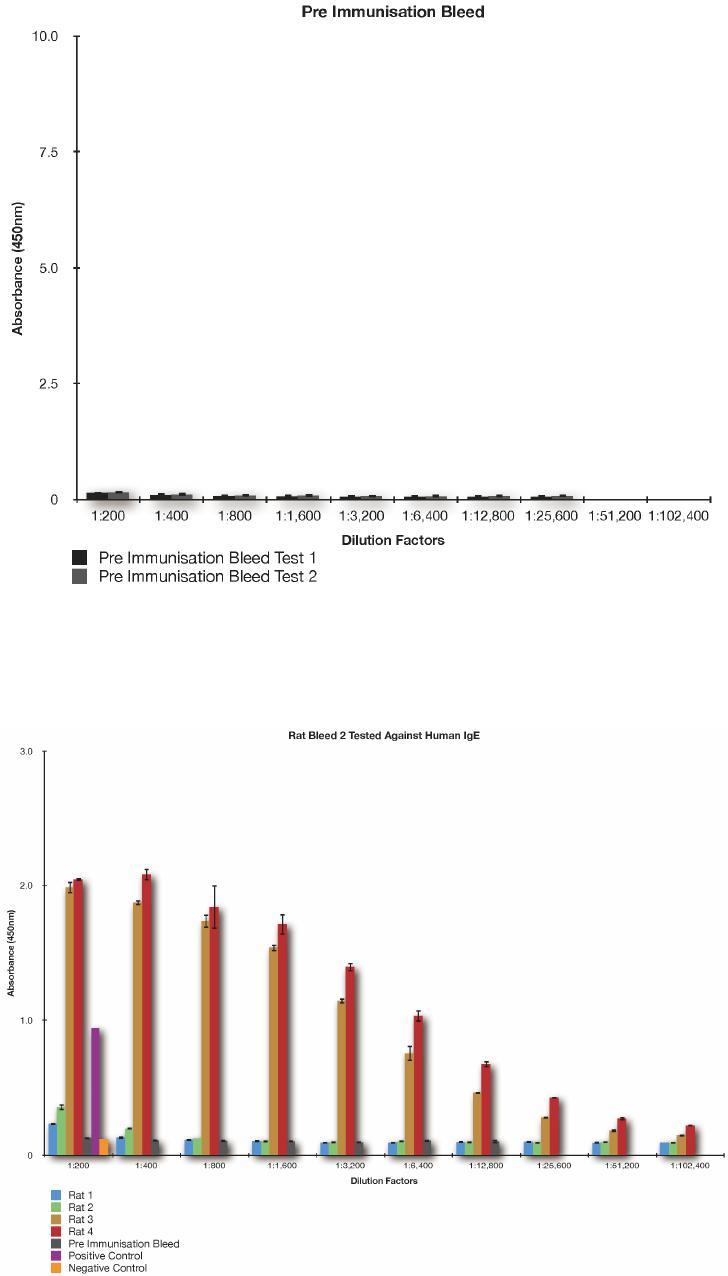
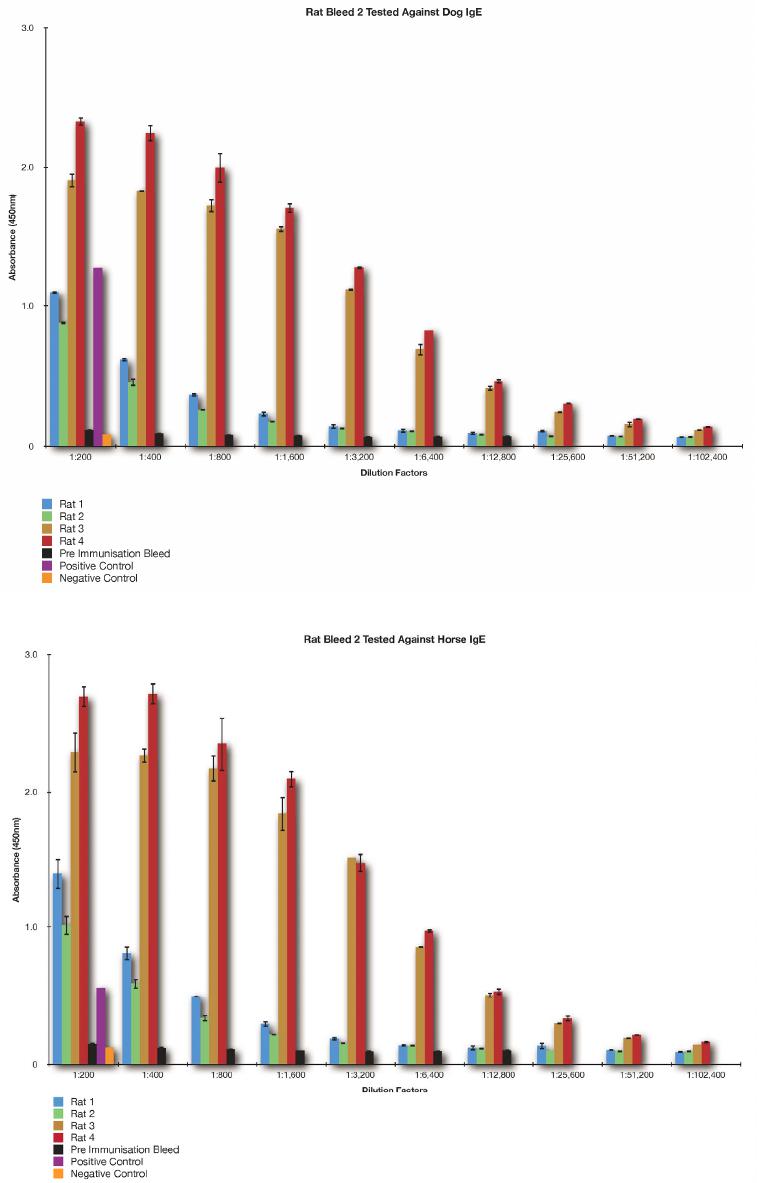
S. S. SABBAN ET AL.
26
Protein of Immunoglobulin Class E,” Lancet, Vol. 2, No.
7558, 1968, pp. 17-18.
doi:10.1016/S0140-6736(68)92889-4
[3] P. J. Barnes, “Therapeutic Strategies for Allergic Dis-
eases,” Nature, Vol. 402, Suppl. B, 1999, pp. 31-38.
[4] B. A. Helm, A. C. Spivey and E. A. Padlan, “Peptide
Blocking of IgE/Receptor Interaction: Possibilities and
Pitfalls,” Allergy, 1997, Vol. 52, No. 12, pp. 1155-1169.
doi:10.1111/j.1398-9995.1997.tb02518.x
[5] W. A. Hook, et al., “Monoclonal Antibodies to Human
IgE,” Federation Proceedings, Vol. 40, 1981.
[6] L. G. Presta, S. J. Lahr, R. L. Shields, J. P. Porter, C. M.
Gorman, B. M. Fendly and P. M. Jardieu, “Humanization
of an Antibody Directed against IgE,” Journal of Immu-
nology, Vol. 151, No. 5, 1993, pp. 2623-2632.
[7] S. Laffer, E. Hogbom, K. H. Roux, W. R. Sperr, P. Valent,
H. C. Bank, L. Vangelista, F. Kricek, D. Kraft, H. Grön-
lund and R. Valenta, “A Molecular Model of Type I Al-
lergy: Identification and Characterization of a Nonana-
phylactic Anti-Human IgE Antibody Fragment that Blocks
the IgE-FcεRI Interaction and Reacts with Receptor-
Bound IgE,” The Journal of Allergy and Clinical Immu-
nology, Vol. 108, No. 3, 2001, pp. 409-416.
doi:10.1067/mai.2001.117593
[8] S. Laffer, C. Lupinek, I. Rauter, M. Kneidinger, A. Dre-
scher, J. H. Jordan, M. T. Krauth, P. Valent, F. Kricek, S.
Spitzauer, H. Englund and R. Valenta, “A High-Affinity
Monoclonal Anti-IgE Antibody for Depletion of IgE and
IgE-Bearing Cells,” Allergy, Vol. 63, No. 6, 2008, pp.
695-702. doi:10.1111/j.1398-9995.2008.01664.x
doi:10.1038/35018500
[9] S. C. Garman, B. A. Wurzburg, S. S. Tarchevskaya, J.-P.
Kinet and T. S. Jardetzky, “Structure of the Fc Fragment
of Human IgE Bound to Its High-Affinity Receptor
FcεRIα,” Nature, Vol. 406, 2000, pp. 259-266.
[10] A. W. Wheeler, J. S. Marshall and J. T. Ulrich, “A Th-1
Inducing Adjuvant, MPL, Enhances Antibody Profiles in
Experimental Animals Suggesting It Has the Potential to
Improve the Efficacy of Allergy Vaccines,” International
Archives of Allergy and Immunology, Vol. 126, No. 2,
2001, pp. 135-139. doi:10.1159/000049504
[11] C. Y. Wang, A. M. Walfield, X. Fang, B. Hammerberg, J.
Ye, M. L. Li, F. Shen, M. Shen, V. Alexander and D. W.
MacGlashan, “Synthetic IgE Peptide Vaccine for Immu-
notherapy of Allergy,” Vaccine, Vol. 21, No. 15, 2003, pp.
1580-1590. doi:10.1016/S0264-410X(02)00732-6
[12] F. R. Ali and M. Larché, “Peptide-Based Immunotherapy:
A Novel Strategy for Allergic Disease,” Expert Review of
Vaccines, Vol. 4, No. 6, 2005, pp. 881-889.
doi:10.1586/14760584.4.6.881
[13] A. Ledin, K. Bergvall, N. S. Hillbertz, H. Hansson, G.
Andersson, A. Hedhammar and L. Hellman, “Generation
of Therapeutic Antibody Responses against IgE in Dogs,
an Animal Species with Exceptionally High Plasma IgE
levels,” Vaccine, Vol. 24, No. 1, 2006, pp. 66-74.
doi:10.1016/j.vaccine.2005.07.052
[14] V. Niederberger and R. Valenta, “Molecular Approaches
for New Vaccines against Allergy,” Expert Review of
Vaccines, Vol. 5, No. 1, 2006, pp. 103-110.
doi:10.1586/14760584.5.1.103
[15] Z. Peng, Q. Liu, Q. Wang, E. Rector, Y. Ma and R. War-
rington, “Novel IgE Peptide-Based Vaccine Prevents the
Increase of IgE and Down-Regulates Elevated IgE in
Rodents,” Clinical & Experimental Allergy, Vol. 37, No.
7, 2007, pp. 1040-1048.
doi:10.1111/j.1365-2222.2007.02741.x
[16] J. Johansson and L. Hellman, “Modifications Increasing
the Efficacy of Recombinant Vaccines; Marked Increase
in Antibody Titers with Moderately Repetitive Variants
of a Therapeutic Allergy Vaccine,” Vaccine, Vol. 25, No.
9, 2007, pp. 1676-1682.
doi:10.1016/j.vaccine.2006.10.055
[17] L. F. Pacios, L. Tordesillas, J. Cuesta-Herranz, E. Com-
pes, R. Sánchez-Monge, A. Palacín, G. Salcedo and A.
Díaz-Perales, “Mimotope Mapping as a Complementary
Strategy to Define Allergen IgE-Epitopes: Peach Pru p 3
Allergen as a Model,” Molecular Immunology, Vol. 45,
No. 8, 2008, pp. 2269-2276.
doi:10.1016/j.molimm.2007.11.022
[18] L. T. Hellman, S. Persson and A. Jansson, “Allergy,”
Vaccines US Patent No. 2009/0191268 A1, 2009.
[19] T. Francis Jr., “The Current Status of the Control of In-
fluenza,” Annals of Internal Medicine, Vol. 43, No. 3,
1955, pp. 534-538. doi:10.7326/0003-4819-43-3-534
[20] T. Francis Jr., “On the Doctrine of Original Antigenic
Sin,” Proceedings of the American Philosophical Society,
Vol. 104, No. 6, 1960, pp. 572-578.
[21] A. P. M. Wilson, C. E. Pullar, A. M. Cmp and B. A.
Helm, “Human IgE Mediates Stimulus Secretion Cou-
pp. 240-244. doi:10.1002/eji.1830230137
a
pling in Rat Basophilic Leukemia Cells Transfected with
the Alpha Chain of the Human High-Affinity Receptor,”
European Journal of Immunology, Vol. 23, No. 1, 1993,
[22] J. M. Hunter, A. P. Vratimos, J. E. Housden and B. A.
ion of the Canine Domain
Helm, “Generation of Canine-Human Fc IgE Chimeric
Antibodies for the Determinat
of Interaction with Fc Epsilon RI Alpha,” Molecular Im-
munology, Vol. 45, No. 8, 2008, pp. 2262-2268.
doi:10.1016/j.molimm.2007.11.015
[23] S. Sabban, “Development of an in Vitro Model System
for Studying the Interaction of Equus caballus IgE with
Its High-Affinity FcεRI Receptor,” Ph.D. Thesis, The
University of Sheffield, Sheffield, 2011,
http://etheses.whiterose.ac.uk/2040/2/Sabban%2C_Sari.p
1-2, 2013, pp. 6-10. doi:10.1016/j.vetimm.2013.01.008
df
[24] S. Sabban, H. Ye and B. A. Helm, “Development of an in
Vitro Model System for Studying the Interaction of Equus
caballus IgE with Its High-Affinity Receptor FcεRI,” Vet-
erinary Immunology and Immunopathology, Vol. 153, No.
[25] M. S. Neuberger, G. T. Williams, E. B. Mitchell, S. S.
Jouhal, J. G. bbitts, “A Hapten-
Specific Chimth Human Physio-
Flanagan and T. H. Ra
aeric IgE Antibody wi
logical Effector Function,” Nature, Vol. 314, No. 6008,
1985, pp. 268-270. doi:10.1038/314268a0
[26] S. L. Morrison, M. J. Johnson, L. A. Herzenberg and V. T.
Oi, “Chimeric Human Antibody Molecules: Mouse Anti-
gen-Binding Domains with Human Constant Region Do-
Copyright © 2013 SciRes. JIBTVA