10, 137-166. doi:10.1139/a02-007 [2] Taylor, A.R. and Chen, H.Y.H. (2011) Multiple success- sional pathways of boreal forest stands in central Canada. Ecography, 34, 208-219. doi:10.1111/j.1466-8238.2011.00689.x [3] Chen, H.Y.H. and Taylor, A.R. (2012) A test of ecological succession hypotheses using 55-year time-series data for 361 boreal forest stands. Global Ecology and Biogeog- raphy, 21, 441-454. doi:10.1111/j.1466-8238.2011.00689.x [4] Daniels, L.D., Marshall, P.L., Carter, R.E. and Klinka, K. (1995) Age structure of Thuja plicata in the tree layer of old-growth stands near Vancouver, British Columbia. Northern Science, 69, 175-183. [5] Oliver, C.D. and Larson, B.C. (1996) Forest stand dyna- mics. John Wiley & Sons, Inc., New York. [6] Smith, D.V., Larson, B.C., Kelty, M.J. and Ashton, P.M.S. (1997) The practice of silviculture: Applied forest ecology. 9th Edition, John Wiley & Sons, Inc., New York. [7] Pothier, D., Raulier, F. and Riopel, M. (2004) Ageing and decline of trembling aspen stands in Quebec. Canadian Journal of Forest Research, 34, 1251-1258. doi:10.1139/x04-017 [8] Brassard, B.W. and Chen, H.Y.H. (2006) Stand structural dynamics of North American boreal forests. Critical Re- views in Plant Sciences 25, 115-137. doi:10.1080/07352680500348857 [9] Bergeron, Y. (2000) Species and stand dynamics in the mixedwoods of Quebec’s southern boreal forest. Ecology, 81, 1500-1516. doi:10.1890/0012-9658(2000)081[1500:SASDIT]2.0.CO; 2 [10] Des Rochers, A. and Gagnon, R. (1997) Is ring count at ground level a good estimation of black spruce age? Ca- nadian Journal of Fore st Research, 27, 1263-1267. doi:10.1139/x97-086 [11] Sano, J. (1997) Age and size distribution in a long-term forest dynamics. For. Ecol. Manage, 92, 39-44. doi:10.1016/S0378-1127(96)03958-8 [12] Lee, P., Hanus, S. and Grover, B. (2000) Criteria for es- timating old growth in boreal mixedwoods from standard timber inventory data. Forest Ecology and Management, 129, 25-30. doi:10.1016/S0378-1127(99)00165-6 [13] Hély, C., Bergeron, Y. and Flannigan, M.D. (2000) Ef- fects of stand composition on fire hazard in mixed-wood Canadian boreal forest. Journal of Vegetable Science, 11 , 813-824. doi:10.2307/3236551 [14] MacPherson, D.M., Lieffers, V.J. and Blenis, P.V. (2001) Productivity of aspen stands with and without spruce un- derstory in Alberta’s boreal mixedwood forests. The F or- estry Chronicle, 77, 351-356. [15] Pedlar, J.H., Pearce, J.L., Venier, L.A. and McKenney, D.W. (2002) Coarse woody debris in relation to distur- bance and forest type in boreal Canada. Forest Ecology and Management, 158, 189-194. doi:10.1016/S0378-1127(00)00711-8 [16] Légaré, S., Bergeron, Y. and Paré, D. (2005) Effect of as- pen (Populus tremuloides) as a companion species on the growth of black spruce (Picea mariana) in the southwe- stern boreal forest of Quebec. Forest Ecology and Man- agement, 208, 211-222. doi:10.1016/j.foreco.2004.12.004 [17] Johnson, E.A., Miyanishi, K. and Weir, J.M.H. (1995) Old- Copyright © 2013 SciRes. OPEN A CCESS  J. M. Fricker et al. / Open Journal of Ecology 3 (2013) 215-223 Copyright © 2013 SciRes. OPEN A CCESS 223 growth, disturbance, and ecosystem management. Cana- dian Journal of Botany, 73, 918-926. doi:10.1139/b95-100 [18] OMNR (2003) Nonlinear height-diameter models for nine boreal forest tree species in Ontario. [19] Rowe, J.S. (1972) Forest regions of Canada. Canadian Forest Servics, Ottawa, Publications, 1300. [20] Environment Canada (2005) Climate normals for Thun- der Bay, ON, Canada (1971-2000). http://www.climate.weatheroffice.ec.gc.ca/climate_norma ls [21] Senici, D., Chen, H.Y.H., Bergeron, Y. and Cyr, D. (2010) Spatiotemporal variations of fire frequency in central bo- real forest. Ecosystems, 13, 1227-1238. doi:10.1007/s10021-010-9383-9 [22] Greif, G.E. and Archibold, O.W. (2000) Standing-dead tree component of the boreal forest in central Saskatchewan. Forest Ecology and Management, 131, 37-46. doi:10.1016/S0378-1127(99)00198-X [23] British Columbia Ministry of Environment, L.A.P. and British Columbia Ministry of Forests (1998) Field manual for describing terrestrial ecosystems. Land Manage, 25. [24] Sims, R.A., Towill, W.D., Baldwin, K.A., Uhlig, P. and Wickware, G.M. (1997) Field guide to the forest ecosys- tem classification for northwestern Ontario. Queen’s Prin- ter for Ontario, Toronto. [25] Soil Classification Working Group (1998) The Canadian system of soil classification. Agriculture of Canadian Pub- lications, 1646. [26] Vasiliauskas, S. and Chen, H.Y.H. (2002) How long do trees take to reach breast height after fire in northeastern Ontario? Canadian Journal of Forest Research, 32, 1889- 1892. doi:10.1139/x02-104 [27] Chen, H.Y.H., Klinka, K. and Kayahara, G.J. (1996) Ef- fects of light on growth, crown architecture, and specific leaf area for naturally established Pinus contorta var la- tifolia and Pseudotsuga menziesii var glauca saplings. Canadian Journal of Forest Research, 26, 1149-1157. doi:10.1139/x26-128 [28] Farrar, J.L. (1995) Trees in Canada. Fitzhenry & White- side Ltd. and the Canadian Forest Servics, Toronto. [29] Kneeshaw, D.D. and Burton, P.J. (1997) Canopy and age structures of some old sub-boreal Picea stands in British Columbia. Journal of Vegetable Science, 8, 615-626. doi:10.2307/3237365 [30] Fleming, R.A., Hopkin, A.A. and Candau, J.N. (2000) In- sect and Disease Disturbance Regimes in Ontario’s For- ests. In: Perera, A.H., Euler, D.L. and Thompson, I.D., Eds., Ecology of a Managed Terrestrial Landscape: Pat- terns and Processes in Forest Landscape of Ontario. Uni- versity of British Columbia Press, Vancouver, 141-162. [31] MacLean, D.A. and Ostaff, D.P. (1989) Patterns of bal- sam fir mortality caused by an uncontrolled spruce bud- worm outbreak. Canadian Journal of Forest Research, 19, 1087-1095. doi:10.1139/x89-165 [32] Bergeron, Y., Leduc, A., Morin, H. and Joyal, C. (1995) Balsam fir mortality following the last spruce budworm outbreak in northwestern Quebec. Canadian Journal of Fore st Research, 25, 1375-1384. doi:10.1139/x95-150 [33] Parent, S., Morin, H. and Messier, C.( 2001) Balsam fir (Abies balsamea) establishment dynamics during a spruce budworm (Choristoneura fumiferana) outbreak: an evalu- ation of the impact of aging techniques. Canadian Jour- nal of Forest Research, 31, 373-376. [34] Burleigh, J.S., Alfaro, R.I., Borden, J.H. and Taylor, S. (2002) Historical and spatial characteristics of spruce bud- worm Choristoneura fumiferana (Clem.) (Lepidoptera: Tortricidae) outbreaks in northeastern British Columbia. Forest Ecology and Management, 168, 301-309. doi:10.1016/S0378-1127(01)00748-4 [35] Bourgeois, L., Messier, C. and Brais, S. (2004) Mountain maple and balsam fir early response to partial and clear- cut harvesting under aspen stands of northern Quebec. Canadian Journal of Forest Research, 34, 2049-2059. doi:10.1139/x04-080 [36] Hill, S.B., Mallik, A.U. and Chen, H.Y.H. (2005) Canopy gap disturbance and succession in trembling aspen domi- nated boreal forests in northeastern Ontario. Canadian Journal of Forest Research, 35, 1942-1951. doi:10.1139/x05-126 [37] Wallenius, T., Kuuluvainen, T., Heikkilä, R. and Lindholm, T. (2002) Spatial tree age structure and fire history in two old-growth forests in eastern Fennoscandia. Silva Fenn, 36, 185-199. [38] Charron, I. and Greene, D.F. (2002) Post-wildfire seed- beds and tree establishment in the southern mixedwood boreal forest. Canadian Journal of Forest Research, 32, 1607-1615. doi:10.1139/x02-085 [39] Wang, G.G. and Kemball, K.J. (2005) Balsam fir and white spruce seedling recruitment in response to understory re- lease, seedbed type, and litter exclusion in trembling as- pen stands. Canadian Journal of Forest Research, 35, 667-673. doi:10.1139/x04-212 [40] Cogbill, C.V. (1984) Dynamics of the boreal forests of the Laurentian Highlands, Canada. Canadian Journal of Fo- rest Research, 15, 252-261. doi:10.1139/x85-043 [41] Barnard, E. (2004) Old-growth: some questions, truths, and consequences. Journal of Forest , 102, 60. [42] Franklin, J.F., Spies, T.A., Van Pelt, R., Carey, A.B., Thorn- burgh, D.A., Berg, D.R., Lindenmayer, D.B., Harmon, M.E., Keeton, W.S., Shaw, D.C., Bible, K. and Chen, J. (2002) Disturbances and structural development of natu- ral forest ecosystems with silvicultural implications, us- ing Douglas-fir forests as an example. Forest Ecology and Management, 155, 399-423. doi:10.1016/S0378-1127(01)00575-8
|