American Journal of Plant Sciences
Vol.4 No.6A(2013), Article ID:33294,7 pages DOI:10.4236/ajps.2013.46A003
High Frequency Sporophytes Regeneration from the Spore Culture of the Endangered Aquatic Fern Isoetes coreana
![]()
1Biological Resources Center, Korea Research Institute of Bioscience and Biotechnology (KRIBB), Daejeon, South Korea; 2Department of Biological Science, Ajou University, Suwon, South Korea; 3Department of Life Science, Soonchunhyang University, Asan, South Korea; 4Green Bio Research Center, Korea Research Institute of Bioscience and Biotechnology (KRIBB), Daejeon, South Korea.
Email: *hkchoi@ajou.ac.kr, *kimsw@kribb.re.kr
Copyright © 2013 Myung Jin Oh et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received April 1st, 2013; revised May 2nd, 2013; accepted May 31st, 2013
Keywords: Aquatic Fern; Isoetes coreana; Spore Culture; Sporophyte Regeneration
ABSTRACT
Using a mixed culture of megaspores and microspores from I. coreana, we established high frequency sporophyte regeneration system. After 20 days of culturing in MS basal medium, microscopic examination showed significant morphological changes and the microspore released numerous small vesicles into the culture medium. Megaspores also showed dramatic morphological changes during its incubation time in culture. The spore wall was cracked by the expansion of the megaspore (about 2 times increase in diameter). Simultaneously, brown spots were observed on the surface of the megaspores. The frequency of female gametophytes developing from immature megaspores cultured in MS basal liquid medium (pH 7) supplemented with 1 mgl−1 GA3 was 46%. However, these female gametophytes derived from megaspore only culture could not differentiate into sporophytes. The mixed culture of microspores and megaspores resulted in successful sporophyte regeneration. The highest frequency (12.3%) of green sporophyte regeneration from mixed spore culture occurred when the cultures were maintained at 25˚C under cool-white fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod. Regenerated sporophytes were transferred to a test tube containing vermiculite and a sand mixture and left there until they had three leaves. After root growth and the fifth leaf had emerged, more than 95% of the regenerated sporophytes were successfully transferred to the soil and grown to mature plants. The sporophyte regeneration system established in this study could be successfully used for the restoration of the endangered aquatic species, I. coreana.
1. Introduction
Isoëtes L., known as the quillworts, is a cosmopolitan genus of lycopsids that consists of approximately 150 or more species found in lake, wetland (swamp and marsh) and terrestrial habitats. [1,2]. A limited number of aquatic plants in this genus have Crassulacean Acid Metabolism (CAM) which makes the aquatic plants especially valuable for research into this unique metabolic pathway [3]. In addition, Isoëtes is a representative of the heterosporous aquatic fern. The microspores are produced in large numbers (approximately 130,000 to 1,000,000) in each microsporangium and the megaspores are usually 50 to 300 in each megasporganium [4]. Megaspores develop into the female gametophytes and the eggs are produced in the archegonia. Microspores develop into the male gametophytes and sperms are released in the antheridia. The flagellated sperm needs water in order to swim to the egg. Sperm and egg are fused to form a zygote and then these zygotes are developed into young sporophytes [5,6]. According to Caplen and Werth [7], “information on the natural history of sexual reproduction in Isoëtes is completely lacking”, but direct observations of sexual reproduction in this genus have been made in vitro but not under different culture conditions [8,9]. Therefore, the development of an in vitro culture system for Isoëtes is required for the study of its life cycles. Studies of tissue cultures in ferns have been utilized as a research instrument for the mass proliferation of ornamental ferns
[10]. The first success in the field of intensive multiplication of plants through in vitro techniques was cited around 1970. The Boston fern (Nephrolepis exaltata cv. Bostoniensis) was the first plant micropropagated in vitro for a commercial purpose [11]. Several studies for obtaining female gametophytes, male gametophytes, and sporeling of Isoëtes species have been done [8,9,12,13]. For example, Koot and Britton [13] investigated the experimental germinations of microspores and megaspores from five species (i.e., I. macrospora, I. tuckermanii, I. riparia, I. acadiensis, and I. echinospora) from North America in small vials with distilled water. However, there are no successful reports about in vitro culture systems for the mass proliferation of Isoëtes species.
Currently, I. coreana is restricted to South Korea [14,15]. Based on our recent field surveys, the habitats of I. coreana have degraded and the plant has disappeared from several locations [15]. Therefore, this species is now considered to be rare and endangered by the Korean government. Recently, calls have been made for urgent conservation strategies to avert its impending extirpation from the mainland of South Korea. Furthermore, it is necessary to restore as many individuals as possible propagated by in vitro sexual fusion [15]. However there is no information on the reproduction and life cycle of I. coreana [14].
This study reports on spore germination and early gametophyte and sporophyte formation of I. coreana in response to changes in pH, temperature, GA3, and light intensity.
2. Materials and Methods
2.1. Spore Collection and Sterilization
Spores of I. coreana were obtained from sporophytes growing in Hwacheon (Kangwon, South Korea). The plants were transferred and grown on a mixture of sandy soil and vermiculite (1:1 v/v). Each leaf broadens to a swollen base up to 5 mm wide where they attach in corm-like base. There is a sac that contains the spores at the bottom of each leaf base. The sacs at the bottom of leaf create two types of spores, female (megaspores, about 0.4 mm diameter) and male (microspores, a few micrometres in diameter). These sacs (sporangia) were carefully detached from the leaf base and classified into microsporangium and megasporangium, respectively.
These collected sporangia were soaked in water for 1h, and then sterilized for 1 min with 70% ethanol. After rinsing the sporangia with sterile distilled water, they were sterilized again for 7 min with a solution of NaClO (1%) containing Tween-20 (0.01%). Subsequently, they were rinsed three times with sterile distilled water and centrifuged at 2000 rpm for 3 min between rinses. After final rinse, sporangia were soaked in sterile distilled water. Sporangia were ruptured by pipette and the spores were released and resuspended in full-strength Murashige and Skoog (MS) [16] liquid basal medium. Spores were transferred to culture media supplemented with or without GA3 and placed under different culture conditions such as pH and light intensity.
2.2. Effect of the Developmental Stage of the Megaspore and Culture Conditions (pH, Temperature, and GA3) on Female Gametophyte Formation
To examine the effect of the developmental stage of the megaspore on female gametophyte formation, white immature and light-brown mature megaspores were cultured in MS basal liquid medium and distilled water respectively. After two weeks in the culture medium, the total number of megaspores initially incubated was examined. After another two weeks in the culture medium, the total number of megaspores showing expansion with brown spots was counted.
The natural habitat of habitats of Isoëtes species is mainly lake and swamp. Therefore, we also examined the effect of pH of the culture medium on female gametophyte formation. The pH of culture medium was adjusted to 5, 5.8, 6.4 and 7 with 1N NaOH. Also, the pH of the distilled water medium was 6.2. After four weeks of incubation in the culture medium, the frequency of female gametophyte formation was examined.
To examine the effect of culture temperature on female gametophyte formation, megaspores were cultured in MS basal medium and sterile distilled water at 18˚C and 25˚C, respectively. All cultures were incubated in the presence of GA3 at a concentration of 1 mgl−1 supplemented in the culture medium or in the absence of GA3 in the culture medium and was repeated three times. The cultures were maintained at 25˚C under cool-white fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod.
2.3. Effect of the Developmental Stage of the Microspore on Male Gametophyte Formation
To examine the effect of the developmental stage of the microspore on male gametophyte formation, white immature and light-brown mature microspores were cultured in MS basal liquid medium and distilled water, respectively. The pH of the culture medium was adjusted to 5, 5.8, 6.4 and 7 with 1N NaOH. Also, the pH of the distilled water was 6.2. After four weeks of incubation, the frequency of male gametophyte formation was examined. After only two weeks, initial morphological changes of the microspores were observed under microscopy.
2.4. Effect of Light Intensity and Stress (Heat and Cold Shock) on Sporophyte Formation from Mixed Spore Culture
To examine the effect of light intensity from a mixed culture of microspores and megaspores on sporophyte formation, mixtures of microspores and megaspores were cultured in MS basal liquid medium and distilled water, respectively. The pH of the culture medium and sterile distilled water was adjusted to 7 with 1 N NaOH. Initial cultures consisted of approximately 100 megaspores. After four weeks of incubation, the number of megaspores developed into green sporophytes was counted.
We also examined the effect of plant growth regulators from a mixed culture of microspores and megaspores on sporophyte formation. Mixtures of microspores and megaspores were cultured in MS basal liquid medium supplemented with 0, 0.1, 0.3, 1, 3, 5, 10 mgl−1 of GA3. 2, 4-D and kinetin, respectively. Also, mixtures of microspores and megaspores were cultured on distilled water supplemented with 0, 0.1, 0.3, 1, 3, 5, 10 mgl−1 of GA3. 2, 4-D and kinetin, respectively. The pH of the culture medium and distilled water was adjusted to 7 with 1 N NaOH. After four weeks of incubation, the frequency of female gametophyte formation was examined. We also examined the effect of heat shock and cold shock on sporophyte formation from spore mixtures. Spore mixtures were pretreated for 72 hrs at 37˚C and 4˚C, respectively. After the cultures were subjected to either heat or cold shock, spore mixtures were cultured in MS basal liquid medium and distilled water. All cultures were done in triplicate. Unless otherwise noted, the cultures were maintained at 25˚C under cool-white fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod.
2.5. Acclimatization of Regenerated Sporophytes Spore Collection and Sterilization
After four weeks of incubating in the light (40 μmol·m−2·s−1 from cool-white fluorescent lamps with a 16-h photoperiod), green sporophytes were subjected to acclimation, transplanted to a test tube containing a vermiculite and sand mixture (1:1), and maintained in a growth chamber (25˚C day/22˚C night, 80 μmol·m−2·s−1 from cool-white fluorescent lamps with a 16-h photoperiod). After another two weeks of incubation in the light (80 mmol·m−2·s−1 from cool-white fluorescent lamps with a 16-h photoperiod), the sporophytes with roots and five leaves were transferred to potting soil.
3. Results and Discussion
3.1. Effect of the Developmental Stage of the Spore and Culture Conditions (pH, Temperature, and GA3) on Female Gametophyte Formation
Isoëtes is a representative heterosporous aquatic fern plant. Megasporoangium (Figure 1A) and microsporoangium (Figure 2A) produces numerous megaspores (approximately 100 to 200 spores per sporangium) and microspores by meiosis, respectively. In general, spore cultures of ferns require surface sterilization before germination procedures [17] although sterilization affected the viability of the spore to defend itself from fungal infections [13]. Sterilization can be done using trophosporophylla [18] and calcium hypochlorite [19]. To remove contaminants effectively, male and female sporangia were carefully detached from the leaf base and surface-sterilized directly. Afterwards, the sterilized spores were released into the culture medium. Freshly isolated mature megaspores showed typical spore size, structure, and surface ornamentation (i.e., cristate type) as seen in I. coreana [14]. The megaspores of I. coreana are trilete, 350 - 450 μm in diameter (Figure 1B). After 20 days of culturing in MS basal medium, the megaspores showed dramatic morphological changes during the culture. The spore wall was cracked by the expansion of the megaspores (about 2 times increase in diameter). Simultaneously, brown spots were observed on the surface of the megaspores. These structural changes were closely related to the initiation of female gametophyte formation (Figure 1C) [4,13]. Spore walls of immature megaspores Figure 1D) with white color also cracked by the expansion of the immature megaspores after three weeks of culturing (Figure 1E). These megaspores developed into female gametophytes with archegonia (Figure 1F).
The highest frequency of female gametophyte formation from immature megaspores was 46% when they were cultured in MS basal liquid medium adjusted to pH 7 in the presence of 1 mgl−1 GA3 (Table 1). However, female gametophyte formation from mature megaspores (12%) was about four times lower than those of imam-

Figure 1. Female gametophyte formation from megaspores of I. coreana. A: Megasporoangium; B: Mature megaspores with light brown color; C: Formation of female gametophytes from mature megaspores with archegonia; D: Immature megaspores with white color; E: After three weeks of culturing, spore walls were cracked by expansion of the megaspore; F: Formation of female gametophytes from immature megaspores with archegonia. Scale bars represents 2 mm (A), 300 um (B-D), and 500 um (E-F).

Table 1. Effect of the developmental stage of the megaspores, temperature and pH of culture medium on female gametophyte formation from megaspores of I. coreana. Mean ± Standard deviation.
ture megaspores. These results indicated that the developmental stage of the megaspores was critical for in vitro female gametophyte formation of I. coreana. In this study, we could not analyze the biochemical composition of the megaspores which differ depending on their maturity. However, we did observed the change in color of the megaspores from white to brown as they matured (Figure 1C). Therefore, we inferred that mature megaspores accumulated spore germination inhibitors for dormancy whereas immature megaspores did not. Elucidating the presence of germination inhibitors in megaspores would be necessary and would be an interesting study into the life cycle of I. coreana.
Culture temperature was also critical for in vitro female gametophyte formation of I. coreana (Table 1). The frequency of female gametophyte formation from immature megaspores increased when they were cultured at 25˚C. However, the frequency of female gametophyte formation decreased when the culture temperature was lowered (18˚C). These results showed that the culture temperature of megaspores was significant for in vitro female gametophyte formation of I. coreana (Table 1). We also investigated the effect of the pH of the culture medium. In an acidic condition (pH 5), the frequency of female gametophyte formation was less than 10%. However, the frequency of female gametophyte formation gradually increased when the acidity decreased to neutral conditions (5.8 to 7). GA3 is a well-known seed germination stimulator. However, the presence of GA3 in culture medium did not show a significant increase in the formation of the female gametophyte of I. coreana. In preliminary studies, we tried to induce callus formation from various tissues of I. coreana. However, PGRs did not show a promotive role for callus and female gametophyte formation (data not shown). These results implied that the sensitivity of Isoëtes to PGRs was different from those of higher plants and terrestrial ferns. Interestingly, we also found that female gametophyte formation of I. coreana was very successful in distilled water compared to MS basal medium except for cultivation at 25˚C for immature megaspores (Table 1). Taking into consideration the natural habitat of Isoëtes is water, it is no surprise that water has the similar potential for female gametophyte formation. The female gametophytes obtained from MS basal medium and distilled water did not show further differentiation. After prolonged incubation, female gametophytes deteriorated and turned a dark brown color and died (data not shown).
The changes of temperature and acidity are important environmental cues for the determination of various
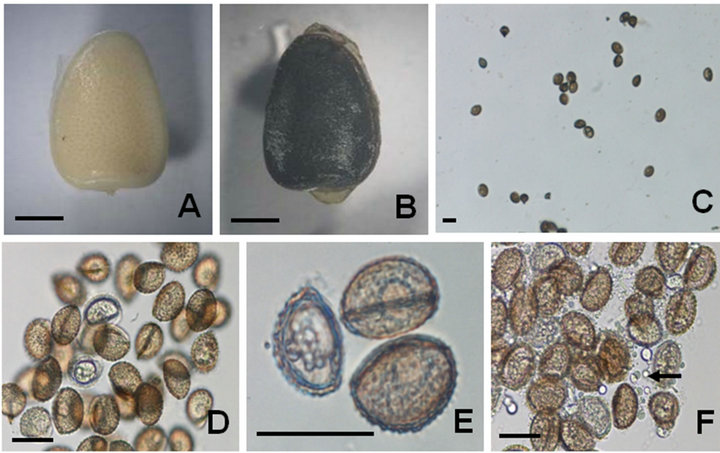
Figure 2. Male gametophyte formation from microspores of I. coreana. A: Immature microsporoangium with white color; B: Mature microsporoangium with dark brown color; C: Microspores released from microsporoangium; D: Enlarged view of microspores after three weeks of culturing. E: Formation of male gametophytes from mature microspores with morphological changes; F: Release of granules (inferring sperms) from male gametophytes after three weeks of culturing. Scale bars represent 2 mm (A-B), and 30 um (C-F).
physiological processes in plants. Until now, the exact mechanism of sporophyte formation of Isoëtes in nature was a mystery. In this study, we found that the degree of maturity of the megaspores, temperature and pH of culture medium had a significant role in the promotion of female gametophyte formation. Considering these results, we suggest that the warm season after cold hardening would be suitable for gametophyte formation from megaspores of I. coreana in natural habitats.
3.2. Male Gametophyte Formation from Microspore Culture
Freshly isolated microspores of I. coreana showed similar spore size, structure and surface ornamentation (i.e., echinate type) to the original description of the species [14]. The microspores of I. coreana are monolete, rounded, 30 - 36 μm long and less than 23 μm wide. Immature (Figure 2A) and mature microsporoangium (Figure 2B) produce numerous microspores by meiosis (Figure 2C). After 20 days of culturing in the MS basal medium, microscopic examination showed significant morphological changes (Figure 2D). These structural changes were closely related to the formation of the male gametophyte (Figure 2E). After further incubation, a number of small vesicles (inferring sperms) were released into the culture medium (Figure 2F) [13]. We also examined the effects of pH, temperature, maturity and PGRs on male gametophyte formation. However, we could not find out the promotive effects between several treatments (data not shown). What is seen here is similar to the results obtained from Isoëtes species of North America [13]. The size of the microspores of I. coreana was smaller than those of Isoëtes species (Lycophyta) from the Southern area of South America. Macluf et al. [20] reported that the size of the microspores was positively correlated with increasing ploidy level, and the larger microspores were associated with terrestrial habitats. The natural habitats of I. coreana are aquatic ponds. The formation of the sporophyte of Isoëtes can take place through sexual reproduction. Therefore, the presence of water is necessary for the fusion of the male and female gametes.
3.3. Effect of Light Intensity, Heat and Cold Shock on Sporophyte Formation from Mixed Spore Cultures
High frequency sporophyte regeneration system via a mixed culture of microspores and megaspores of I. coreana was successfully established (Figure 3). After initiation of static incubation, numerous microspores adhered to megaspores (Figure 3A). And then, megaspores were totally enclosed by microspore aggregates after one week of culturing (Figure 3B). After two weeks of cul-
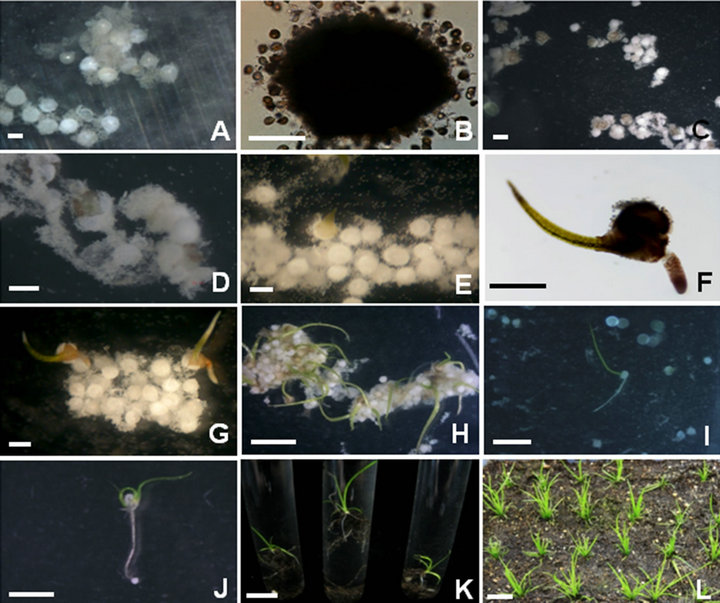
Figure 3. Sporophyte formations from mixed spore cultures of I. coreana. A: Initial adhesion between numerous microspores and megaspores; B: Enlarged view of adhesion between microspores and megaspores; C: Initial development of female gametophytes after two weeks of culturing; D: Enlarged view of initial female gametophytes formation; E: Elongation of the first leaf from young sporophytes; F: Enlarged view of young sporophytes with rooting; G: Multiple young sporophytes formation from mixed spore culture; H: Elongation of the first leaf from green young sporophytes; I: Elongation of roots from young sporophytes; J: Emergence of the second leaf from young sporophytes; K: Acclimatization of regenerated sporophytes in test tubes; L: Soil transfer of sporophytes after acclimatization. Scale bars represents 100 um (B), 300 um (A, C-G), 1 cm (H-K) and 5 cm (L).
turing, the spore walls cracked and lightbrown female gametophytes emerged from the megaspores (Figures 3C and D). These small light-green female gametophytes developed into young green sporophytes (Figure 3E). After another week of incubation, young sporophytes with rooting were formed without any treatments (Figures 3F and G). The exact developmental mechanism of sporophyte formation from Isoëtes species has not been fully elucidated yet [8]. The formation of the sporophyte of Isoëtes can take place through sexual reproduction. Therefore, the fusion of the eggs and sperms is necessary for sporophyte formation. In this study, we could not examine the exact route of spore fusion. However, we observed that female gamete formation was possible from the mixed culture of male and female spores (Figure 3) or megaspores only cultures (Figure 2). We also found that the female gametes derived from mixed spore cultures were able to develop into green young sporophytes whereas the female gametes derived from megaspore only cultures were not able to do at all. These results clearly showed that mix of male and female spores was critical for sporophyte development of Isoëtes. Considering these results, we inferred that there were any biochemical processes between sperm and egg triggering the development of sporophyte during mixed spore cultures. Further studies are needed for the elucidation of initial developmental processes of sporophyte formation.
The highest frequency (12.3%) of green sporophyte formation from mixed spore cultures occurred when the cultures were maintained at 25˚C under cool-white fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod (Figure 4). When the light intensity decreased (10 μmol·m−2·s−1) with the other culture conditions remaining the same, the frequency of sporophyte formation was slightly decreased (11%). Even though the light intensity did not significantly affect the frequency of sporophyte formation, however further development of young sporophyte was retarded in the low light treatments. Furthermore, we could not observe the sporophyte formation in the absence of light. In addition female gametophytes formed in the dark incubation were able to develop normal green sporophytes when they were transferred to a light condition. These results indicated that the process of sporophyte formation of I. coreana required light. And suitable light intensity is also necessary for further development and differentiation of sporophyte.
We also examined the effect of temperature on sporophyte formation (Figure 4). The frequency of sporophyte formation was 9.5% when cultured at 18˚C under coolwhite fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod. Although the effect of low temperature was not significant when compared to a megaspore only culture (Table 1), we found that the frequency of sporophyte formation was decreased at a low temperature. In severe heat shock (37˚C for 72 hrs) and cold shock (4˚C for 72 hrs) pretreatment, the frequency of sporophyte formation sharply decreased to 0.9% and 1.6%, respectively. In addition the culture periods required for emergence of sporophytes from a mixed spore culture was prolonged approximately one week when the culture temperature was lowered to 18˚C. This result indicated
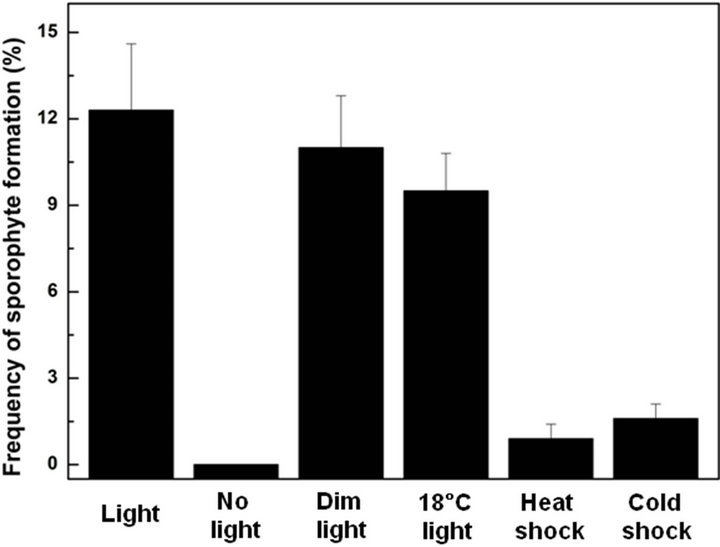
Figure 4. Effects of light intensity, temperature and pretreatments on sporophyte formation from mixed spore cultures of I. coreana.
that culture temperature was also critical for sporophyte formation. This result was in good agreement with those obtained from temperature requirement for germination of Isoëtes species from north america [13]. Considering these results, we suggest that low temperature has an inhibitory role in gametophyte formation as well as sporophyte formation. Furthermore, the cold treatment serves to postpone germination until a time when the temperature rises to a sufficient level where germination is not inhibited.
3.4. Acclimatization of Regenerated Sporophytes
After elongation of the first leaf and root, the second leaf was developed from the corm lobes (Figures 3H-J). After four weeks of culturing, two to three leaves were developed from the sporophytes. Preliminarily, we examined what would be a suitable stage for regenerated sporophytes to be transferred to the soil. Sporophytes with less than three leaves could not survive when transferred to vermiculite and sand mixture (data not shown). Sporophytes with more than three leaves were transferred to test tubes containing a vermiculite and sand mixture (1:1). And then sterile distilled water was added into the tube (Figure 3K). The fifth leaf appeared after another two weeks of incubation at 25˚C under cool-white fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod. More than 95% of the regenerated sporophytes from spore culture were successfully acclimatized. These sporophytes were transferred again to an ornamental soil mixture without water filling and grown to normal sporophytes (Figure 3L). After another 5 months of culturing at 25˚C under cool-white fluorescent light (40 μmol·m−2·s−1) with a 16 h photoperiod, sporophytes were grown to mature sporophytes with microsporangia and megasporangia.
In this study, we established a high frequency sporophyte regeneration system with an incubation time of two months via a mixed culture of microspores and megaspores of I. coreana. To our knowledge, this is the first successful report of in vitro proliferation of sporophyte of I. coreana. The sporophyte regeneration system established in this study could be directly applied for the restoration of the endangered species, I. coreana and used for the mass proliferation of this plant in vitro. Also, regenerated sporophytes from mixed spore cultures could be used for studying the genetic relationship. Furthermore, an in vitro culture system of I. coreana could be applied for an alternative conservation system of germplasm.
4. Acknowledgements
This work was supported by a grant from KRIBB Research Initiative Program, and grant from the Ministry of Environment, as the Eco-Technopia 21 project (KIEST 052-071-049).
REFERENCES
- W. C. Taylor and R. J. Hickey, “Habitat, Evolution and Speciation in Isoetes,” Annals of the Missouri Botanical Garden, Vol. 79, No. 3, 1992, pp. 613-622. doi:10.2307/2399755
- W. C. Taylor, N. T. Luebke, D. M. Britton, R. J. Hickey and D. F. Brunton, “Isoetaceae,” FNA Editorial Committee, Flora of North America, Oxford University Press, New York, 1993, pp. 64-75.
- J. E. Keeley, “CAM Photosynthesis in Submerged Aquatic Plants,” Botanical Review, Vol. 64, No. 2, 1998, pp. 121- 175. doi:10.1007/BF02856581
- N. S. Parihar, “The Biology and Morphology of Pteridophytes,” Indian Universities Press, Allahabad, 1996.
- K. R. Stern, “Introductory Plant Biology”, 3rd Edition, W. C. Brown, Publishers, Dubuque, 1985, p. 515.
- COSEWIC, “COSEWIC Assessment and Status Report on the Prototype Quillwort Isoetes prototypus in Canada,” Committee on the Status of Endangered Wildlife in Canada, Ottawa, 2005.
- C. A. Caplen and C. R. Werth, “Isozymes of the Isoëtes riparia Complex, I. Genetic Variation and Relatedness of Diploid Species,” Systematic Botany, Vol. 25, No. 2, 2000, pp. 235-259. doi:10.2307/2666641
- S. J. Sam, “A Germination Method for Isoëtes,” American Fern Journal, Vol. 72, No. 2, 1982, p. 61. doi:10.2307/1547062
- W. C. Taylor and N. T. Luebke, “Germinating Spores and Growing Sporelings of Aquatic Isoetes,” American Fern Journal, Vol. 76, No. 1, 1986, pp. 21-24. doi:10.2307/1547396
- H. Fernandez and M. A. Revilla, “In Vitro Culture of Ornamental Ferns,” Plant Cell, Tissue and Organ Culture, Vol. 73, No. 1, 2003, pp. 1-13. doi:10.1023/A:1022650701341
- D. Cachita-Cosma, “Metode in Vitro la Plantele de Cultura,” Edit. Ceres, Bucuresti, Agricultural Sciences, University of Florida, 1987. http://edis.ifas.ufl.edu
- B. M. Boom, “Intersectional Hybrids in Isoëtes,” American Fern Journal, Vol. 70, No. 1, 1980, pp. 1-4. doi:10.2307/1546200
- L. S. Koot and D. M. Britton, “A Comparative Study of Spore Germination of Some Isoëtes Species of Northeastern North America,” Canadian Journal of Botany, Vol. 60, No. 9, 1982, pp. 1679-1687. doi:10.1139/b82-218
- Y. H. Chung and H.-K. Choi, “ Isoëtes coreana, a New Species from Korea,” Korean Journal of Plant Taxonomy, Vol. 16, 1986, pp. 1-12 (in Korean, with English Abstract).
- C. Kim, H. R. Na and H.-K. Choi, “Genetic Diversity and Population Structure of Endangered Isoetes coreana in South Korea Based on RAPD Analysis,” Aquatic Botany, Vol. 89, No. 1, 2008, pp. 43-49. doi:10.1016/j.aquabot.2008.02.004
- T. Murashige and F. Skoog, “A Revised Medium for Rapid Growth and Bioassays with Tobacco Tissue Culture,” Plant Physiology, Vol. 15, No. 3, 1962, pp. 473- 497. doi:10.1111/j.1399-3054.1962.tb08052.x
- K. P. Martin, S. Sini, C.-L. Zhang, A. Slater and P. V. Madhusoodanan, “Efficient Induction of Apospory and Apogamy in Vitro in Silver Fern (Pityrogramma calomelanos L.),” Plant Cell Reports, Vol. 25, No. 12, 2006, pp.1300-1307. doi:10.1007/s00299-006-0215-5
- S Hedge, “Propagation of Some Ornamental Plants Using Biotechnology,” Dissertation, Mangalore University, Mangalore, 1998.
- E. A. Simabukuro, A. F. Dyer and G. M. Felippe, “The Effect of Sterilization and Storage Conditions on the Viability of Spores of Cyathea delgadii,” American Fern Journal, Vol. 88, No 2, 1998, pp. 124-132. doi:10.2307/1547226
- C. C. Macluf, M. A. Morbelli and G. E. Giudice, “Microspore Morphology of Isoetes Species (Lycophyta) from Southern South America,” Botanical Review, Vol. 72, No. 2, 2006, pp. 121-134. doi:10.1663/0006-8101(2006)72[121:MMOISL]2.0.CO;2
NOTES
*Corresponding authors.