Journal of Behavioral and Brain Science
Vol.04 No.12(2014), Article ID:52838,19 pages
10.4236/jbbs.2014.412054
Brain and Law: An EEG Study of How We Decide or Not to Implement a Law
Armando Freitas da Rocha1, Eduardo Massad2,3*, Fabio Theoto Rocha1,4,5, Marcelo Nascimento Burattini2,6
1Research on Natural and Artificial Intelligence (RANI), Jundiaí, Brazil
2School of Medicine, University of São Paulo Brazil, São Paulo, Brazil
3London School of Hygiene and Tropical Medicine, London, UK
4Instituto de Pesquisas em Teoria da Informação (IPTI), Aracajú, Brazil
5Laboratory of Signal Processing, Department od Electrical Engeneering, Faculdade de Engenharia Industrial, São Paulo, Brazil
6Escola Paulista de Medicina, Universidade Federal de São Paulo, São Paulo, Brazil
Email: *edmassad@usp.br
Copyright © 2014 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY).
http://creativecommons.org/licenses/by/4.0/



Received 6 October 2014; revised 23 November 2014; accepted 7 December 2014
ABSTRACT
Brazil has introduced a referendum regarding the prohibition of firearm commerce and propaganda arguments have invoked socially and personally driven issues in the promotion of voting in favor of and against firearm control, respectively. Here, we used different techniques to study the brain activity associated with a voter’s perception of the truthfulness of these arguments and their influence on voting decisions. Low-resolution tomography was used to identify the possible different sets of neurons activated in the analysis of the different types of propaganda. Linear correlation was used to calculate the amount information 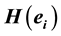 provided to different electrodes about how these sets of neurons enroll themselves to carry out this cognitive analysis. The results clearly showed that vote decision was not influenced by arguments that were introduced by propaganda, which was typically driven by specific social or self-interest motives. However, different neural circuits were identified in the analysis of each type of propaganda argument, independently of the declared vote (for or against the control) intention.
provided to different electrodes about how these sets of neurons enroll themselves to carry out this cognitive analysis. The results clearly showed that vote decision was not influenced by arguments that were introduced by propaganda, which was typically driven by specific social or self-interest motives. However, different neural circuits were identified in the analysis of each type of propaganda argument, independently of the declared vote (for or against the control) intention.
Keywords:
EEG, Brain Mapping, Neuromarketing, Language Understanding, Politics, Election, Decision Making

1. Introduction
As discussed before [1] : Any living being uses all possible resources to keep maintaining its identity in space and time. This means that any living being has as primordial goals to obtain not only the resources to synthesize their constituent chemicals, to obtain energy to support the chemical transactions and to allow it to creating other similar living beings, but also to achieve some goals dependent on some more abstract matters. Because of this, living beings have sets of goals (values) to attain and as resources on Earth are finite, this creates needs for goods and services to support FSV. Needs in turn motivates the individual to act in order to satisfy them.
Man is a social being and living together generates conflicts of interests triggered by opposing personal and social needs. Morality and law are aimed to offer solutions to solve or avoid these conflicts, providing two distinct sets of rules or norms for such a purpose. Whenever rules are broken and wrongdoing is established, punishment has to be provided and enforced to avoid future norm breaking. Wrong doing is prevented by coercion. Moral enforcement is a matter for divine or community reproach; therefore the degree of coercion of moral rules is low. In case of law, enforcement is the matter of Justice and Government. Justice is in charge of judging wrong doing and prescribing punishment, while Government is in charge of enforcing punishment [2] [3] .
Positive law is a man-made law that obliges or specifies an action establishing specific rights for an individual or group in order to prevent wrong doing that may cause damage for the individual or group [4] [5] . To create a positive law is, therefore, necessary to identify a damage avoided and define an action to prevent it. A good positive law is well justified by an important threat to the individual and/or group and proposes effective action to prevent this well identified menace (e.g. [4] ).
From time to time, tragedies like Newtown, CN shooting trigger waves of heated and emotional debates about some sort of gun control are basically polarized at two opposite ends of a broad spectrum: personal liberties versus safety and firearm injury. Proposals for gun control laws vary widely from country to country; while some countries try to restrict the sales of highly destructive weapons, other countries try to restrict the sales of all types of weapons. Within this second group, Brazil has recently carried out a referendum regarding the prohibition of firearm commerce (FC).
Media propaganda (Table 1) used arguments (PA) tried, either to convince voters about the treat to the individual and society of having a free firearm commerce (YA) in order to promote voting in favor (Yes) firearm commerce control, or to stress the inefficacy of this kind of control (NA) in order to promote voting against (No) it (see Table 1). If PA was believed and NA discredited, firearm commerce control would be considered as an efficient way to prevent the damage caused by firearm house accident, traffic dispute, mass killing, etc. On the contrary if NA was not believed or PA discredited, firearm control should be ruled out in the referendum. However, to this true, voters would have to consider that propaganda influenced their voting.
YA and NA have different ontological origins. YAs are based on empirical facts because any threat is about incurred damage (see Table 1). Because of this, YA analysis requires recalling information from episodic memory. In contrast, NAs are supported by reasoning about personal versus collective rights, causes of criminality, etc. (see Table 1). Because of this, NA analysis requires using neural circuits supporting analytical reasoning.
The possible influence of a propaganda argument on vote decision depends on the supposed social threat being considered as real and important in YA case, or the alleged proposition being considered as right and implying and important personal loss in NA case. The threat veracity (Table 1) demands its probability (p) being higher than zero, whereas its importance directly depends on p and the possible damage. Logical inference determines proposition veracity (Table 1) and the probability of failing to implement an adequate firearm control as well as the expected personal damage due to this failure is suggested to be high if the proposition is considered to be true. In this line of reasoning, highly probable believed YA would justify voting for firearm commerce control, while highly accepted NA would justify voting against the control. In order to influence vote decision, any PA has to justify one type of voting, but as each voter may have their own reasons (R) for voting Yes or No, even a justified PA may be of no influence if it does not match one of these reasons or it is not motivated by media propaganda to be included in R.
Rocha et al. [6] employed Principal Component Analysis (PCA) to study the amount of information 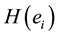 provided by electrode
provided by electrode  about vote intention one week prior to Election Day and showed that brain activity differed for “Yes” and “No” voters. “Yes” voters recruited neural circuits that have been previously described [7] in the Theory of Mind (ToM), which are involved in mentalizing third party intentions and important in the analyses of social issues. In contrast, “No” voters recruited neural circuits that have been shown to be involved
about vote intention one week prior to Election Day and showed that brain activity differed for “Yes” and “No” voters. “Yes” voters recruited neural circuits that have been previously described [7] in the Theory of Mind (ToM), which are involved in mentalizing third party intentions and important in the analyses of social issues. In contrast, “No” voters recruited neural circuits that have been shown to be involved

Table 1. Poll opinion questionnaire.
with self-awareness and self-related processing [7] -[10] .
In this context and from the above discussion, the following questions arise:
1) Are different neural circuits involved in YA and NA analysis because each type of argumentation has different ontological origins?
2) Is YA analysis mostly supported by episodic memory and social centered neural circuits and NA analysis mostly supported by analytical and self-related processing neural circuits?
In their study, Rocha et al. [6] asked 1136 people to complete a poll opinion questionnaire (PQ) to evaluate how true media propaganda arguments (PA) were, how much PA justified the referendum and how much the PA would influence their vote. In addition, 32 randomly selected individuals had their EEG recorded during their PQ responses. Here we used Loreta technology [11] and PCA analysis [6] to study 3 different EEG epochs of 2 seconds prior to the moments when the truth (T), justification (J) and influence (J) evaluations were decided, in order to test the hypotheses raised by questions 1 and 2 above.
2. Materials and Methods
2.1. Poll Opinion
A total of 1136 people living in the Great São Paulo area, a region inhabited by approximately 16 million people during the week that preceded the National Fire Arm Referendum, answered a poll opinion questionnaire (Table 1) about voting intention and propaganda arguments in favor (Y) and against (N) firearm commerce prohibition. Table 2 provides information on the sample population.
Table 2. Characteristics of the sample population.
The questionnaire shown in Table 2 was aimed to provide information about:
1) the intended vote (v) on the Brazilian Firearm Referendum;
2) a second voting opinion (v’) if the volunteers would change his/her opinion before the Election Day;
3) the truth of the propaganda arguments in favor of (Y) and against (N) firearm commerce prohibition;
4) if each argument justified (Y) or (N) the referendum; and
5) if each argument would influence (Y) or (N) the individual’s voting intention.
Note that, as we intended to study the correlation between arguments and voting intention, and not voting decision-making, we carried out the measurements one week before the moment of voting.
PCA analysis confirmed the distinction between Y and N arguments because responses to the poll questions were divided into distinct components, as shown in Table 3.
2.2. EEG Recordings
Sixteen females and 16 males (Table 2), all Caucasians and aged from 20 to 57 years old, were selected to have their EEG recorded (electrodes were placed according to the 10/20 system with an impedance of less than 10 kohm; low bandpass filter (50 Hz); a sampling rate of 256 Hz and 10 bit resolution, and ear lobe reference) while responding to the PQ. The EEG was visually inspected for artifacts prior to its processing, and the events associated with a poor EEG (e.g., when eye movements could compromise the results of the regression analysis) were discarded.
Two networked personal computers were used in the present study: one for the EEG recording and the other to sequentially display the different arguments in Table 1, where the individual’s evaluation of each argument’s truth (Yes or No) was requested. If it did (Yes) or did not (No) justify the referendum and if it would (Yes) or would not (No) influence the individual’s vote intention. The volunteers were allowed to take as much time as they needed to make their decisions as well as to indicate whether they were undecided about any of these questions. We recorded the following times, 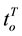 and
and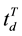 , as the time question when the argument truth was introduced and the time of individual’s decision, respectively;
, as the time question when the argument truth was introduced and the time of individual’s decision, respectively; 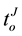 and
and 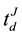 represented the time question about the referendum justification was introduced and the time of the individual’s decision, respectively, and
represented the time question about the referendum justification was introduced and the time of the individual’s decision, respectively, and 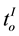 and
and 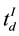 represented the time question about the argument voting influence was introduced and the time of the individual’s decision, respectively. The following EEG epochs were obtained for the analysis:
represented the time question about the argument voting influence was introduced and the time of the individual’s decision, respectively. The following EEG epochs were obtained for the analysis: 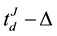
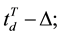 and
and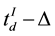 . The value of Δ was set as 2 seconds to ensure the volunteers were allowed sufficient time to reason and to make a decision that did not compromise our analysis.
. The value of Δ was set as 2 seconds to ensure the volunteers were allowed sufficient time to reason and to make a decision that did not compromise our analysis.
Low Resolution Electromagnetic Tomography (Loreta), which was developed by Pascual-Marqui [11] , was used to localize the potential EEG source generators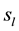 . The corresponding T, J and I EEG epochs were averaged for each electrode into different files according to each argument type (Y or N), which generated the EEG averaged files TY and TN; JY and JN; and IY and IN; respectively. Each of these files consisted of the corresponding EEG averages that were calculated for each of the 20 electrodes used to record the associated electrical activity for each decision epoch. A grand average was calculated for each of these files, and the corresponding z-score determined the EEG times that were statistically significant for the Loreta Analysis. Only those EEG moments with a z-score greater than 1.961 (5% significance level) were selected for this analysis (Figure 1). Only the areas showing the best match for each selected EEG moment was assumed as potential EEG source generators
. The corresponding T, J and I EEG epochs were averaged for each electrode into different files according to each argument type (Y or N), which generated the EEG averaged files TY and TN; JY and JN; and IY and IN; respectively. Each of these files consisted of the corresponding EEG averages that were calculated for each of the 20 electrodes used to record the associated electrical activity for each decision epoch. A grand average was calculated for each of these files, and the corresponding z-score determined the EEG times that were statistically significant for the Loreta Analysis. Only those EEG moments with a z-score greater than 1.961 (5% significance level) were selected for this analysis (Figure 1). Only the areas showing the best match for each selected EEG moment was assumed as potential EEG source generators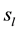 . Loreta analyses provided anatomical information and the Brodmann area number to localize the source of the EEG activity that was recorded at each selected time.
. Loreta analyses provided anatomical information and the Brodmann area number to localize the source of the EEG activity that was recorded at each selected time.
The amount of information  provided by electrode
provided by electrode  was calculated to summarize information by electrodes
was calculated to summarize information by electrodes  about the identified sources
about the identified sources . Because EEG data are assumed to be a weighted sum of the electrical activity of different sources, correlation analysis of the EEG activity
. Because EEG data are assumed to be a weighted sum of the electrical activity of different sources, correlation analysis of the EEG activity  recorded by the different electrodes
recorded by the different electrodes  may be used to calculate
may be used to calculate  in order to summarize information provided by each electrode
in order to summarize information provided by each electrode
Table 3. PCA analysis of poll opinion the questionnaire responses.

Figure 1. Grand average calculated for TY, JY and IY (left), JY and TN and IN (right). The red dots mark the EEG moments that were used for the Loreta calculations because their z-scores were above 1.961.
 about all involved sources
about all involved sources  into a single variable as proposed by Rocha et al. [6] [12] . The rationality is the following.
into a single variable as proposed by Rocha et al. [6] [12] . The rationality is the following.
The Pearson correlation R is + 1 in the case of a perfect positive (increasing) linear relationship (correlation), −1 in the case of a perfect decreasing (negative) linear relationship (anticorrelation), and some value between −1 and + 1 in all other cases, indicating the degree of linear dependence between the variables. As it approaches zero there is less of a relationship (closer to uncorrelated). The closer the coefficient is to either −1 or +1, the stronger the correlation between the variables. In this context, the correlation strength  is equal to
is equal to . If data
. If data ,
,  furnished by two electrodes
furnished by two electrodes ,
,  provide equivalent information about sources
provide equivalent information about sources  then Pearson correlation coefficient
then Pearson correlation coefficient  calculated for
calculated for ,
,  will approach
will approach , otherwise it will approach 0. The highest uncertainty about the information equivalence provided by
, otherwise it will approach 0. The highest uncertainty about the information equivalence provided by ,
,  occurs when the correlation strength
occurs when the correlation strength  approaches 0.5. Therefore, in the same line of reasoning used by Shannon to define the amount of information provided by a random variable, it is proposed that the informational equivalence,
approaches 0.5. Therefore, in the same line of reasoning used by Shannon to define the amount of information provided by a random variable, it is proposed that the informational equivalence,  of
of ,
,  furnished by
furnished by ,
,  is the expected value
is the expected value  of the information
of the information  provided by
provided by . Therefore:
. Therefore:
 (1)
(1)
Such that if  then
then  and if
and if  or
or  then
then .
.
Now, given
 (2)
(2)
the entropy of  is
is
 (3)
(3)
and it quantifies the mean informational equivalence from  concerning that provided by all other
concerning that provided by all other , because the different electrodes (information channels) provide different, but correlated, information about
, because the different electrodes (information channels) provide different, but correlated, information about .
.
In this context, we propose that
 (4)
(4)
quantifies the information provided by  about the sources
about the sources  involved in a cognitive task solving, because
involved in a cognitive task solving, because
a) if  for all
for all  then
then ,
,  for all
for all , and consequently
, and consequently . This indicates that
. This indicates that 
 does not provide any additional information about the sources
does not provide any additional information about the sources ;
;
b) for all other conditions  and quantifies the information provided by
and quantifies the information provided by  about the sources
about the sources .
.
Principal Component Analysis (PCA) is a statistical tool to investigate patterns of covariation in a large number of variables and to determine if information may be condensed into small sets of these variables called principal components [12] . This transformation is defined in such a way that the first principal component is the one that accounts for as much of the variability in the data as possible, and each succeeding component in turn explain the subsequent amount of variance possible under the constraint that it be orthogonal to (i.e., uncorrelated with) the preceding components. Factorial mappings are proposed to represent the activity of the neural circuits enrolled in a cognitive task because they condensed the information provided by the electrodes sampling this neural activity. In this ways, factorial analysis does not map brain areas activated by a cognitive task, but provides information to disclose the activity of circuits composed by neurons distributed on different areas of the brain recruited by the cognitive task, because  measures the amount of information provided by
measures the amount of information provided by  about spatial and temporal distribution of
about spatial and temporal distribution of . PCA was applied here to study the covariation of
. PCA was applied here to study the covariation of  during each of the selected epochs. This analysis revealed that three factors had accounted for more than 80% of the covariation in
during each of the selected epochs. This analysis revealed that three factors had accounted for more than 80% of the covariation in  and that the factors had eigenvalues that were greater than 1.4. The factorial brain maps were constructed to describe the results of the PCA using the procedures employed by Rocha et al. [1] [12] taking the loading values
and that the factors had eigenvalues that were greater than 1.4. The factorial brain maps were constructed to describe the results of the PCA using the procedures employed by Rocha et al. [1] [12] taking the loading values  of
of  for each of the factors
for each of the factors  into account. To estimate the potential similarities between each factorial brain mapping that was associated to each
into account. To estimate the potential similarities between each factorial brain mapping that was associated to each  for each EEG time epoch, the authors used Pearson correlation coefficients, which were calculated for their respective loading values
for each EEG time epoch, the authors used Pearson correlation coefficients, which were calculated for their respective loading values .
.
3. Results
Poll opinion and EEG data analysis are described below.
3.1. Poll Opinion
The results obtained from the PQ analysis are shown in Table 4. All of the voters considered half of the “Yes” arguments (YA) and 76% of the “No” arguments (NA) as true. The PA was assumed to not justify the FC by more than 75% of the individuals, and more than 64% of responders declared that this would not influence their vote. The data did not significantly change if the Yes and No voters were considered separately. These results showed that people were more convinced of the NA and coherently assumed that the PA did not justify the FC. The majority of voters declared that their voting decision would be determined by other reasons than those proposed by the PA.
3.2. EEG Results
PCA and Loreta analyses for each EEG epoch and the comparison of the results obtained with each technique are discusses bellow.
3.2.1. Truth Evaluation
Grand averages (Figure 1) were clearly different for YA and NA, having 40 significant moments (Z score greater than 1.961) in NA case and 70 of these moments in YA case. These differences resulted in two distinct LS sets of identified Loreta sources associated with Truth evaluation of each type of argument (Figure 2). Because of this PCA mappings associated with Truth evaluation were clearly different for YA and NA (Figure 2).
Loreta sources were mostly identified at BA 10, 11, 17, 18, 29 and 47 for both types of argument but the frequency these sources were located at these locations differed for NA compared to YA.
PCA disclosed the existence of 3 patterns of  covariation that explained more than 60% of data covariance. P1 pattern consisted of F7, T3 and P4 in the YA case and PZ and F3 in the NA case. P2 pattern mostly consisted of the responses obtained by the right frontal electrodes in the YA case and the left frontal electrodes in the NA case. P3pattern consisted of F3, C3, P3, O1, FZ, CZ, PZ, OZ, C4 and O2 in the YA case and the FZ, C3, C4, P4, PZ, OZ and O2 electrodes in the NA case.
covariation that explained more than 60% of data covariance. P1 pattern consisted of F7, T3 and P4 in the YA case and PZ and F3 in the NA case. P2 pattern mostly consisted of the responses obtained by the right frontal electrodes in the YA case and the left frontal electrodes in the NA case. P3pattern consisted of F3, C3, P3, O1, FZ, CZ, PZ, OZ, C4 and O2 in the YA case and the FZ, C3, C4, P4, PZ, OZ and O2 electrodes in the NA case.
Pearson’s determination coefficient is used to calculate . This means that
. This means that  measures the information provided by each
measures the information provided by each  about the sets of neurons activated by the cognitive task being processed. Loreta analysis identifies these sets of activated neurons and PCA discloses those electrodes providing similar information about them. Since the electrical activity recorded by each
about the sets of neurons activated by the cognitive task being processed. Loreta analysis identifies these sets of activated neurons and PCA discloses those electrodes providing similar information about them. Since the electrical activity recorded by each  depends on the distance from the electrode to the activated neural sources
depends on the distance from the electrode to the activated neural sources , the nearest
, the nearest  to each electrode that significantly loaded in each PCA pat- tern are the most probable set of neurons contributing to distinguish these patterns. Figure 3 display the XY coordinates of nearest
to each electrode that significantly loaded in each PCA pat- tern are the most probable set of neurons contributing to distinguish these patterns. Figure 3 display the XY coordinates of nearest  to each PCA pattern in the case of Truth evaluation and Table 5 identify these sources.
to each PCA pattern in the case of Truth evaluation and Table 5 identify these sources.
Table 4. Percentage of the different responses to the poll questions.
Yes voters―people declaring an intention to vote Yes. No voters―people declaring an intention to vote No. The data in red represents a significant difference when the Yes and No arguments were compared.

Figure 2. Truth evaluation: the most frequent locations of Loreta sources identified for NA and YA (upper graph) and PCA mappings associated with these identified sources.

Figure 3. PCA mappings and spatial location of those Loreta sources associated with each PCA pattern in case of truth evaluation. The sources are those described in Table 5.
Loreta analysis and PCA comparison shows that pattern P1 is correlated with  located at BA 6, 9 and 10 in NA case and BA 7, 22, 40 and 47 in YA case. In addition, sources associated with P2 and located at Middle Frontal Gyrus (BA 9, 10 and 11), Superior Frontal Gyrus (BA 9, 10 and 11), Medial Frontal Gyrus (BA 10) and Inferior Frontal Gyrus (BA 47) are common to both NA and YA; whereas those located at Precuneus (BA 7); Superior Frontal Gyrus (BA 8), Supramarginal Gyrus (BA 40) and Middle Frontal Gyrus (BA 46) were specific to NA, and those located at Medial Frontal Gyrus (BA 9 and 11), Precentral Gyrus (BA 44) and Inferior Frontal Gyrus (BA 45) were specific to YA. In case of pattern P3, Postcentral Gyrus (BA 7), Superior Parietal Lobe (BA 7), Cuneus (BA 18 and 19) and Middle Occipital Gyrus (BA 18 and 19) were the only common source locations for NA and YA, all other sources are different for each type of argument.
located at BA 6, 9 and 10 in NA case and BA 7, 22, 40 and 47 in YA case. In addition, sources associated with P2 and located at Middle Frontal Gyrus (BA 9, 10 and 11), Superior Frontal Gyrus (BA 9, 10 and 11), Medial Frontal Gyrus (BA 10) and Inferior Frontal Gyrus (BA 47) are common to both NA and YA; whereas those located at Precuneus (BA 7); Superior Frontal Gyrus (BA 8), Supramarginal Gyrus (BA 40) and Middle Frontal Gyrus (BA 46) were specific to NA, and those located at Medial Frontal Gyrus (BA 9 and 11), Precentral Gyrus (BA 44) and Inferior Frontal Gyrus (BA 45) were specific to YA. In case of pattern P3, Postcentral Gyrus (BA 7), Superior Parietal Lobe (BA 7), Cuneus (BA 18 and 19) and Middle Occipital Gyrus (BA 18 and 19) were the only common source locations for NA and YA, all other sources are different for each type of argument.
Table 5. Loreta sources associated with each PCA pattern: Truth Identification.
Grey background highlight common source locations.
3.2.2. Justification Evaluation
Grand averages (Figure 2) were clearly different for YA and NA, having 50 significant moments in NA case and 80 of these moments in YA case. These differences resulted in two distinct LS sets of identified Loreta sources associated with Truth evaluation of each type of argument (Figure 2). Because of this PCA mappings associated with Truth evaluation were clearly different for YA and NA (Figure 4).

Figure 4. Justification evaluation: the most frequent locations of Loreta sources identified for NA and YA (upper graph) and PCA mappings (lower graph) associated with these identified sources. PCA mappings showing pattern P1 in green and to dark blue; pattern P2 in yellow and brown, and pattern P3 in rose to dark red.
Loreta sources were mostly identified at BA 10, 11, 17, 18 and 47 for both types of argument but the frequency these sources were located at these locations differed for NA compared to YA.
PCA mappings associated with Justification evaluation were clearly different for YA and NA (Figure 4). The P1 pattern was reduced in electrode P4 for the YA case and in the occipital electrodes for the NA case. Pattern P2 was very similar for both YA and NA, consisting of bilateral frontal electrodes. Pattern 3 consisted of F3, C3, P3, O1, FZ, CZ, PZ, OZ, C4 and O2 responses in the YA case, and comprised the F3, C3, P3, CZ, PZ and C4 electrodes.
Loreta analysis and PCA comparison (Figure 5 and Table 6) shows that pattern P1 is correlated with  lo-
lo-

Figure 5. PCA mappings and spatial location of those Loreta sources associated with each PCA pattern in case of Justification evaluation. The sources are those described in Table 6.
Table 6. Loreta sources associated with each PCA pattern: Justification.
cated at BA 17, 18 and 19 in NA case and BA 7 and 22 in YA case. In addition, sources associated with P2 and located at Middle Frontal Gyrus (BA 10, 11 and 46), Superior Frontal Gyrus (BA 8, 9, 10 and 11), Medial Frontal Gyrus (BA 10 and 11), Supramarginal Gyrus (BA 40) and Inferior Frontal Gyrus (BA 47) are common to both NA and YA; whereas those located at Rectal Gyrus (BA 11) and Inferior Parietal Gyrus (BA 40) were specific to NA, and those located at Orbital Gyrus (BA 11 and 47), Precentral Gyrus (BA 44) and Inferior Frontal Gyrus (BA 45) were specific to YA. In case of pattern P3, Postcentral Gyrus (BA 7) and Superior Frontal Gyrys (BA 6 and 9), were the only common source locations for NA and YA, all other sources are different for each type of argument.
3.2.3. Influence Evaluation
Grand averages (Figure 1) were clearly different for YA and NA, having 35 significant moments in NA case and 56 of these moments in YA case. These differences resulted in two distinct LS sets of identified Loreta sources associated with Truth evaluation of each type of argument (Figure 6). Because of this PCA mappings

Figure 6. Influence evaluation: the most frequent locations of Loreta sources identified for NA and YA (upper graph) and PCA mappings (lower graph) associated with these identified sources.
associated with Truth evaluation were clearly different for YA and NA (Figure 2).
Loreta sources were mostly identified at BA 7, 10, 11, 18 and 21 for both types of argument but the frequency these sources were located at these locations differed for NA compared to YA.
PCA mappings associated with Justification evaluation were clearly different for YA and NA (Figure 6). Pattern P1 consisted of F3, T5 and T6 in the YA case and occiptial electrodes in the NA case. Pattern P2 was very similar for both YA and NA, consisting of bilateral frontal electrodes. Pattern 3 consisted of P3, O1, FZ, CZ, PZ, OZ, C4, P4 and O2 in the YA case and the F3, C3, P3, CZ, PZ and C4 electrodes.
Loreta analysis and PCA comparison (Figure 7 and Table 7) shows that pattern P1 is correlated with  mostly located at BA 18 in NA case and BA 21 and 22 in YA case. In addition, sources associated with P2 and located at Middle Frontal Gyrus (BA 10 and 11), Medial Frontal Gyrus (BA 10 and 11)) and Superior Frontal Gyrus (BA 8, 10 and 11) are common to both NA and YA; whereas those located at Middle Frontal Gyrus (BA 8 and 47) and Rectal Gyrus (BA 11, and those located at Inferior Frontal Gyrus (BA 10), Middle Frontal Gyrus (BA 46), Orbital Gyrus (BA 47), Precentral Gyrus (BA 44) and were specific to YA. In case of pattern P3, Postcentral Gyrus (BA 5 and 7), Superior Frontal Gyrys (BA 6), Inferior Temporal Gyrus (BA 21) and Superior Temporal Gyrus (BA 22) were the only common source locations for NA and YA, all other sources are different for each type of argument.
mostly located at BA 18 in NA case and BA 21 and 22 in YA case. In addition, sources associated with P2 and located at Middle Frontal Gyrus (BA 10 and 11), Medial Frontal Gyrus (BA 10 and 11)) and Superior Frontal Gyrus (BA 8, 10 and 11) are common to both NA and YA; whereas those located at Middle Frontal Gyrus (BA 8 and 47) and Rectal Gyrus (BA 11, and those located at Inferior Frontal Gyrus (BA 10), Middle Frontal Gyrus (BA 46), Orbital Gyrus (BA 47), Precentral Gyrus (BA 44) and were specific to YA. In case of pattern P3, Postcentral Gyrus (BA 5 and 7), Superior Frontal Gyrys (BA 6), Inferior Temporal Gyrus (BA 21) and Superior Temporal Gyrus (BA 22) were the only common source locations for NA and YA, all other sources are different for each type of argument.
4. Discussion
A total of 1136 people living in the Great São Paulo area were asked whether media propaganda would influence or not their vote decision in the Brazilian Firearm Commerce Referendum. As discussed in the introduction, in order to justify a given type o vote, an argument has to be true and high probable. Each voter has their own set of arguments or reasons R for voting for (Yes) or against (No) firearm commerce control that may be dependent on many causes and believes. Therefore, it does not necessarily follow that a justified media argument will influence vote decision. In this line of reasoning, any media campaign is aimed to manipulate the inclusion of as many justified arguments pro a vote decision as possible in R.
Poll opinion results showed that arguments (YA) for voting Yes was considered true by 50% of the volunteers and while 76% of them agreed with arguments (NA) for voting No, no matter their declared voting intention. Despite this, only 20% of the volunteers considered that media arguments would justify the intended vote. As discussed before, besides being true, an argument has to be high probable in order to justify the intended vote decision. Therefore, it may be said that media campaign failed in proving their arguments to probable enough to justify voting for or against firearm control. In line with these results, more than 60% of the volunteers declared that media propaganda would not influence their vote decision. Because for most of people, media arguments were not justified, media propaganda also failed in changing voter’s set of reasons R. This was a remarkable finding because it shows that the vote decision was guided by reasons other than those that are usually discussed in a debate when any form of gun control gains national attention. The analysis of the EEGs recorded during the PQ responses shed some light on this subject.
Both Loreta analysis and PCA show that the same individual recruited different neural circuits during T, J and I epochs to analyze YA and NA irrespective of their vote intention about firearm commerce prohibition. YA analysis recruited larger set of neurons than those recruited by NA, indicating, perhaps that YA evaluation requires a reasoning more complex than that required by NA.
The core of YA arguments is an empirical fact that would characterize a social threat to be avoided by the firearm control. Voters would have to recall these facts from memory in order to evaluate their probability p and associated damage. If p > 0 then the argument is true and if p > 0 then it is frequent. Now, if probability and damage are high an important social threat is identified. In contrast, the core of NA arguments implies reasoning about personal versus collective rights, causes of criminality, etc. In this case, it is necessary first to establish if the argument true and then its expected personal benefit and risk have to be estimated. Now, if argument is true, expected personal benefit is low and risk is high a strong case is made for No voting. Because of this, it is expected that neural circuits involved in YA and NA analysis have to be different as observed here. This is in line with other propositions in the literature, e.g., that of “exploration-explotation” dilemma by Nathaniel et al. [13] .
It is proposed here that PCA pattern P3 it is associated with the memory recalling of the fact proposed by YA, because this pattern is associated with sources located at BA 9 and 10 in the frontal cortex [14] -[19] , BA 18 and 19 in the posterior cortex [20] and BA 38 and 40 at the temporal cortex [14] [20] [21] . In addition, pattern P2 it

Figure 7. PCA mappings and spatial location of those Loreta sources associated with each PCA pattern in case of Influence evaluation. The sources are those described in Table 7.
Table 7. Loreta sources associated with each PCA pattern: Influence.
is associated mostly with sources located at the right frontal cortex (mostly BA 9, 10, 11 and 47) that has an important role in episodic memory handling as discussed by Henson et al. [22] and Lepage et al. [16] .
Complementing the above hypothesis, it is assumed here that PCA pattern P3 it is associated with the analytical reasoning involved in establishing the veracity of the fact proposed by NA, because this pattern is associated with sources located at BA 11 in the frontal cortex, BA 3 and 5 in the parietal cortex and BA 17 and 18 in the posterior cortex [22] -[25] . In addition, pattern P2 it is associated mostly with sources located at the left frontal cortex (mostly BA 9, 10, 11, 46 and 47) that has in important role in analytical reasoning as proposed by Canessa et al. [22] , Knauff et al. [23] and Mackey et al. [25] .
Finally, it must be stressed that P1 is the largest and most distinctive PCA pattern associate with truth evaluation of YA and NA as it was the most distinctive pattern between Yes and No voters [6] .
From the above, it is proposed here that pattern P2 disclosed those frontal executive neural circuits handling temporal-parietal-occipital circuits disclosed by pattern P3 and in charge of data recalling in case of episodic memory and YA analysis and in charge of logically inferring the truth of NA. Rocha et al. [6] also considered P2 as associated with executive frontal neural circuits handling those other neural circuits disclosed by P3 and in charge of differentiating Yes from No voters.
YA arguments were socially driven, which was typical for arguments in favor of any gun control in a variety of societies. ToM refers to the ability to attribute mental states to others and the ability to infer the emotional experiences of others (empathy), which are important processes in social cognition. Brain imaging studies in healthy subjects have described a brain system involving the medial prefrontal cortex, inferior frontal gyrus (IFG), superior temporal sulcus and temporal pole in ToM processing [7] [26] . In addition, the role of IFG in ToM circuits has been thought to block self-centered reasoning [8] [10] [27] [28] . YA sources were located at all these areas (Figure 2 and Figure 3) and while IFG located at BA 47 is common to both YA and NA analysis, IFG located at BA 45 is part of P2 in YA case but not in NA case. Taken together, these results corroborate the hypothesis that socially driven neural circuits are important for establishing the veracity of YA.
In contrast with the above findings, NA arguments were self-interest driven, which is typical of arguments against any gun control in a variety of societies. Humans know others as outside speakers, visual images and recognized actors stored in our retrospective memory. Our own selves are identified by inner speech, somesthetic imagery, actions and episodic/autobiographic memories. The superior frontal and lingual gyrus have been reported [24] [29] -[32] as important structures associated with self-intended actions. The precuneus is a structure considered as a hub connecting the parietal and prefrontal regions [33] [34] , and thereby plays an important role in various functions, including retrospective, episodic and autobiographic memories [7] and self reasoning [7] [10] [27] . Many of the sources discriminating NA from YA are located in the superior frontal, fusiform and lingual regions as well as in the precuneus, supporting the hypothesis that self-related circuits are important in establishing NA truth. The precuneus is also among the structures where YA discriminating sources were located; however, these sources were located in Brodmann area 7 and 19 in the YA and NA cases, respectively. Furthermore, in the rectal gyrus, which is one of the regions where the NA discriminating sources were located, the authors proposed that it is involved in evaluating the degree of NA truth. These results corroborated the hypothesis that self-interest driven neural circuits are important for establishing the veracity of NA.
Patterns P2 and P3 dramatically changed from truth to justification evaluation in NA case. P2 and P3 are associated to sources located predominantly at the left hemisphere during truth evaluation and bilaterally located in case of justification analysis. In this case, P3 is also associated to sources located at more anterior places in comparison to truth evaluation. All patterns were similar during justification and influence evaluation for NA. As discussed above truth evaluation of NA implies logical inference whereas justification analysis implies to estimate expected benefits and risks. Because of this, neural circuits supporting each type of analysis to be different as observed here. In addition, NA were believed by more than 75% volunteers but 75% of then considered that they did not justify voting No. Therefore, it may be supposed that expected risks were considered greater than benefits. Because NAs were considered not justified they would not influence vote decision. Because risks were supposed greater than benefits NAs would not influence vote decision. This may explain because brain activity was similar during both J and I epochs.
Pattern P3 did not change from truth to justification evaluation in YA case but P2 was associated to sources located predominantly at the left hemisphere during truth evaluation and bilaterally located in case of justification analysis. In addition, pattern P2 did not change from justification to influence evaluation in YA case but P3 was associated with sources located at a small number of locations during influence than justification evaluation. To be considered as justified, YAs have to be true and frequent. Around 50% of the volunteers considered them as true, but less than 30% of them considered that arguments would justify vote. These means that most of volunteers did not considered the appointed fact as frequent or associated with an important social damage. But these evaluations are also of empirical nature. To be true the probability of the proposed fact has to be greater than 0, and to be considered frequent it must be much greater than 0. So, the same circuits must be involved in both analyses. Because YAs were considered not justified they would not influence vote decision as observed here. This may explain why because brain activity may involve a smaller number of areas during I epoch in comparison to J epoch.
5. Conclusion
In conclusion, it may be said that our results show that different neural circuits support YA and NA analyses because these analyses enrolled neurons located at different cortical areas. YA analysis was associated with activation of cortical areas known to be involved in retrospective and episodic memory as well as with areas known to be involved in evaluating others’ intention. In contrast, NA analysis was associated with cortical areas that are proposed to be involved with self-interest evaluation. Moreover, information about neural mechanism involved in voting decision-making obtained in this work helped to differentiate distinct ontological origins of YA and NA.
Acknowledgements
This work was partially supported by LIM01-HCFMUSP, CNpQ and FAPESP.
Ethical Aspects
All participants signed an informed consent. This work has been approved by the Ethics Committee of the School of Medicine of the University of Sao Paulo.
References
- Rocha, A.F. (2013) Toward a Better Understanding of the Relationship between Neurosciences and Law. Law & Neuroscience Journal, 4. http://papers.ssrn.com/abstract=2313182
- Hobbes, T. (1651) Leviathan. Penguin Classics, Penguin Books, New York.
- Rousseau, J.J. (1968) Du Contrat Social: Texte Original. Penguin Books Limited, London.
- Hart, H.L.A. (1961) Concept of Law.
- Kelsen, H. (1960) Reine Rechteslehre. Verlag Fraz Deuticke, Viena.
- Rocha, A.F., Theoto, F., Burattini, M.N. and Massad, E. (2010) Neurodynamics of an Election. Brain Research, 1351, 198-211. http://dx.doi.org/10.1016/j.brainres.2010.06.046
- Frith, C.D. and Frith, U. (2006) The Neural Basis of Mentalizing. Neuron, 50, 531-534. http://dx.doi.org/10.1016/j.neuron.2006.05.001
- Cavanna, A.R. and Trimble, M.R. (2006) The Precuneus: A Review of Its Functional Anatomy and Behavioral Correlates. Brain, 129, 564-583. http://dx.doi.org/10.1093/brain/awl004
- Hartwright, C.E., Apperly, I.A. and Hansen, P.C. (2006) Multiple Roles for Executive Control in Belief-Desire Reasoning: Distinct Neural Networks Are Recruited for Self Perspective Inhibition and Complexity of Reasoning. NeuroImage, 61, 921-930. http://dx.doi.org/10.1016/j.neuroimage.2012.03.012
- Van den Ouden, H.E.M., Frith, U., Frith, C. and Blakemore, S.-J. (2005) Thinking about Intentions. NeuroImage, 28, 787-796. http://dx.doi.org/10.1016/j.neuroimage.2005.05.001
- Pasqual-Marqui, R.D., Esslen, M., Kochi, K. and Lehmann, D. (2002) Functional Imaging with Low-Resolution Brain Electromagnetic Tomography (LORETA): A Review. Methods and Findings in Experimental and Clinical Pharmacology, 24, 91-95.
- Rocha, F.T., Rocha, A.F., Massad, E. and Menezes, R.X. (2005) Brain Mappings of the Arithmetic Processing in Children and Adults. Cognitive Brain Research, 22, 359-372. http://dx.doi.org/10.1016/j.cogbrainres.2004.09.008
- Daw, N.D., O’Doherty, J.P., Dayan, P., Seymor, B. and Dolan, R.J. (2006) Cortical Substrates for Exploratory Decisions in Humans. Nature, 441, 876-879. http://dx.doi.org/10.1038/nature04766
- Addis, D.R. and Schacter, D.L. (2008) Constructive Episodic Simulation: Temporal Distance and Detail of Past and Future Events Modulate Hippocampal Engagement. Hippocampus, 18, 227-237. http://dx.doi.org/10.1002/hipo.20405
- Cabeza, R., Dolcos, F., Graham, R. and Nyberg, L. (2002) Similarities and Differences in the Neural Correlates of Episodic Memory Retrieval and Working Memory. Neuroimagen, 16, 317-330.
- Lepage, M., Ghaffar, O., Nyberg, L. and Tulving, E. (2000) Prefrontal Cortex and Episodic Memory Retrieval Mode Proceedings of the National Academy of Sciences of the United States of America, 97, 506-511. http://dx.doi.org/10.1073/pnas.97.1.506
- Platel, H., Baron, J.-C., Desgranges, B., Bernard, F. and Eustache, F. (2003) Semantic and Episodic Memory of Music Are Subserved by Distinct Neural Networks. NeuroImage, 20, 244-256. http://dx.doi.org/10.1016/S1053-8119(03)00287-8
- Ranganath, C., Johnson, M.K. and D’Ésposito, M. (2003) Prefrontal Activity Associated with Working Memory and Episodic Long-Term Memory. Neuropsychologia, 41, 378-389. http://dx.doi.org/10.1016/S0028-3932(02)00169-0
- Rugg, M.D., Fletcher, P.C., Frith, C.D., Frackowiak, R.S.J. and Dolan, R.J. (1996) Differential Activation of the Prefrontal Cortex in Successful and Unsuccessful Memory Retrieval. Brain, 119, 2073-2083. http://dx.doi.org/10.1093/brain/119.6.2073
- Baron, J.C. and Jones, T. (2012) Oxygen Metabolism, Oxygen Extraction and Positron Emission Tomography: Historical Perspective and Impact on Basic and Clinical Neuroscience. Neuroimage, 61, 492-504. http://dx.doi.org/10.1016/j.neuroimage.2011.12.036
- Scheeringa, R., Petersson, K.M., Oostenveld, R., Norris, D.G., Hagoort, P. and Bastiaansen, M.C.M. (2009) Trial-by-Trial Coupling between EEG and BOLD Identifies Networks Related to Alpha and Theta EEG Power Increases during Working Memory Maintenance NeuroImage, 44, 1224-1238. http://dx.doi.org/10.1016/j.neuroimage.2008.08.041
- Canessa, N., Gorini, A., Cappa, S.F., Piattelli-Palmarini, M., Danna, M., Fazio, F. and Perani, D. (2005) The Effect of Social Content on Deductive Reasoning: An fMRI Study. Human Brain Mapping, 26, 30-43. http://dx.doi.org/10.1002/hbm.20114
- Henson, R.N.A., Shallice, T. and Dolan, R.J. (1999) Right Prefrontal Cortex and Episodic Memoryretrieval: A Functional MRI Test of the Monitoring Hypothesis. Brain, 122, 1367-1381. http://dx.doi.org/10.1093/brain/122.7.1367
- Knauff, K., Mulack, T., Kassubek, J., Salih, H.R. and Greenlee, M.W. (2002) Spatial Imagery in Deductive Reasoning: A Functional MRI Study. Cognitive Brain Research, 13, 203-212. http://dx.doi.org/10.1016/S0926-6410(01)00116-1
- Goel, V. and Dolan, R.J. (2001) Functional Neuroanatomy of Three-Term Relational Reasoning. Neuropsychologia, 39, 901-909. http://dx.doi.org/10.1016/S0028-3932(01)00024-0
- Mackey, A.P., Miller Singley, A.T. and Bunge, S.A. (2013) Intensive Reasoning Training Alters Patterns of Brain Connectivity at Rest. Journal of Neuroscience, 33, 4796-4803.
- Völlm, B.A., Taylor, A.N.W., Richardson, P., Corcoran, R., Stirling, J., McKie, S., et al. (2006) Neuronal Correlates of Theory of Mind and Empathy: A Functional Magnetic Resonance Imaging Study in a Nonverbal Task. NeuroImage, 29, 90-98. http://dx.doi.org/10.1016/j.neuroimage.2005.07.022
- Lundstrom, B.N., Ingvar, M. and Petersson, K.M. (2005) The Role of Percuneus and Frontal Cortex during Source Memory Episodic Retrieval. NeuroImage, 27, 824-834. http://dx.doi.org/10.1016/j.neuroimage.2005.05.008
- Van der Meer, L., Groenewold, N.A., Nolen, W.A., Pijnenborg, M. and Aleman, A. (2011) Inhibit Yourself and Understand the Other: Neural Basis of Distinct Processes Underlying Theory of Mind. NeuroImage, 56, 2364-2374. http://dx.doi.org/10.1016/j.neuroimage.2011.03.053
- Martino, J., Gabarrós, A., Deus, J., Juncadella, M., Acebes, J.J., Torres, A. and Pujol, J. (2011) Intrasurgical Mapping of Complex Motor Function in the Superior Frontal Gyrus. Neuroscience, 179, 131-142. http://dx.doi.org/10.1016/j.neuroscience.2011.01.047
- Nakagawa, S., Nishiike, S., Tonoike, M., Takeda, N. and Kubo, T. (2002) Measurements of Brain Magnetic Fields Associated with Apparent Self-Motion. International Congress Series, 1232, 367-371. http://dx.doi.org/10.1016/S0531-5131(01)00790-7
- Samson, D., Apperly, I.A., Chiavarino, C. and Humphreys, G.W. (2004) Left Tempoparietal Junction Is Necessary for Representing Someone Else’s Belief. Nature Neuroscience, 7, 499-500. http://dx.doi.org/10.1038/nn1223
- Sulpizio, V., Committeri, G., Lambrey, S., Berthoz, A. and Galati, G. (2013) Selective Role of Lingual/Parahippo- campal Gyrus and Retrosplenial Complex in Spatial Memory across Viewpoint Changes Relative to the Environmental Reference Frame. Behavioral Brain Research, 242, 62-75. http://dx.doi.org/10.1016/j.bbr.2012.12.031
- Bullmore, E. and Sporns, O. (2009) Complex Brain Networks: Graph Theoretical Analysis of Structural and Functional Systems. Nature Reviews Neuroscience, 10, 186-198. http://dx.doi.org/10.1038/nrn2575
NOTES

*Corresponding author.