Advances in Bioscience and Biotechnology
Vol.08 No.01(2017), Article ID:73635,24 pages
10.4236/abb.2017.81003
Hepatoprotective Activity of Chitosan Nanocarriers Loaded with the Ethyl Acetate Extract of a Stenotrophomonas sp. Bacteria Associated with the Red Sea Sponge Amphimedon ochracea in CCl4 Induced Hepatotoxicty in Rats
Nihad M. Abdel Monein1, Galila A. Yacout1, Haiam M. Aboul-Ela2, Mohamed A. Shreadah2*
1Biochemistry Department, Faculty of Science, Alexandria University, Alexandria, Egypt
2Marine Biotechnology and Natural Products Lab., National Institute of Oceanography and Fisheries, Alexandria, Egypt

Copyright © 2017 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: December 15, 2016; Accepted: January 17, 2017; Published: January 20, 2017
ABSTRACT
The present study investigated the protective role of chitosannano-carriers loaded with the ethyl acetate extract of a Stenotrophomonas sp. bacteria associated with the Red Sea Sponge Amphimedon ochracea CCl4 induced hepatic damage in rats using the hepatological parameters. Healthy 80 male Sprague- Dawley rats weighed 50 - 80 gm were used. The animals were randomly divided into eight groups. Serum ALT and AST, albumin, direct buluibin and histopathological examinations were investigated. Liver targeting of the drugs has been achieved after administration of drug loaded spherical particles. This mode of administration of drugs enhanced its overall delivery to the liver via passive targeting. The present study revealed that the treatment with ethyl acetate bacterial extract (10 mg/Kg body weight) protected rats from hepatotoxic action of CCl4. Usage of chitosan nanoparticles loaded with the bacterial extract as a drug delivery system has greatly improved the curative effect of bacterial extract. Moreover, histopathological studies showed marked reduction in fatty degeneration and centrizonal necrosis in animals receiving the nano-chitosan loaded with the bacterial extract along with ethanol compared to the control group.
Keywords:
Hepatoprotective Activity, Chitosan Nanocarriers, Extract of Stenotrophomonas sp. Bacteria, Red Sea Sponge

1. Introduction
Liver is one of the largest organs in human body and the chief site for intense metabolism and excretion. So it has a surprising role in the maintenance, performance and regulating homeostasis of the body. It is involved with almost all the biochemical pathways to growth, fight against disease, nutrient supply, energy provision and folk reproduction [1] . The major functions of the liver are carbohydrate, protein and fat metabolism, detoxification, secretion of bile and storage of vitamin. A healthy liver is a crucial factor for overall health and well- being. But it is continuously and variedly exposed to environmental toxins and abused by poor drug habits, as well as alcohol, prescribed and over-the-counter drug which can eventually lead to various liver ailments like hepatitis, cirrhosis and alcoholic liver disease [2] [3] . Thus, liver diseases are some of the fatal diseases in the world today. Marine sponges for the past decades have been considered as a very fertile field for the discovery of bioactive natural chemical substances with respect to the diversity of their primary and secondary chemical components and metabolites. As a significant portion of the bioactive metabolites, thought originally to be products of the source animal often synthesized by their symbiotic microbiota, researchers have paid more attention to microorganisms as a renewable source of bioactive natural products. More specifically, bacteria from the marine environment have shown great potential as suggested by the diversity of secondary metabolites [4] [5] . In contrast to the terrestrial environment where plants are the most prolific sources of natural products, this leading position in the sea is taken by invertebrates such as sponges, tunicates, bryozoans, or mollusks, with sponges being the most prolific sources of natural products [6] . The wealth of their secondary metabolites has been related to the role of chemical defense played by these constituents; such ecological function is believed to be crucial for the survival of the producer organisms, which are soft-bodied, sessile or slow-moving animals, lacking, in most cases, morphological defense structures such as shells or spines [7] . Sponges have been the focus of much recent interest due to the following two main (and often interrelated) factors: (i) they form close associations with a wide variety of microorganisms, and (ii) they are a rich source of biologically active secondary metabolites. This increasing research interest has greatly improved our knowledge of sponge- microbe interactions [7] . With the development of the phylogenetic analysis of 16S rRNA gene sequences, putative identification of bacteria by comparison with sequences from known closely related organisms can be easily achieved. The efficiency of many drugs is often limited by their potential to reach the site of therapeutic action. In most cases, only a small amount of administered dose reaches the target site, while the majority of the drug distributes throughout the rest of the body in accordance with its physicochemical and biological properties. Therefore, developing a drug delivery system that optimizes the pharmaceutical action of drug and reducing its toxic side effects in vivo is a challenging risk. One of the approaches is the use of colloidal drug carriers that can provide site specific or targeted drug delivery combined with optimal drug release profiles. Among these carriers, liposomes Cand nanoparticles have been the most extensively investigated. Liposomes present some technological limitations including poor reproducibility and stability as well as low drug entrapment efficiency. Polymeric nanoparticles, which possess a better reproducibility and stability profiles than liposomes have been proposed as alternative drug carriers that overcome many of these problems. Nanoparticles are solid colloidal particles with diameters ranging from 1 - 1000 nm. They consist of drug carriers in which the active ingredient is dissolved, dispersed, entrapped, encapsulated, adsorbed or chemically attached. Polymers used to form nanoparticles can be both synthetic and natural polymers. Most of the polymers prepared from water insoluble polymers are involved heat, organic solvent or harsh conditions that can be harmful to the drug stability. Moreover, some preparation methods such as emulsionpolymerization and solvent evaporation are complex and require a number of preparation steps that are more time and energy consuming. In contrast, water soluble polymers offer mild and simple preparation methods without the use of organic solvent and high shear force. Among water soluble polymers available chitosan is one of the most extensively studied. This is because chitosan possess some ideal properties of polymeric carriers for nanoparticles such as biocompatible, biodegradable, nontoxic and inexpensive. Furthermore, it possesses positively charge and exhibits absorption enhancing effect. These properties render chitosan a very attractive material as a drug delivery carrier. In the last two decades, chitosan nanoparticles have been extensively developed and explored for pharmaceutical application [8] [9] [10] [11] . Thus, the objective of the present study was designed to test the hepatoprotective activity of the ethyl acetate extract of the sponge associated bacterial extract as well as its nano chitosan loaded form as a drug delivery system.
2. Material and Methods
2.1. Sampling of the Sponge Sample
The Red Sea represents a lot of different environmental ecosystem with more than 10 thousands sea organisms. However, it started to suffer from several major threats such as urbanization and coastal development, industries including power and desalination plants and refineries, recreation and tourism, waste water treatment facilities, power plants, coastal mining and quarrying activities, oil bunkering and habitat modification such as the filling and conversion of wetlands [12] - [27] . Recently, several biologically active compounds were isolated from the Red Sea especially from marine sponges and their associated microorganisms [28] [29] [30] . Sponge sample was collected by SCUBA diving at depth 1.5 m in the Red Sea (El-Gona station; GPS: N: 27˚22'39.98'', E: 33˚40'58.95'') in May 2010 (Figure 1). The sponge sample was taxonomically identified as Amphimedonochrace [31] . Order Haplosclerida: Family Niphatidae (Figure 2). Taxonomic identification was performed by Prof. Rob. W. M. VanSoest (University of Amsterdam, Netherlands). A voucher fragment is incorporated in the collections of the Zoological Museum of Amsterdam, now part of the Netherlands

Figure 1. Egypt map showing the sponge sampling site (El-Gouna, Egypt) on Red Sea.

Figure 2. Sponge sample (Amphimedon ochracea).
Centre for Biodiversity Naturalis NCB Naturalis) at Leiden, The Netherlands, under registration number (ZMA Por. 22534). Another sponge sample was also deposited in the Marine Biotechnology and Natural Products Lab. (MBNP) at the national Institute of Oceanography and Fisheries (NIOF) under registration number (MBNP/Sp-1).
2.2. Processing of the Sponge Sample
Sponge was transferred to plastic bags containing seawater and transported to the laboratory. Sponge specimen were rinsed in sterile seawater, cut into pieces with sterile scalpels of ca. 1 cm and then thoroughly homogenized in a sterile mortar with 10 volumes of sterile seawater. The supernatant was diluted in ten- fold series (10−1, 10−2 and 10−3) using autoclaved environmental water. Fifty microliter aliquots of each dilution were placed on various agar plates’ solid media.
2.3. Isolation of Sponge Associated Bacteria
Two growth media were prepared to isolate a wide range of the sponge associated bacteria; Marine Agar [32] and ISP Medium 2 [33] served as general rich media to grow many heterotrophic marine bacteria. All media were supplemented with 0.2 µm pore size filtered cycloheximide (100 µg/ml), nystatin (25 µg/ml) and nalidixic acid (25 µg/ml). Cycloheximide and nystatin inhibit fungal growth, while nalidixic acid inhibits many fast-growing Gram-negative bacteria [34] . All media contained DifcoBacto agar (18 g/l) and were prepared in 1 lartificial seawater (NaCl, 234.7 g, MgCl2∙6H2O, 106.4 g, Na2SO4, 39.2 g, CaCl2, 11.0 g, NaHCO3, 1.92 g, KCl, 6.64 g, KBr, 0.96 g, H3BO3, 0.26 g, SrCl2, 0.24 g, NaF, 0.03 g and ddH2O to 10.0 l) [35] . The inoculated plates were incubated at 28˚C for 5 - 10 days. Distinct colony morphotypes were picked and re-streaked until visually free of contaminants. Isolates were inoculated into liquid media ISP Medium 2. The isolates were maintained on plates for short-term storage and long-term strain collections were set up in medium supplemented with 30% glycerol at −80˚C [36] .
2.4. Molecular Identification and Phylogenetic Analysis
The isolated bacteria were first identified to the species level by PCR amplification of the 16S rRNA gene, BLAST analysis, and comparison with sequences in the GenBank nucleotide database. Specifically, the 16S rRNA gene from the strain was amplified using universal primers 27f (5’-AGAGTTTGATCCTGGC- TCAG-3’) and 1492r (5’-GTTACCTTGTTACGACTT-3’). The PCR conditions used were the same as those described previously [37] . The PCR products were purified and sequenced by the GATC-Biotech. Company (Tübingen, Germany). The sequences were compared with known sequences in the GenBank nucleotide database and the species level was identified as the nearest phylogenetic neighbor with 98% - 100% similarity [38] . Sequence alignment and phylogenetic analysis were performed using the Mega-5 software package [39] . Tree construction was conducted using neighbor joining algorithm (Jukes-Cantor correction) with bootstrap values based on 1000 replications. The 16S rRNA gene sequence of the isolate was deposited in GenBank under the accession Numbers indicated in brackets: HA-MS-52 [JX484799].
2.5. Crude Extract Preparation
The bacterial isolate was cultured in 500 ml Erlenmeyer flasks containing 200 ml of ISP Medium 2. The liquid cultures were grown for 7 - 14 days depending on their growth rate at 30˚C while shaking at 150 rpm. Crude extracts were prepared from whole cultures containing cells and broths by ethyl acetate equal
volume/three times, evaporated and dried under reduced pressure using a rotary evaporator.
2.6. Gas Chromatography-Mass Spectrophotometer (GC-MS) Analysis
Analysis was conducted using gas chromatography-mass spectrophotmeter; GC-MS (Trace DSQII MS). MS conditions were as follows: Detector mass spectrometer voltage 70 eV and its source temperature was 300˚C. The injector temperature was 240˚C and the split less mode 0.5 µL injection. The HP 55% dimethyl-95% dimethyl tertiary-butyl non-polar column was performed with length 30 cm × 0.25 mm, coating thickness film 0.25 µm. The oven was adjusted at 80˚C for 1 min and initial time 1.5 min with 40˚C which ended by a final temperature of 300˚C and 4 min hold time where the total run time was 15 min. The compounds were identified by comparison with the standards, and also matched with the in-built libraries The components were identified by comparing their retention times with those of authentic samples, as well as by comparing their mass spectra with those of Wiley 275 Library [40] . Quantitative data were obtained by the peak normalization technique using integrated FID response.
2.7. Determination of Total Phenolic Contents
Total phenolic compounds in the bacterial extracts were determined by the method of Taga, et al. (1984) [41] . One hundard microliters Folin-Ciocalteau reagent (50%, v/v) in distilled water and 2 ml sodium carbonate (2%, v/v) in distilled water were added and mixed well with 100 µl of bacterial extract. After 2 h incubation at RT, the absorbance of the resulting blue color solution was measured at 750 nm. Different concentrations (0 - 0.9 mg/ml) of standard gallic acid were used and processed similarly as sample. The concentration of total phenolic in bacterial extracts was determined as mg gallic acid equivalent in 1 ml of the extract using the standard curve of the gallic acid.
2.8. Determination of Total Flavonoid Contents
The total flavonoid content was determined by a colorimetric method of Zhishen, et al. (1999) [42] . A 0.25 ml of the bacterial extract or (+)-catechin standard solution was mixed with 1.25 ml of distilled water, followed by the addition of 0.75 ml of 5% sodium nitrite solution. After 6 min, 0.15 ml of 10% aluminum chloride solution was added and the mixture was made up to 2.5 ml with distilled water and well mixed. The absorbance was measured immediately at 510 nm using a spectrophotometer. The results were expressed as mean ml of (+)-cate- chin equivalents.
2.9. Preparation of Chitosan Nanoparticles
Chitosan nanoparticles were prepared according to the ionotropic gelation pro- cess [43] . Blank nanoparticles were obtained upon the addition of a tripolyphosphate (TPP) aqueous solution (1 mg/ml) to a chitosan solution (2 mg/ml) stirredat room temperature (rate of 400 rpm). The formation of nanoparticles was a result of the interaction between the negative groups of TPP and the positively charged aminogroups of chitosan. The ratio of chitosan/TPP was established according to the preliminary studies. Extract loaded nanoparticles were obtained according to the same procedure and the ratio of chitosan/TPP remained unchanged. Nanoparticles were collected by centrifugation at 10000 rpm for 40 min and supernatant were discarded.
2.10. Determination of Loading Capacity and Efficiency of Nanoparticles
Loading capacity of extract loaded nanoparticles was determined by ultra-cen- trifugation of samples at 30000× g and 10˚C for 30 min. The amount of free extract was determined in clear supernatant by measuring the total phenolic compound of the extract supernatant of non-loaded nanoparticles as basic correction using HPLC semi-preparative. The extract loading capacity (LC) of nanoparticles was calculated from the equations indicated below:
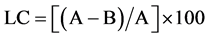
where A is the total amount of phenolic compounds in the extract; B is the free amount of phenolic compounds in the supernatant.
2.11. Animal and Experimental Design
Healthy 80 male Sprague-Dawley rats weighed 50 - 80 gm, purchased from the animal house of Faculty of Medicine, Alexandria University, and were used for the study. The animals were randomly divided into eight groups of ten animals each after an acclimatization period of two weeks. They were fed with standard diet, had free access to water, housed under standard conditions of humidity and controlled temperature (25˚C) with 12 hours light and 12 hours dark exposure. The protocol was approved by the Institutional Animal Ethics Committee (623/02/b/CPCSEA).
Group I (−ve control): Rats received saline solution subcutaneously for one week.
Group II (+ve control): Rats received subcutaneously CCl4 (1 ml/kg body weight/day) for one week (induced toxicity group).
Group III and IV were received 10 mg/Kg b.w./day bacterial extract for one week, while group V received it for two weeks and they served as one-week- treated, control and two weeks-treated groups, respectively.
Group III (one week extract-treated group): Rats received subcutaneously CCl4 (1 ml/kg body weight/day) for one week and 10 mg/Kg body weight/day of extract.
Group IV (control/extract-treated group): Rats received saline solution subcutaneously and 10 mg/Kg body weight extract for one week.
Group V (two weeks extract-treated group): Rats received subcutaneously CCl4 (1 ml/kg body weight/day) for one week (as in group II) and 10 mg/Kg body weight/day of bacterial extract for two weeks.
At the same time, Groups VI, VII and VIII were received 10 mg/Kg b.w./day of bacterial extract-chitosan loaded nanoparticles and they served as one week, two weeks and control treated nano extract-chitosan groups, respectively.
Group VI (one week nano extract-chitosan treated group): Rats received subcutaneously CCl4 (1 ml/kg body weight/day) for one week and 10 mg/Kg body weight/day of extract-chitosan loaded nanoparticles.
Group VII (two weeks nano extract-chitosan treated group): Rats received subcutaneously CCl4 (1 ml/kg body weight/day) for one week and 10 mg/Kg body weight/day of extract-chitosan loaded nanoparticles for two weeks.
Group VIII (control/nano-chitosan treated group): Rats received saline solution subcutaneously and 10 mg/Kg body weight nano-extract for one week.
At the end of experiment (one week for one-week-treated rats and two weeks for two-weeks-treated rats), rats were sacrificed under diethyl ether anesthesia.
2.12. Blood Sample Collection
Unheparinized blood samples were collected through retro-orbital puncture, allowed to clot and centrifuged at 3000 rpm to separate clear serum. Kept for 15 min at RT, sera were separated by centrifugation at 3000 rpm for 20 min and stored at −20˚C until used.
2.13. Histopathology Studies
The liver tissue was immediately removed after sacrifice and fixed in 10% formol saline. Paraffin sections of 5µ thickness were prepared, stained by Hematoxylene and Eosin (H & E) and examined microscopically.
2.14. Biochemical Measurements for the Assessment of Liver Function
a. Determination of liver function enzymes; Alanine Transaminase (ALT) and Aspartate Transaminase (AST)
Serum ALT and AST activities were determined colorimetrically using Diamond Diagnostic Kits (Egypt) according to Reitman and Frankel [44] . The absorbance of the sample (As) was read against blank at 540 nm after 5 min. the activity of the ALT and AST were expressed as U/ml and determined using standard curves.
b. Determination of serum albumin
Serum albumin was determined colorimetrically using Diamond Diagnostic Kits (Egypt) according to the method of Doumas, et al. (1971) [45] . Albumin level was calculated using the following equation:
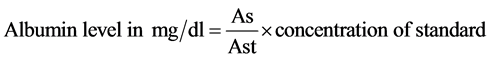
where As = sample absorbance, Ast = standerd absorbance.
c. Determination of direct bilirubin
Serum direct bilirubin was determined colorimetrically using Diamond Diagnostic Kits (Egypt), according to Hymans Van den Bergh and Mueller (1916) [46] . Direct (conjugated) bilirubin couples directly with a diazonium salt of 3,5-dichloroaniline (DPD) in an acid medium to form azobilirubin. The direct bilirubin in serum is directly proportional to the color development of azobilirubin which is measured bichromatically at 540/600 nm.
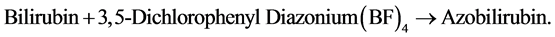
2.15. Histopathological Study
The fixed liver tissues in formalin were dehydrated in ascending grades of alcohol, and then cleaned by immersing the tissues in xylene for 1 h (three times), followed by impregnation in melted paraffin, in wax, then in oven at 60˚C for 1 h. The specimens were embedded in paraffin and were left to solidify at RT. Using a rotatory microtone, sections of 5 µm thick were cut and mounted on clean glass slides. Sections were stained with hematoxylin and eosin (H & E) and examined for any histopathological changes [47] .
Haematological Methods
Haemoglobin (Hb) concentration, packed cell volume (PCV), red blood cell count (RBC), mean corpuscular volume (MCV), and mean corpuscular haemoglobin concentration (MCHC) were measured by the method of Schalm, et al. (1975) [48] .
2.16. Statistical Analysis
Results were reported as Mean ± S.D. All biochemical analyses were carried out in triplicate measuring, and data were tested by one-way analysis of variance using SPSS program, version 11.5, *p < 0.05 regarded as significant.
3. Results and Discussion
3.1. Phylogenetic Analysis of the Bacterial Strain
The bacterial isolate was identified using 16S ribosomal DNA sequences [49] . The tested isolates belong to the Stenotrophomonas sp. (Proteobacteria) (Figure 3). Interestingly, the Proteobacteria have been reported from diverse marine sponges irrespective of their taxonomic identity, geographic location, or natural products profile [50] [51] . The isolate HA-MS-52 revealed species-level similarity to Stenotrophomonas maltophilia.
3.2. GC-MS Analysis
Capillary GC in combination with mass spectrometry was found to be the method of choice for the analysis of complex mixtures of bioactive compounds. This technique allows much more precise alkaloidal identifications to be made than by comparative TLC and GLC methods, and enables even trace constituents to be analyzed [52] . The crude extracts of the bacterial isolate HA-MS-52 was subjected to GC/MS analysis, and it revealed 6 compounds (Table 1, Figure 4).

Figure 3. Molecular phylogenetic analysis of the bacterial isolate HA-MS-52 using Maximum Likelihood Method. The scale bar indicates 0.5 substitutions per nucleotide position.

Table 1. Bioactive compounds identified in the crude bacterial extract by Gas-Chroma- tography-Mass Spectrophotometer (GC-MS) Analysis.
All the revealed compounds were identified regarding to the internal database which confirmed the presence of a variety of natural product families including mono- and di-terpenes, steroids, phenolics, flavonoids, carotenoids and fatty acids. The occurrence of different bioactive compounds suggests a wide range of biological application of the tested extracts. Sponges are important source of functional components for the development of new chemotherapeutic agents. Phytochemical investigation of the ethyl acetate extracts of the studied sponge- associated bacteria revealed the presence of various phytochemical such as phenolic compounds, flavonoids, saponins, steroids and tannins.

Figure 4. Gas Chromatography-Mass Spectrophotometer (GC-MS) chromatogram of ethyl acetate crude bacterial extract.
3.3. Total Phenolic Content and Total Flavonoid Contents
Highly significant quantities of phenolic contents were found in the crude bacterial extract measured as mg gallic acid equivalent/ml (2.7491 ± 0.046). The extract also shows highly significant quantities of flavonoid contents measured as equivalents of +catechin (12.6 ± 0.634 mg catechin equivalent/ml).
3.4. Loading of Bacterial Extract No. 5 on the Prepared Chitosan Nanoparticles Physical Characterization of Chitosan Nanoparticles
Stability of nanoparticles in suspension is directly proportional to zeta potential. The zeta potential of nanoparticles was found to be 30 mV. The observed positive zeta potential might be due to residual amine group which act as carrier of the bacterial extract.
3.5. Assessment of Liver Function
a. Liver function enzymes; Alanine Transaminase (ALT) and Aspartate transaminase (AST)
Figure 5 shows that the activity of both serum ALT and AST enzymes (markers for the liver function) were significantly (p = 0.002) elevated for the rats in Group II i.e., +ve control group (carbon tetrachloride treated group) comparing to rats in Group I, i.e., −ve control one. Administration of 10 mg/Kg body weight

Figure 5. Levels of alanine aminotransferase (ALT) and aspartate transferase (AST) enzymes in rats era of the eight animal groups (Data are means of three measurements). *ANOVA test is significant at p ≤ 0.05.

Figure 6. Chitosan nanoparticles and Extract-loaded chitosan nanoparticles as shown under the transmission electron microscope (TEM). Numerical values are shown in white box represent the particle diameter size on nm. The particle size of the extract- loaded chitosan nanoparticles showed difference from those of placebo particles (extract-non-loaded nanoparticles). The prepared chitosan nanoparticles in the placebo group were in average size of 443.95 ± 90.580 nm. Whereas, the extract-loaded nanoparticles was in average size of 684 ± 243.032nm (Figure 6).
of the ethyl acetate bacterial extract (bacterial isolate HA-MS-52) to rats in Group III (One week extract-treated group) and Group V (Two weeks extract- treated group) remarkably prevented the effect of carbon tetrachloride which induced elevation of serum ALT and AST as shown in Group II (+ve control group).
Effects of the bacterial extract on the activities of serum ALT and AST in rat of CCl4 treated groups either as plain extract (Groups III and V) or loaded in chitosan nanoparticles (Groups VI and VII) is shown in Table 2 and Figure 5. Group II (+ve control group) showed marked elevation of serum ALT and AST levels (56.19 ± 28.09 U/l and 71.51 ± 24.94 U/l, respectively) comparing to group I; −ve control (21.38 ± 4.03 U/l and 7.72 ± 2.16 U/l, respectively). Group III (One week extract-treated group), showed marginal reversal of elevated ALT and AST levels in serum (46.76 ± 14.16 U/l and 56.44 ± 20.68 U/l, respectively).
At the same time, the administration of the bacterial extract for one week more in Group V (two weeks extract-treated group) showed marked elevation of serum ALT and AST levels comparing to Group III (One week extract-treated group) (24.93 ± 7.44 U/l and 41.37 ± 19.69 U/l, respectively). The serum AST and ALT levels of rats in Group IV (control extract-treated group) showed slight elevation on (27.56 ± 7.11 U/l and 13.67 ± 22.79 U/l, respectively) comparing to Group I (−ve control group).
On the other hand, Group VI (One week nano extract-chitosan treated group) exhibited a higher reversal of elevated serum ALT and AST levels comparing to their levels in Group III, i.e., one week extract-treated group as well as Group V, i.e., two weeks extract-treated group (18.96 ± 7.29 U/l and 31.85 ± 7.35 U/l, respectively). However, Group VI (one week nano extract-chitosan treated rats) showed lower reversal of elevated ALT and AST levels as compared to Group III (one week extract-treated group) (69.41 ± 28.21 U/l and 56.44 ± 20.68 U/l, respectively). On the other hand, Group VII (one week nano extract-chitosan treated rats) showed the best reversal effect of ALT and AST levels among all induced groups (groups III, V, VI and VII), especially when compared to the two weeks extract-treated rats in Group V (10.85 ± 7.35 U/l and 41.37 ± 19.69 U/l, respectively). Moreover, serum ALT and AST levels in control nano-chito- san treated rats (Group VIII) showed lower reversal effect of the elevated serum ALT and AST levels when compared to the two weeks nano extract-chitosan treated rats in Group VII (39.91 ± 15.82 U/l and 30.38 ± 9.77 U/l, respectively).
Table 2. Serum levels of AST and ALT enzymes among the different rat groups.
*ANOVA test is significant at p ≤ 0.05.
b. Levels of serum albumin
Low significance observation (p = 0.1) has been shown in serum albumin level among all experimental groups (Table 3). Group I values ranged from (1.748 - 2.636 g/dl), Group II (2.494 - 4.604 g/dl), Group III (2.352 to 3.417 g/dl), Group IV (1.734 - 3.162 g/dl), Group V (1.719 - 3.304 g/dl), Group VI (1.130 - 2.821 g/dl), Group VII (2.238 - 3.460 g/dl) and Group VIII (1.854 - 3.432 g/dl). Based on comparison between the mean values, the maximum albumin levels were recorded among animals in Group II and the lowest were among Group IV (Figure 7).
c. Levels of serum direct bilirubin
Serum direct bilirubin differs significantly (p = 0.02) among all groups but with low significance (Table 4). Group I (the control group) values ranged from (1.008 - 1.224 mg/dl), Group II (2.491 - 2.678 mg/dl), Group III (1.037 - 2.736 mg/dl), Group IV (0.965 - 2.592 mg/dl), Group V (0.950 - 1.224 mg/dl), Group VI (0.965 - 1.138 mg/dl), Group VII (1.152 - 1.382 mg/dl) and Group VIII (0.994 - 1.152 mg/dl). Based on comparison between the mean values, the maximum
Table 3. Levels of albumin in rat sera of the different eight animal groups.
*ANOVA test is significant at p ≤ 0.05.

Figure 7. Levels of albumin in rat sera of the different eight animal groups (data are means of three measurements). *ANOVA test is significant at p ≤ 0.05.
Table 4. Levels of direct bilirubin in rat sera of the eight animal groups.
*ANOVA test is significant at p ≤ 0.05.
albumin levels were observed among animals in Group II and the lowest were among Group IV (Figure 8). Figure 8 shows all the in vivo values of the biochemical parameters measured in the eight animal groups.
3.6. Histopathology of the Liver Samples
The photomicrographs of the liver sections of all rat groups under investigation were given in Figure 8. Histopathological profile of the control animals showed normal hepatic architecture with discrete hepatic cells, well preserved cytoplasm and sinusoidal spaces (Figure 8). Disarrangement of normal hepatic cells with intense centrilobular necrosis was observed in Figure 8 of Group II (+ve control group; induced group) CCl4 intoxicated liver (Figure 8). Moderate accumulation of fatty lobules and cellular necrosis were observed in the animal treated with 10 mg bacterial extract suspension either as plain or loaded into chitosan nanoparticles (Figure 8). Conclusively, the areas of necrosis and ballooning degeneration of hepatocytes were observed in the toxic groups. The group of animals treated with the extract showed a marked protective effect with decreased necrotic zones and hepatocellular degeneration.
3.7. Complete Blood Analysis
a. Effect of the bacterial extract on RBCs and related indices
CCl4 significantly decreased RBCs counts and Hb concentration. Administration of the bacterial extract significantly increased the RBCs count, Hb and haematocrite concentration when compared to control or carbon tetrachloride-in- toxicated group as shown in (Table 5).
b. Effect of the bacterial extract on total and differential leucocytic counts
CCl4 significantly affected total and differential leucocytes counts. There was a significant difference in the total WBCs count between all groups. Lymphocytes and eosinophiles percent were significantly increased by CCl4 in (Group I). Monocyte percent was significantly increased in extract treated rats (Group III)

Figure 8. Levels of direct bilirubin in rat sera of the eight animal groups (data are means of three measurements). *ANOVA test is significant at p ≤ 0.05.
Table 5. Effect the bacterial extract (plain and chitosan-loaded nanoparticles) on some hematological indices among the eight rat groups.
RBC: Red blood corpuscles, Hb: Hemoglobin, HCT: Haematocrite, MCV: Mean corpuscular volume, MCH: Mean corpuscular hemoglobin, MCHC: Mean corpuscular hemoglobin concentration. Values are Mean ± SD. *ANOVA test is significant at p ≤ 0.05.
(Table 6).
Effect on blood platelets
Results indicated that platelet counts (PLT) × 103 µl in control group was 632.22 ± 71.72. In CCl4-intoxicated rats, platelet counts, platelet distribution width (PDW) and mean platelet volume (MPV) was significantly (p > 0.05) decreased in comparison to normal control. Administration of bacterial extract either in the form of plain emulsion or carried into Thenano chitosan particles significantly increased platelets counts, platelet distribution width (PDW) and mean platelet volume (MPV) to levels nearly similar to that of normal control (Table 7).
The elevated levels of serum enzymes, ALT and AST of rats dosed with CCl4 to induce liver injury (+ve control group) are indicative of cellular leakage and loss of functional integrity of liver cell membrane (Table 2) [53] [54] . These values are significantly higher in Group II (+ve control group) comparing to Group I (−ve control group) indicating a possible induction of liver damage using CCl4 [55] . Although certain factors such as sex differences, haemolysis of red blood cells, presence of activators and inhibitors as well as presence of pyridoxine
Table 6. Effect of the bacterial extract (plain and chitosan-loaded nanoparticles) on total and differential leucocytes count among the eight rat groups.
WBcs: White blood corpuscules, NEUT: Neutrophils, LYMPH: Lymphocytes, MONO: Monocytes, EO: Eosinphils. Values are means ± S.D. *ANOVA test is significant at p ≤ 0.05.
Table 7. Effect of the bacterial extract (plain and chitosan-loaded nanoparticles) on PLT, PDW and MPV among the eight rat groups.
PLT: Platelets, PDW: Platelet distribution width, MPV: Mean platelet volume. Values are means ±S.D. *ANOVA test is significant at p ≤ 0.05.
(Vitamin B6), may influence the serum AST level since the concentration of AST in erythrocyte is roughly tenfold than normal serum level [56] . The concurrent elevations of serum AST together with ALT indicates that the above factors may likely not be the cause of the elevated serum enzyme activities, but rather it is more likely to be due to the toxicity induced by CCl4. Treatment with the ethyl acetate bacterial extract decreased significantly the serum levels of ALT and AST. At the same time, the group treated with the extract-chitosan nanoparticles, decreased significantly the serum levels of ALT and AST comparing to +ve control group and approaching their values of −ve control group. This result could be due to the stabilization of plasma membrane as well as the repair of hepatic tissue damage caused by CCl4 (Table 2 and Figure 7). The biochemical findings were further substantiated by the liver histological examinations for the liver. CCl4 produces an experimental damage that histologically resembles viral hepatitis [57] . Toxicity begins with the change in endoplasmic reticulum, which results in the loss of metabolic enzymes located in the intracellular structures [51] . The toxic metabolite CCl3 radical is produced which is further converted to trichloromethylperoxy radical by cytochrome P450 2E1 enzyme. This radical binds covalently to the macromolecules and causes peroxidative degradation of cellular membrane leading to the necrosis of hepatocytes [58] .
The concentration of serum direct bilirubin level in Group II (+ve control group, Table 4 and Figure 8) increased significantly comparing to its level in the −ve control group (Group I). The value of serum direct bilirubin indicates the presence of hyperbilirubunaemia, which could be due to liver dysfunction. However, hyperbilirubunaemia is not always present in well compensated cirrhosis [59] and the presence of increased direct bilirubin is probably an indication of acute hepatitis induced by CCl4. The serum levels of albumin in +ve control group (Group II) was found to be significantly decreased comparing to its corresponding value of the −ve control one (Group I). The decrease in serum albumin level in the induced toxicity group i.e., +ve control group (Group II) is associated with hepatic damage which may lead to impaired synthesis of albumin due to impairment of hepatic function and the effect may be severe in chronic hepatitis. The administration of the bacterial extract to Groups III and IV for 7 days using CCl4 as well as to Groups V and VII induced hepatotoxicity for 14 days, reduced significantly (p < 0.05) the levels of serum AST, ALT as well as direct bilirubin comparing to their levels in Group II (+ve control group).The significant decrease in liver function indices in the bacterial extract administered animals might be due to the decreased leakage of liver hepatocytes enzymes. Accordingly, this indicates that the bacterial extract may reduce and/or reversed the toxic effect of CCl4 and prevent leakage of hepatocyte enzymes into blood circulation. The reduction in serum albumin in Group II (administered with 1 ml/Kg CCl4 ; +ve control group) was significantly elevated when the rats treated with bacterial extract as shown in Groups III and IV, as well as Groups V and VI treated with nano-extract. This elevation could be related to the curative effect of bacterial extract against liver cell damage induced by CCl4. Clawson (1989) [60] reported that, the decline in protein content may be due to defect in protein biosynthesis as well as disruption and disassociation of polyribosomes from endoplasmic reticulum following administration of CCl4. According to Rajesh and Latha (2004) [61] , the improvements in the level of serum albumin of induced hepatotoxicity rats after their treatment with the natural products may be due to the promotion of ribosome assembly on endoplasmic reticulum which facilitates uninterrupted protein biosynthesis [55] .
The histopathological changes were observed in the present study including ballooning and fatty degeneration, congestion, reactive changes and loss of normal structure of hepatocytes of rats induced toxicity group (+ve control group), in comparison with Group I (?ve control group). These changes were nearly normalized, when bacterial extract, in dose of 10 mg/Kg b.w. was administered either loaded in chitosan nanoparticles (Groups VI and VII) or not loaded in chitosan nanoparticles (Groups III and V). Many phytoconstituents present in the bacterial extract, are reported to possess hepatoprotective property, like phenolic compounds [61] . Flavonoids and polyphenols [62] , terpenoids [63] and glycosides [64] also were reported to possess antioxidant properties which in turn could exert hepatoprotective effects.
Hepatocytes of ?ve control rats group have a regular morphological structure (Figure 9(A)). In case of Group II (+ve control group), when bacterial extract, in dose of 10 mg/Kg b.w. was co-administered with CCl4 for one week, some of the liver cells were damaged while the rest were nearly normalized as shown in Figure 9(C). In Group III (extract treated control): treated by the 10 mg/Kg b.w. bacterial extract; showed normal hepatic architecture (Figure 9(D)). In case of Group IV (control extract treated group) where rats were treated by 10 mg/Kg b.w. extract with CCl4 for one week revealed some hepatic cells in normal case but the others were damaged (Figure 9(E)). When the rats were treated by the same extract dose for week more without CCl4 intoxication, the rats’ hepatocytes were normal. In case of Group V (two weeks bacterial extract treated group) the liver cells showed the central vein and necrosis while some cells appeared in normal shape as shown in Figure 9(F). The liver of Group VI (one week nano








Figure 9. (A) Histopathological examination of −ve control group (Group I). The liver is showing normal hepatic architecture with discrete hepatic cells, well preserved cytoplasm and sinusoidal spaces; (B) Histopathological examination of +ve control group (Group II). The liver is showing disarrangement of normal hepatic cells with intense centrilobular necrosis and ballooning degeneration of hepatocytes; (C) Histopathological examination of one week extract-treated group (Group III). The liver is showing mild necrosis; (D) Histopathological examination of the control extract-treated group (Group IV). The liver is showing mild necrosis; (E) Histopathological examination of two weeks extract-treated group (Group V). The liver is showing moderate accumulation of fatty lobules and cellular necrosis; (F) Histopathological examination of two weeks extract-treated group (Group VI). The liver is showing moderate accumulation of fatty lobules and cellular necrosiswith decreased necrotic zones and hepatocellular degeneration; (G) Histopathological examination of two weeks extract-treated group (Group VII). The liver is showing decreased necrotic zones and hepatocellular degeneration; (H) Histopathological examination of two weeks extract-treated group (Group VIII). The liver is showing moderate accumulation of fatty lobules and cellular necrosis.
extract-chitosan treated group), necrotic and lipid cells were observed as shown in Figure 9(G), while Group VIII (control nano-chitosan treated group): treated by the 10 mg/Kg b.w. nano-chitosan (no extract) with CCl4 for one week; their liver showed necrotic cells (Figure 9(H)). Accordingly, we could say that the bacterial extract, is the more responsible factor for the hepatic protection against CCl4 toxicity rather than chitosan.
c. Effect of the bacterial extracton Blood parameters
Although no available literature could be obtained concerning the effect of the tested bacterial ethyl acetate extracts on blood cellular elements, however the increased RBCs count and Hbas well as HCT of rats’ blood in extract-treated rats could be attributed to its active constituents. The increased monocyte counts in blood of rats treated with ethyl acetate bacterial extractmaybe related to the anti- inflammatory effect of their active constituents [65] [66] and to their immune protectant effect [67] [68] . The present data clearly demonstrate that the administration of ethyl acetate bacterial extract either as plain emulsion or loaded in chitosan nanoparticles has restored the platelets counts which were decreased after CCl4-induced intoxication.
4. Conclusion
In conclusion, the results of the present study suggest that the bacterial extract has an ability to protect the liver against CCl4-induced liver damage, and it could be associated with decreased oxidative stress as well as free radical-mediated tissue damage. Delivery of the bacterial extract through chitosan nanoparticles is a better way to improve the efficacy of result in expression of bioactivity of the bacterial extract as a hepatoprotective agent.
Cite this paper
Abdel Monein, N.M., Yacout, G.A., Aboul-Ela, H.M. and Shreadah, M.A. (2017) Hepatoprotective Activity of Chitosan Nanocarriers Loaded with the Ethyl Acetate Extract of a Stenotrophomonas sp. Bacteria Associated with the Red Sea Sponge Amphimedon ochracea in CCl4 Induced Hepatotoxicty in Rats. Advances in Bioscience and Biotechnology, 8, 27-50. http://dx.doi.org/10.4236/abb.2017.81003
References
- 1. Priya, S. and Nelhaji, S. (2015) Hepatoprotective Activity of Ethanolic Extract of Diospyros Virgininana in CCl4 Induced Hepatotoxicity in Swiss Albino Rats. International Journal of PharmTech Research, 8, 444-447.
- 2. Ravikiran, Y., Manjunath, C., Balasubramannian, T., Gnanasekaran, D., Ashokkumar, U. and Brahmaiah, Y. (2012) Evaluation of Hepatoprotective Activity of Haldinia Cordifolia against Parcetamol Induced Liver Damage in Rats. Der Pharmacia Lettre, 4, 768-774.
- 3. Tonkiri, A., Essien, E.S. and Akaninwor, J.O. (2014) Evaluation of Hepatoprotective and in Vivo Antioxidant Activity of the Methanolic Stem Extract of Costusafer. (Bush Cane) in Alcohol Induced Liver Cirrhosis in Rats. Journal of Biological and Food Science Research, 3, 29-34.
- 4. Aboul-Ela, H.M., Shreadah, M.A., Abdel-Monem, N.M., Yakout, G.A. and VanSoest, R.W.M. (2012) Isolation, Cytotoxic Activity and Phylogenetic Analysis of Bacillus sp. Bacteria Associated with the Red Sea Sponge Amphimedon ochracea. Advances in Bioscience and Biotechnology, 3, 815-823. https://doi.org/10.4236/abb.2012.37101
- 5. Mondol, M.A., Shin, H.J. and Islam, M.T. (2013) Diversity of Secondary Metabolities from Marine Bacillus Species: Chemistry and Biological Activity. Marine Drugs, 11, 2846-2972.
https://doi.org/10.3390/md11082846 - 6. Mehbub, M.F., Franco, J.L.C. and Zhang, W. (2014) Marine Sponge Derived Natural Products between 2001 and 2010: Trends and Opportunities for Discovery of Bioactivities. Marine Drugs, 12, 4539-4577. https://doi.org/10.3390/md12084539
- 7. Livett, B.G., Gaylera, K.R. and Zhang, Z. (2004) Drugs from the Sea: Conopeptides as Potential Therapeutics. Current Medicinal Chemistry, 11, 1715-1723.
https://doi.org/10.2174/0929867043364928 - 8. Newman, D.J. and Cragg, G.M. (2004) Marine Natural Products and Related Compounds in Clinical and Advanced Preclinical Trials. Journal of Natural Products, 67, 1216-1238.
https://doi.org/10.1021/np040031y - 9. Simmons, T.L. andrianasolo, E., McPhail, K., Flatt, P.M. and Gerwick, W.I. (2005) Marine Natural Products as Anticancer Drugs. Molecular Cancer Therapeutics, 4, 333-342.
- 10. Thomas, C. and Shrupp, P. (2005) Biotechnological Potential of Marine Sponges and Their Associated Bacteria as Producers of New Pharmaceuticals. Journal of International Biotechnology Law, 2, 257-264. https://doi.org/10.1515/jibl.2005.2.5.217
- 11. Micheal, W., Taylor, R., Doris, S. and Michael, W. (2007) Sponge-Associated Microorganisms: Evolution, Ecology, and Biotechnological Potential. Microbiology and Molecular Biology Reviews, 71, 295-347. https://doi.org/10.1128/MMBR.00040-06
- 12. Dharmara, S., Ashokkumar, B. and Dhevendaran, K. (2010) Isolation of Marine Streptomyces and the Evaluation of Its Bioactive Potential. African Journal of Microbiology Research, 4, 240-248.
- 13. Hentschel, U., Piel, J., Degnan, S.M. and Taylor, M.W. (2012) Genomic Insights into the Marine Sponge Microbiome. A Review. Nature Reviews Microbiology, 1, 641-654.
https://doi.org/10.1038/nrmicro2839 - 14. Shriadah, M.A., Okbah, M.A. and El-Deek, M.S. (2004) Trace Metals in the Water Columns of the Red Sea and the Gulf of Aqaba, Egypt. Water, Air, & Soil Pollution, 153, 115-124.
https://doi.org/10.1023/B:WATE.0000019938.57041.21 - 15. Okbah, M.A., Shata, M.A. and Shriadah, M.A. (2005) Gochemical Forms of Trace Metals in Mangrove Sediments-Red Sea. Chemistry and Ecology, 21, 23-36.
https://doi.org/10.1080/02757540512331323953 - 16. Fahmy, M.A., Shriadah, M.A., AbulSoud, A., Abdel Rahman, S.M. and Shindy, M. (2005) Hydrography and Chemical Characteristics of the Coastal Water along the Gulf of Suez. Egyptian Journal of Aquatic Research, 31, 1-14.
- 17. Abdel-Halim, A.M., Aboel-Khair, E.M., Fahmy, M.A. and Shreadah, M.A. (2007) Environmental Assessment on the Aqaba Gulf Coastal Waters, Egypt. Egyptian Journal of Aquatic Research, 33, 1-14.
- 18. Abo-el-Khair, E.A., Abdel Halim, A.M., Shriadah, M.A. and Fahmy, M.A. (2007) Environmental Conditions of the Suez Gulf and the Red Sea Coastal Waters, Egypt. Proceedings of the 8th International Conference on the Mediterranean Coastal Environment, Alexandria, 13-17 November 2007, 517-526.
- 19. Abo-El khair, E.M., Abdel Halim, A.M., Fahmy, M.A. and Shreadah, M.A. (2008) Environmental Impact Assessment of Northern Red Sea Regions during 2005-2007. Egyptian Journal of Aquatic Research, 34, 20-30.
- 20. Shreadah, M.A., Said, T.O., El Zokm, G. and Masoud, M.S. (2008) Physico-Chemical Characterititics of the Surficial Sediments along the Egyptian Red Sea Coasts. Egyptian Journal of Aquatic Research, 34, 16-34.
- 21. Shreadah, M.A., Said, T.O, Abd El Ghani, S.A. and Ahmed, A.M. (2008) Alkyllead and Alkyltin Species in Different Fishes Collected from the Suez Gulf, Egypt. Egyptian Journal of Aquatic Research, 34, 64-73.
- 22. Shreadah, M.A. Masoud, M.S., Said, T.O. and El Zokm, G. (2008) Application of IR, X-Ray, TGA and DTA to Determine the Mineral Composition of the Sediments and Study of Reaction Kinetics along the Egyptian Red Sea Coasts. Egyptian Journal of Aquatic Research, 34, 83-95.
- 23. Gurguess, S.M. Shreadah, M.A., Fahmy, M.A. Aboul El Kheir, E. and Abdel Halim, A.M. (2009) Assessment of Water Quality in the Red Sea Using in Situ Measurements and Remote Sensing Data. Egyptian Journal of Aquatic Research, 35, 1-13.
- 24. Said, T.O., Shreadah, M.A., Abdel Ghani, S.A. and Abdel Ahmed, A.M. (2010) Alkyltin and Alkayllead Compounds in Coastal Water of Suez Gulf, Egypt. Egyptian Journal of Aquatic Research, 36, 33-42.
- 25. Masoud, M.S., Said, T.O., El-Zokm, G. and Shreadah, M.A. (2010) Speciation of Fe, Mn and Zn in Surficial Sediments from the Egyptian Red Sea Coasts. Chemical Speciation and Biodiversity, 22, 257-269. https://doi.org/10.3184/095422910x12894975123773
- 26. Masoud, M.S., Said, T.O. El-Zokm, G. and Shreadah, M.A. (2012) Assesment of Heavy Metals Contamination in Surface Sediments of the Egyptian Red Sea Coasts. Australian Journal of Basic and Applied Sciences, 6, 44-58.
- 27. Shreadah, M.A., Said, T.O., Abdel Ghani, S.A. and Ahmed, A.M. (2011) Distribution of Different Organotin and Organolead Compounds in Sediment of Suez Gulf. Journal of Environmental Protection, 2, 545-554. https://doi.org/10.4236/jep.2011.25063
- 28. Hegazy, M.F., Mohamed, T.A., Elshamy, A.I., Hassanien, A.A., Abdel Azimd, N.S., Shreadah, M.A., Abdel Gawad, I.I. and El kady, E.M. (2015) A New Steroid from the Red Sea Soft Coral Lobophytum lobophytum. Natural Products Research, 30, 340-344.
https://doi.org/10.1080/14786419.2015.1046871 - 29. Hegazy, M.F., Gamal-Eldeen, A.M., Mohamed, T.A., Alhammady, M.A., Hassanien, A.A., Shreadah, M.A., Abdelgawad, I.I. and Elkady, E.M. (2015) Cytotoxic Constituents from the Red Sea Soft Coral Nephthea sp. Natural Products Research, 30, 1266-1272.
https://doi.org/10.1080/14786419.2015.1055266 - 30. Poksch, P., Edrada, R. and Ebel, R. (2006) Implications of Marine Biotechnology on Drug Discovery, In: Proksch, P. and Muller, W., Eds., Frontiers in Marine Biotechnology, Horizon Bioscience, Norfolk, 1-19.
- 31. Keller, C. (1889) Die Spongienfauna des rothen Meeres (I. Halfte). Zeitschrift für Wissenschaftliche Zoologie, 48, 311-405.
- 32. Weiner, R.M., Segall, A.M. and Colwell, R.R. (1985) Characterization of Marine Bacterium Associated with Crassostrea virginica. Applied and Environmental Microbiology, 49, 83-90.
- 33. Shilling, E.B. and Gottlieb, D. (1996) Methods for Characterization of Streptomyces Species. International Journal of Systemic Bacteriology, 16, 313-340.
https://doi.org/10.1099/00207713-16-3-313 - 34. Okwu, D.E. (2004) Phytochemical and Vitamin Content of Indigenous Species of South-Eastern Nigeria. Journal of Sustainable Agriculture and the Environment, 6, 30-37.
- 35. Evans, W.C. (1989) Trease and Evans Pharmacognosy. 13th Edition, Balliere Tindall, London, 378-689.
- 36. Sofowora, A. (1993) Medicinal Plants and Traditional Medicine in Africa. 2nd Edition, Spectrum Books Limited, Ibadan, 134-156.
- 37. Odebiyi, A. and Sofowora, A.E. (1978) Phytochemical Screening of Nigerian Medicinal Plants, Part III. Lloydia, 41, 234-246.
- 38. Edeoga, H.O., Okwu, D.E. and Mbaebie, B.O. (2005) Phytochemical Constituents of Some Nigeria Medicinal Plants. African Journal of Biotechnology, 4, 685-688.
https://doi.org/10.5897/AJB2005.000-3127 - 39. Sambrook, J., Fritsh, E.F. and Maniatis, T. (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, New York.
- 40. Antolovich, M., Prenzler, P.D., Patsalides, E., Mc Donald, S. and Robards, K. (2002) Methods for Testing Antioxidant Activity. Analyst, 127, 183-198. https://doi.org/10.1039/b009171p
- 41. Tega, M.S., Miller, E.E. and Pratt, D. (1984) Chia Seeds as a Source of Natural Lipid Antioxidants. Journal of the American Oil Chemists’ Society, 61, 928-931.
https://doi.org/10.1007/BF02542169 - 42. Jia, Z., Tang, M. and Wu, J. (1999) Research on Antioxidant Activity Offalvonoids from Natural Material. Food Chemistry, 64, 555-559. https://doi.org/10.1016/S0308-8146(98)00102-2
- 43. Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M. and Rice-Evans, C. (1999) Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorizing Assay. Free Radical Biology and Medicine, 26, 1231-1237. https://doi.org/10.1016/S0891-5849(98)00315-3
- 44. Reitman, S. and Frankel, S. (1957) A Colorimetric Method for the Determination of Serum Glutamic Oxalacetic and Glutamic Pyruvic Transaminases. American Society of Clinical Pathologists, 28, 56-63. https://doi.org/10.1093/ajcp/28.1.56
- 45. Doumas, B.T., Walson, W.A. and Biggs, H.G. (1971) Albumin Standards and the Measurement of Serum Albumin with Bromcresol Green. Clinica Chimica Acta, 31, 87-96.
https://doi.org/10.1016/0009-8981(71)90365-2 - 46. Van den Bergh, A.A. and Mueller, P. (1916) Ueber eine direkte und eine indirekte Diazoreaktion auf Bilirubin. Biochemische Zeitschrift, 77, 90-103.
- 47. Griffith, J. and Farris, E. (1942) The Osseous System. In: Griffith, J. and Farris, E., Eds., The Rat in Laboratory Investigation, JB Lippincott, Philadelphia, 418-419.
- 48. Schlam, O.W., Jain, N.C. and Carol, E.I. (1975) Veterinary Heamatology. 3rd Edition, Lea and Febiger, Philadelphia, 144-167.
- 49. Proksch, P., Edrada, R. and Ebel, R. (2002) Drugs from the Seas-Current Status and Microbiological Implications. Applied Microbiology and Biotechnology, 59, 125-134.
https://doi.org/10.1007/s00253-002-1006-8 - 50. Weinheimer, A. and Spraggins, R. (1969) The Occurrence of Two New Prostaglandin Derivatives (15-epi-pGA2 and Its Acetate, Methyl Ester) in the Gorgonian Plexaura homomalla Chemistry of Coelenterates. Tetrahedron Letters, 15, 5185-5188.
https://doi.org/10.1016/S0040-4039(01)88918-8 - 51. Bergmann, W. and Feeney, R. (1951) Contribution on the Study of Marine Sponges, 32. The Nucleosides of Sponges. Journal of Organic Chemistry, 16, 981-987.
https://doi.org/10.1021/jo01146a023 - 52. Taki, M., Burgoyne, D.L. and Ersen, R.J. (1994) Effect of Contignasterol on Histamine Release Induced by Anti-Immunoglobulin E from Rat Peritoneal Mast Cells. Journal of Pharmaceutical Sciences, 83, 1234-1235. https://doi.org/10.1002/jps.2600830909
- 53. Park, J., Saravanakumar, G., Kim, K. and Kwon, I. (2010) Targeted Delivery of Low Molecular Drugs Using Chitosan and Its Derivatives. Advanced Drugs Delivery Reviews, 62, 28-41.
https://doi.org/10.1016/j.addr.2009.10.003 - 54. Drotman, R.B. and Lawhorn, G.T. (1978) Serum Enzymes as Indicators of Chemically Induced Liver Damage. Drug and Chemical Toxicology, 1, 163-171.
https://doi.org/10.3109/01480547809034433 - 55. Price, N.C. and Stevens, L. (2003) Fundamentals of Enzmology. 3rd Edition, Oxford University Press, Oxford, 404-406.
- 56. Alhassan, A.J., Sule, M.S., Hassan, J.A., Baba, B.A. and Aliyu, M.D. (2009) Ideal Hepatotoxicity Model in Rats Using Varying Concentrations of Carbon Teterachloride (CCl4). Bayero Journal of Pure and Applied Sciences, 2, 185-187.
- 57. James, G.W.L. and Pickering, R.W. (1976) The Protective Effect of a Novel Compound RU-18492 on Galactosamine Induced Hepatotoxicity in Rats. Drug Research, 26, 2197-2199.
- 58. Recknagel, R.O., Glende, E.A., Dolak, J.A. and Waller, R.L. (1989) Mechanism of Carbon Tetrachloride Toxicity. Pharmacology & Therapeutics, 43, 139-154.
https://doi.org/10.1016/0163-7258(89)90050-8 - 59. Ranawat, L., Bhatt, J. and Patel, J. (2010) Hepatoprotective Activity of Ethanolic Extracts of Bark of Zanthoxylum armatum DC in CCl4 Induced Hepatic Damage in Rats. Journal of Ethnopharmacology, 127, 777-780. https://doi.org/10.1016/j.jep.2009.10.019
- 60. Clawson, G.A. (1989) Mechanisms of Carbon Tetrachloride Hepatotoxicity. Pathology and Immunopathology Research, 8, 104-112. https://doi.org/10.1159/000157141
- 61. Rajesh, M.G. and Latha, M.S. (2004) Preliminary Evalutation of the Antihepatoxic Activity of Kamilari, a Polyherbal Formulation. Journal of Ethnopharmacology, 91, 99-104.
https://doi.org/10.1016/j.jep.2003.12.011 - 62. Previati, M., Corbacella, E., Astolfi, L., Catozz, M., Khan, M., Lampronti, I., Gambri, R., Capitani, S. and Martini, A. (2007) Ethanolic Extract from Hemidesmus indicus (Linn) Displays Otoprotectantactivities on Organotypic Cultures without Interfering on Gentamicin Uptake. Journal of Chemical Neuroanatomy, 34, 128-133.
https://doi.org/10.1016/j.jchemneu.2007.05.009 - 63. Lakshman, K., Jayaprakash, B. and Joshi, H. (2005) Comparative Anti-Inflammatory Activity Studies of Four Species of Sariva. Indian Journal of Pharmaceutical Sciences, 67, 481-482.
- 64. Kumar, A.K.R., Krishna, K.R., Chandramohan, G., FbKhanum, A. and Bawa, S. (2007) Beeswax Polyphenols as Suppressoprs of CCl4-Induced Oxidative Stress in Rats. Indian Journal of Physiology and Pharmacology, 51, 361-367.
- 65. Patel, P.B., Patel, T.K., Shah, P., Baxi, S.N., Sharma, H.O. and Tripathi, C.B. (2010) Protective Effects of Ethanol Extract of Gymnosporia Montana (Roth) Bemth. In Paracetamol Induced Hepatotoxicity in Rats. Indian Journal of Pharmaceutical Sciences, 72, 392-396. https://doi.org/10.4103/0250-474X.70493
- 66. Natarajan, K.S., Narasimhan, M., Radha, S.K. and Shanmugasundaram, E.R.B. (2006) Antioxidant Activity of a Salt-Spice-Herbal Mixture against Free Radical Induction. Journal of Ethnopharmacology, 105, 76-83.
https://doi.org/10.1016/j.jep.2005.09.043 - 67. Shashidhara, M.G., Giridharcb, P., UdayaSankarb, K. and Manohar, B. (2013) Bioactive Principles from Cordyceps sinensis: A Potent food Supplement—A Review. Journal of Functional Foods, 5, 1013-1030. https://doi.org/10.1016/j.jff.2013.04.018
- 68. Levy, A.S., Simon, O., Shelly, J. and Gardener, M. (2006) 6-Shogaol Reduced Chronic Inflammatory Response in the Kness of Rats Treated with Complete Freund’s adjuvant. BMC Pharmacology, 6, 12. https://doi.org/10.1186/1471-2210-6-12