Open Journal of Blood Diseases
Vol.3 No.4(2013), Article ID:40772,14 pages DOI:10.4236/ojbd.2013.34023
AKT3 rSNPs, Transcriptional Factor Binding Sites and Human Disease
![]()
Department of Pediatrics, University of Washington, Seattle, USA.
Email: nburoker@u.washington.edu
Copyright © 2013 Norman E. Buroker. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. In accordance of the Creative Commons Attribution License all Copyrights © 2013 are reserved for Hans and the owner of the intellectual property Norman E. Buroker. All Copyright © 2013 are guarded by law and by Hans as a guardian.
Received August 28th, 2013; revised September 28th, 2013; accepted October 5th, 2013
Keywords: AKT3; rSNPs; TFBS; Human Disease
ABSTRACT
Seven rSNPs (rs10157763, rs10927067, rs12031994, rs2125230, rs2345994, rs4132509 and rs4590646) in intron one of thev-akt murine thymoma viral oncogene homolog 3 (AKT3) gene have been significantly associated with either Chronic Mountain Sickness, Renal Cell Carcinoma risk or Aggressive Prostate Cancer. These rSNP alleles alter the DNA landscape for potential transcriptional factors (TFs) to attach, resulting in changes in transcriptional factor binding sites (TFBS). The alleles of each rSNP were found to produce unique TFBS resulting in potential changes in TF AKT3 regulation. These regulatory changes are discussed with respect to the three diseases.
1. Introduction
The phosphatidylinositol 3-kinase (PI3K)/AKT pathway plays a key role in numerous cellular functions including proliferation, adhesion, migration, invasion, metabolism and survival [1] as well as angiogenesis. The PI3K/AKT pathway modulates the expression of angiogenic factors such as nitric oxide and angiopoietins [2]. The v-akt murine thymoma viral oncogene homolog 3 (AKT3) is one of three isoforms of the AKTs which are major downstream targets of growth factor receptor tyrosine kinases that signal through PI3K [3]. The AKT3 single nucleotide polymorphism (SNP, rs4590656) from intron one was recently found to be significantly associated with three physiological parameters (hemoglobin, hematocrit and red blood cell count) in chronic mountain sickness patients indicating the strong association of this gene with angiogenesis [4]. Other intron one AKT3 SNPS (rs10157763, rs10927067 and rs2125230) have been significantly associated with aggressive prostate cancer (PCA) [5] in addition to the SNPs (rs4132509, rs- 12031994, rs2345994) which have been recently found to be significantly associated with renal cell carcinoma risk (RCC) [6]. This suggests that intron one maybe a regulatory region for the AKT3 gene. To follow up on this possibility these seven SNPs were examined for associations to potential transcription factor binding sites (TFBS).
Single nucleotide changes that affect gene expression by impacting gene regulatory sequences such as promoters, enhances, and silencers are known as regulatory SNPs (rSNPs) [7-10]. A rSNPs within a transcriptional factor binding site (TFBS) can change a transcriptional factor’s (TF) ability to bind its TFBS [11-14] in which case the TF would be unable to effectively regulate its target gene [15-19]. This concept is examined for the above AKT3 rSNPs of intron one and their allelic association with TFBS. In this report, the rSNP associations with changes in potential TFBS and their possible relationship to the reported diseases or conditions have been examined.
2. Materials and Methods
2.1. Identifying TFBS
The JASPAR CORE database [20,21] and ConSite [22] were used to identify the TFBS in this study. JASPAR is a collection of transcription factor DNA-binding preferences used for scanning genomic sequences where ConSite is a web-based tool for finding cis-regulatory elements in genomic sequences. The TFBS and rSNP location within the binding sites have previously been discussed [4]. The Vector NTI Advance 11 computer program (Invitrogen, Life Technologies) was used to locate the TFBS in the AKT3 gene (NCBI Ref Seq NM_005465) from 11 kbp upstream of the transcriptional start site to 400 bp past the 3’UTR which represents a total of 35.5 Kbp.
3. Results
3.1. AKT3 rSNPs and TFBS
The AKT3 gene transcribes a serine/threonine kinase that plays a key role in regulating cell survival, insulin signaling, angiogenesis and tumor formation. The AKT3 SNP (rs4590656) has been found to be significantly associated with Hb and Hct in Tibetan Chinese with CMS. The AKT3-C allele creates two unique TFBS for the ARNT: AHR and HIF1α: ARNT TFs which are involved xenobiotic metabolism and cellular and systemic responses to hypoxia, respectively. The AKT3-T allele creates other unique TFBS for the HNF4α and PAX2 TFs which are involved in the expression of several hepatic genes and the conserved DNA-binding paired box, respectively which have previously been reported. In addition, the AKT3-T allele also creates two unique TFBS for the GFI TF which functions as a repressor and plays a role in hematopoiesis and oncogenesis and the SPIB TF which is a transcriptional activator that acts as a lymphoid-specific enhancer (Table 1 and Supplement).
The AKT3 SNPs (rs4132509, rs12031994 and rs2345994) are significantly associated with the risk of RCC [6]. The rs4132509 AKT3-C allele creates one unique TFBS for the NFE2L1:MAFG TF, where NFE2L1 coordinates the up-regulation of cytoprotective genes via the antioxidant response element while MAFG is involved in cell differentiation of erythrocytes. The minor AKT3-A allele does not create any uniquely different TFBS (Table 1 and Supplement). The rs12031994 AKT3- G allele creates seven unique TFBS for the FOXD1, FOXF2, FOXO3, NFYA, NKX2-5, SOX5 and SOX9 TFs those functions are described in the supplement. One TF standout is that FOXD1 may play a role in tumor formation which should have an effect on RCC. The minor AKT3-A allele creates nine unique TFBS for the EN1, GATA1, 2 & 3, HLTF, PDX1, PRRX2, TAL1: GALA1 and ZNF354C TFs those functions are also described in the supplement. Of these the TAL1: GALA1 TF is of interest since it is involved with the genesis of hemopoietic malignancies and may play a role in hemopoietic differentiation and a positive regulator of erythroid differentiation. The rs2345994 AKT3-G allele creates two unique TFBS for the SRY and SOX10 TFs which are part of a family of TFs involved in the regulation of embryonic development and in the determination of the cell fate (Supplement). The minor AKT3-A allele creates four unique TFBS for the ARID3A, FOXL1, MEF2A and NKX2-5 TFs of which the MEF2A TF is probably the most interesting since it activates many muscle-specific, growth factor-induced and stress-induced genes (Table 1 & Supplement).
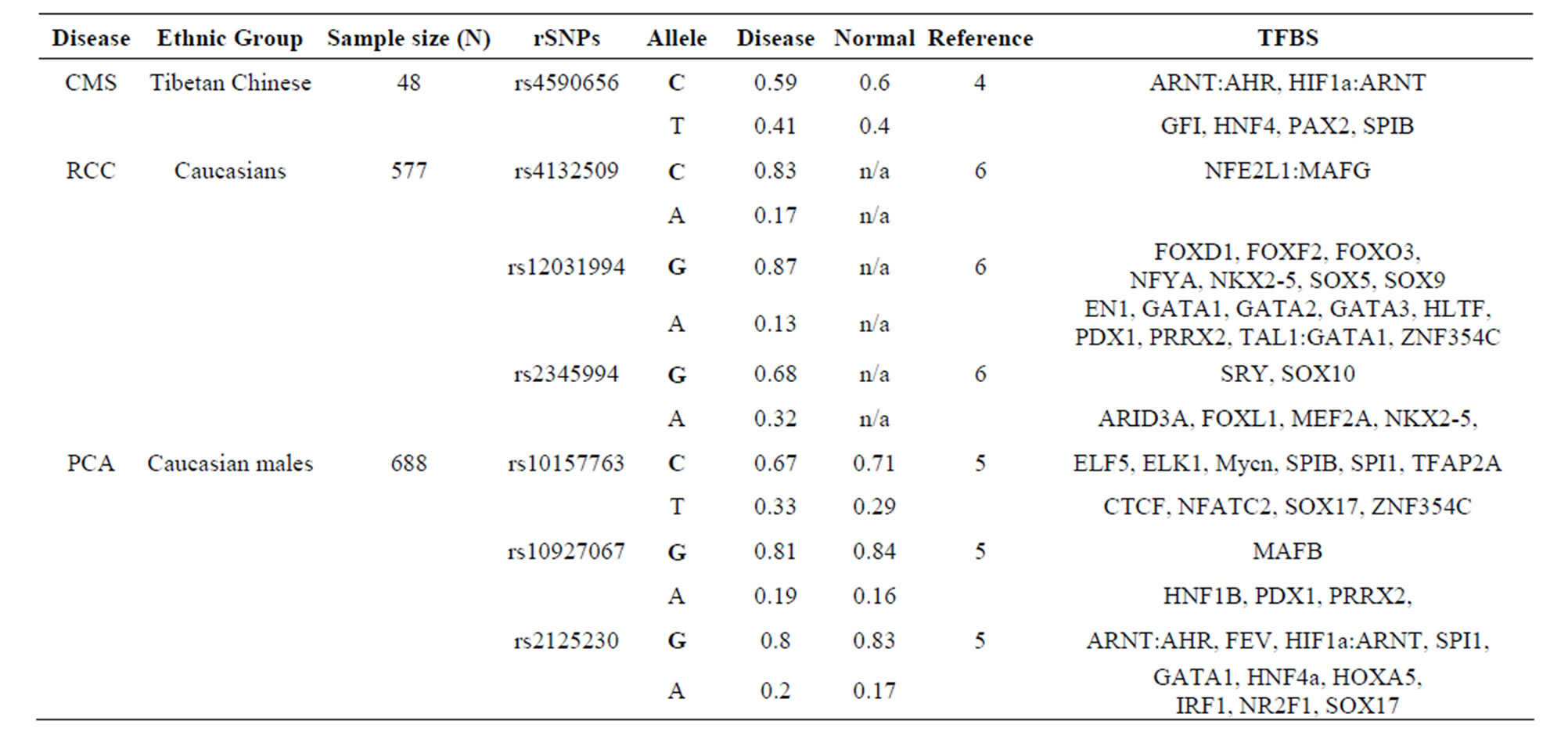
Table 1. Human diseases and AKT3 rSNPs found to be significantly associated in the referenced study. Common allele in bold. rSNPs alleles alter the transcriptional factor binding sites (TFBS) in intron one of the gene. The rSNP alleles are found only in these TFBS. CMS = Chronic Mountain Sickness; RCC = Renal Cell Carcinoma risk; PCA = Aggressive Prostate Cancer.
The AKT3 SNPs (rs10157763, rs10927067 and rs- 2125230) are significantly associated with PCA [5]. The rs10157763 AKT3-C allele creates six unique TFBS for the ELF5, ELK1, Mycn, SPIB, SPI1 and TFAP2A TFs whose function are described in the Supplement. Of these TFs the Mycn is probably the most interesting since it is a member of the MYC family and is associated with a variety of tumors. The minor AKT3-T allele creates four unique TFBS for the CTCF, NFATC2, SOX17 and ZNF354C TFs which are part of the transcription machinery for gene regulation (Table 1 and Supplement). The rs10927067 AKT3-G allele creates one unique TFBS for the MAFB TF that is important in the regulation of lineage-specific hematopoiesis. The minor AKT3-A allele creates three unique TFBS for the HNF1B, PDX1 and PRRX2 TFs of which the HNF1B TF is of interest because it is a member of the homeodomain-containing superfamily of transcription factors and regulates development of the embryonic pancreas (Table 1 and Supplement). The rs2125230 AKT3-G allele creates four unique TFBS for the ARNT: AHR, FEV, HIF1α: ARNT and SPI1 TFs. The ARNT: AHR and HIF1α: ARNT TFs are discussed above while the FEV and SPI1 TFs are involved as a transcriptional regulator and involved in myeloid and B-lymphoid cell development, respectively (Table 1 and Supplement). The minor AKT3-A allele creates six unique TFBS for the GATA1, IRF1, HNF4a, NR2F1, SOX17 and HOXA5 TFs where the HOXA5 TF is of interest since it upregulates the tumor suppressor p53 and has an important role in tumorigenesis.
4. Discussion
Genome-wide association studies (GWAS) over the last decade have identified nearly 6500 disease or trait-predisposing SNPs where only 7% of these are located in protein-coding regions of the genome [23,24] and the remaining 93% are located within non-coding areas [25,26] such as regulatory or intergenic regions. SNPs which occur in the putative regulatory region of a gene where a single base change in the DNA sequence of a potential TFBS may affect the process of gene expression are drawing more attention [7,9,27]. A SNP in a TFBS can have multiple consequences. Often the SNP does not change the TFBS interaction nor does it alter gene expression since a transcriptional factor (TF) will usually recognize a number of different binding sites in the gene. In some cases the SNP may increase or decrease the TF binding which results in allele-specific gene expression. In rare cases, a SNP may eliminate the natural binding site or generate a new binding site. In which cases the gene is no longer regulated by the original TF. Therefore, functional rSNPs in TFBS may result in differences in gene expression, phenotypes and susceptibility to environmental exposure [27]. Examples of rSNPs associated with disease susceptibility are numerous and several reviews have been published [27-30].
The AKT3 SNP (rs4590656) in the potential TFBS and the corresponding nucleotide occurrence (%) in the BS (Supplement) have previously been discussed [4] as well as the number of other similar TFBS found in the gene. The rs4132509 AKT3-C allele located in the NFE2L1: MAFG TFBS has a 76% occurrence in humans; however, this BS occurs 140 other times in the gene (Supplement) and therefore the SNP would probably not have any impact on the regulation of the gene. The rs4132509 AKT3- A allele does not have any unique TFBS and therefore should not have any impact on gene regulation. The rs12031994 AKT3-G allele occurs in seven unique TFBS (Table 1), but perhaps the most interesting two are FOXF2 and NFYA which each occur only once in the gene. The rs12031994 AKT3-G allele [G (−strand) or C (+strand)] is a highly conserved nucleotide (97% and 93%) in these respective TFBS. The rs12031994 AKT3- A allele occurs in nine unique TFBS (Table 1), but perhaps the most interesting two are GATA1 and TAL1: GATA1 which again each only occurs once in the gene. The rs12031994 AKT3-A allele [A (−strand) or T (+strand)] is a highly conserved nucleotide (96% and 98%) in these respective TFBS. This SNP might be expected to have an influence on AKT3 regulation and could impact human disease. The rs2345994 AKT3-G allele occurs in two unique TFBS (SRY and SOX10) which both have multiple copies in the gene (Table 1). The rs2345994 AKT3-A allele occurs in four unique TFBS (Table 1), but perhaps the most interesting one is the MEF2A TFBS since it occurs only three times in the gene and the A nucleotide has a 98% occurrence (Supplement). The rs10157763 AKT3-C allele occurs in six unique TFBS (Table 1), but perhaps the most interesting TFBS is TFAP2A since it occurs four times in the gene and the allele is 100% conserved (Supplement). The rs10157763 AKT3-T allele occurs in four unique TFBS (Table 1) but perhaps the most important TFBS is the CTCF TF which occurs only once in the gene and is a transcriptional regulator protein with 11 highly conserved zinc finger domains. The rs10927067 AKT3-G allele occurs only in one unique TFBS which is MAFB. The MAFB TFBS occurs only two times in the gene and the G nucleotide has an 80% occurrence in the sequence for humans. The rs10927067 AKT3-A allele occurs in three unique TFBS, but perhaps only the HNF1B TFBS would be of interest since it occurs once in the gene and the A nucleotide has an 80% occurrence in humans. The rs2125230 AKT3-G allele occurs in four unique TFBS, but only the FEV and HIF1a:ARNT TFBS would be of interest since they both occur three times in the gene and G nucleotide occurrence is 100% and 99%, respectively. The rs2125230 AKT3-A allele has six unique TFBS, but perhaps the IRF1 and NR2F1 would be of interest since they both occur only once in the gene and A nucleotide occurrence is 95% and 92%, respectively.
Kidney cancer accounted for an estimated 4% of new cancer cases in the United States in 2012 [31] and approximately 85% of the kidney cancers are renal cell carcinomas (RCCs) [32]. There has been compelling evidence for genetic susceptibility to RCC [33-35]. Recently, a RCC study involving SNP polymorphisms in the mTOR pathway which includes the AKT3 gene found that obesity, weight gain, physical activity and genetic variants may influence susceptibility to the disease [6]. After analyzing three AKT3 SNPs which were found to be statistically significant in the RCC study, it was determined that these SNPs do alter the potential TFBS for TFs regulating the gene (Table 1). Perhaps the most interesting TFBS change occurs with the SNP rs12031994; as an example, TAL1: GATA1 occurs only with the minor AKT3-A allele and is implicated in the genesis of hemopoietic malignancies.
Prostate cancer (PCA) is the second leading cause of cancer-related deaths among men in the United States [36] and the American Cancer Society estimates that 26% - 29% of all new cancer cases and cancer-related deaths are attributed to PCA cancer [36]. PCA risk factors include older age, ethnic race and family history while other potential contributors include lifestyle, genetic factors and imbalances in biological pathways [5]. GWAS has resulted in the detection of numerous PCA susceptibility loci [37] from which three AKT3 SNPs that have been previously associated with PCA [5] have been found to be also associated with changes in TFBS within intron one. Perhaps the most interesting TFBS change occurs with the rs10157763 SNP whose minor allele AKT3-T creates the chromatin insulator-binding factor (CTCF) TFBS that is found only once in the gene (Table 1 and Supplement). CTCF is a highly conserved zinc finger TF implicated in diverse genomic regulatory functions, including transcriptional activation/repression, insulation, imprinting [38] which restricts enhancers from activating AKT3 and thereby restraining angiogenesis [39]. The CTCF protein has been shown to be involved with chromatin loop formation and co-associates with the zinc finger domains [38] such as the Zfx TFBS. Also worth mentioning is the change in the interferon (IRF1) TFBS which occurs with the SNP rs2125230 minor allele AKT3-A allele. Although other IRF1 TFBS are found in the gene this BS has a 95% occurrence of the A nucleotide which is regulated by the SNP (Supplement). Changes in the IRS1 TFBS for the gene are important since interferon is involved in fighting tumors.
Human diseases or conditions that have been significantly associated with rSNPs of the AKT3 gene are shown in the Table 1 along with rSNP allele-specific TFBS. What a change in the rSNP alleles can do, is to alter the DNA landscape around the SNP for potential TFs to attach and regulate a gene. This change in the regulatory landscape can alter gene regulation which in turn can result in human disease, a change in condition or illness. In this report, examples have been described to illustrate that a change in rSNP alleles can provide different TFBS which in turn are also significantly associated with human disease.
REFERENCES
- A. G. Bader, S. Kang, L. Zhao and P. K. Vogt, “Oncogenic PI3K Deregulates Transcription and Translation,” Nature Reviews. Cancer, Vol. 5, No. 12, 2005, pp. 921-929.
- J. Karar and A. Maity, “PI3K/AKT/mTOR Pathway in Angiogenesis,” Frontiers in Molecular Neuroscience, Vol. 4, 2011, p. 1. http://dx.doi.org/10.3389/fnmol.2011.00051
- J. R. Testa and A. Bellacosa, “AKT Plays a Central Role in Tumorigenesis,” Proceedings of the National Academy Sciences of the United States of America, Vol. 98, No. 20, 2001, pp. 10983-10985. http://dx.doi.org/10.1073/pnas.211430998
- N. E. Buroker, X. H. Ning, Z. N. Zhou, K. Li, W.J. Cen, X. F. Wu, W. Z. Zhu, C. R. Scott and S. H. Chen, “AKT3, ANGPTL4, eNOS3, and VEGFA Associations with high Altitude Sickness in Han and Tibetan Chinese at the Qinghai-Tibetan Plateau,” International Journal of Hematology, Vol. 96, No. 2, 2012, pp. 200-213. http://dx.doi.org/10.1007/s12185-012-1117-7
- N. A. Lavender, E. N. Rogers, S. Yeyeodu, J. Rudd, T. Hu, J. Zhang, G. N. Brock, K. S. Kimbro, J. H. Moore, D. W. Hein and L. C. Kidd, “Interaction among ApoptosisAssociated Sequence Variants and Joint Effects on Aggressive Prostate Cancer,” BMC Medical Genomics, Vol. 5, No. 11, 2012 , p. 1-15. http://dx.doi.org/10.1186/1755-8794-5-11
- X. Shu, J. Lin, C. G. Wood, N. M. Tannir and X. Wu, “Energy Balance, Polymorphisms in the mTOR Pathway, and Renal Cell Carcinoma Risk,” Journal of the National Cancer Institute, Vol. 105, No. 6, 2013, pp. 424-432. http://dx.doi.org/10.1093/jnci/djt005
- J. C. Knight, “Functional Implications of Genetic Variation in Non-Coding DNA for Disease Susceptibility and Gene Regulation,” Clinical Science, Vol. 104, No. 5, 2003, pp. 493-501. http://dx.doi.org/10.1042/CS20020304
- J. C. Knight, “Regulatory Polymorphisms Underlying Complex Disease Traits,” Journal of Molecular Medicine, Vol. 83, No. 2, 2005, pp. 97-109. http://dx.doi.org/10.1007/s00109-004-0603-7
- X. Wang, D. J. Tomso, X. Liu and D. A. Bell, “Single Nucleotide Polymorphism in Transcriptional Regulatory Regions and Expression of Environmentally Responsive Genes,” Toxicology and Applied Pharmacology, Vol. 207, No. 2, 2005, pp. 84-90. http://dx.doi.org/10.1016/j.taap.2004.09.024
- X. Wang, D. J. Tomso, B. N. Chorley, H. Y. Cho, V. G. Cheung, S. R. Kleeberger and D. A. Bell, “Identification of Polymorphic Antioxidant Response Elements in the Human Genome,” Human Molecular Genetics, Vol. 16, No. 10, 2007, pp. 1188-1200. http://dx.doi.org/10.1093/hmg/ddm066
- F. Claessens, G. Verrijdt, E. Schoenmakers, A. Haelens, B. Peeters, G. Verhoeven and W. Rombauts, “Selective DNA Binding by the Androgen Receptor as a Mechanism for Hormone-Specific Gene Regulation,” The Journal of Steroid Biochemistry and Molecular Biology, Vol. 76, No. 1-5, 2001, pp. 23-30. http://dx.doi.org/10.1016/S0960-0760(00)00154-0
- M. H. Hsu, U. Savas, K. J. Griffin and E. F. Johnson, “Regulation of Human Cytochrome P450 4F2 Expression by Sterol Regulatory Element-Binding Protein and Lovastatin,” Journal of Biological Chemistry, Vol. 282, No. 8, 2007, pp. 5225-5236. http://dx.doi.org/10.1074/jbc.M608176200
- H. Takai, S. Araki, M. Mezawa, D. S. Kim, X. Li, L. Yang, Z. Li, Z. Wang, Y. Nakayama and Y. Ogata, “AP1 Binding Site Is Another Target of FGF2 Regulation of Bone Sialoprotein Gene Transcription,” Gene, Vol. 410, No. 1, 2008, pp. 97-104. http://dx.doi.org/10.1016/j.gene.2007.11.017
- N. E. Buroker, J. Y. Huang, J. Barboza, D. R. Ledee, R. J. Eastman Jr., H. Reinecke, X. H. Ning, J. A. Bassuk and M. A. Portman, “The Adaptor-Related Protein Complex 2, Alpha 2 Subunit (AP2α2) Gene Is a Peroxisome Proliferator-Activated Receptor Cardiac Target Gene,” The Protein Journal, Vol. 31, No. 1, 2012, pp. 75-83. http://dx.doi.org/10.1007/s10930-011-9379-0
- C. N. Huang, S. P. Huang, J. B. Pao, T. C. Hour, T. Y. Chang, Y. H. Lan, T. L. Lu, H. Z. Lee, S. H. Juang, P. P. Wu, C. Y. Huang, C. J. Hsieh and B. Y. Bao, “Genetic Polymorphisms in Oestrogen Receptor-Binding Sites Affect Clinical Outcomes in Patients with Prostate Cancer Receiving Androgen-Deprivation Therapy,” Journal of Internal Medicine, Vol. 271, No. 5, 2012, pp. 499-509. http://dx.doi.org/10.1111/j.1365-2796.2011.02449.x
- C. N. Huang, S. P. Huang, J. B. Pao, T. Y. Chang, Y. H. Lan, T. L. Lu, H. Z. Lee, S. H. Juang, P. P. Wu, Y. S. Pu, C. J. Hsieh and B. Y. Bao, “Genetic Polymorphisms in Androgen Receptor-Binding Sites Predict Survival in Prostate Cancer Patients Receiving Androgen-Deprivation Therapy,” Annals of Oncology: Official Journal of the European Society for Medical Oncology, Vol. 23, No. 3, 2012, pp. 707-713. http://dx.doi.org/10.1093/annonc/mdr264
- B. Yu, H. Lin, L. Yang, K. Chen, H. Luo, J. Liu, X. Gao, X. Xia and Z. Huang, “Genetic Variation in the Nrf2 Promoter Associates with Defective Spermatogenesis in Humans,” Journal of Molecular Medicine, Vol. 90, No. 11, 2012, pp. 1333-1342. http://dx.doi.org/10.1007/s00109-012-0914-z
- J. Wu, M. H. Richards, J. Huang, L. Al-Harthi, X. Xu, R. Lin, F. Xie, A. W. Gibson, J. C. Edberg and R. P. Kimberly, “Human FasL Gene Is a Target of β-Catenin/T-Cell Factor Pathway and Complex FasL Haplotypes Alter Promoter Functions,” PLoS ONE, Vol. 6, No. 10, 2011, Article ID: e26143. http://dx.doi.org/10.1371/journal.pone.0026143
- M. Alam, V. Pravica, A. A. Fryer, C. P. Hawkins and I. V. Hutchinson, “Novel Polymorphism in the Promoter Region of the Human Nerve Growth-Factor Gene,” International Journal of Immunogenetics, Vol. 32, No. 6, 2005, pp. 379-382. http://dx.doi.org/10.1111/j.1744-313X.2005.00541.x
- J. C. Bryne, E. Valen, M. H. Tang, T. Marstrand, O. Winther, I. da Piedade, A. Krogh, B. Lenhard and A. Sandelin, “JASPAR, the Open Access Database of Transcription Factor-Binding Profiles: New Content and Tools in the 2008 Update,” Nucleic Acids Research, Vol. 36, Suppl. 1, 2008, pp. D102-D106. http://dx.doi.org/10.1093/nar/gkm955
- A. Sandelin, W. Alkema, P. Engstrom, W. W. Wasserman and B. Lenhard, “JASPAR: An Open-Access Database for Eukaryotic Transcription Factor Binding Profiles,” Nucleic Acids Research, Vol. 32, Suppl. 1, 2004, pp. D91-D94. http://dx.doi.org/10.1093/nar/gkh012
- A. Sandelin, W. W. Wasserman and B. Lenhard, “Consite: web-Based Prediction of Regulatory Elements Using Cross-Species Comparison,” Nucleic Acids Research, Vol. 32, Suppl. 2, 2004, pp. W249-W252. http://dx.doi.org/10.1093/nar/gkh372
- E. Pennisi, “The Biology of Genomes. Disease Risk Links to Gene Regulation,” Science, Vol. 332, No. 6033, 2011, p. 1031. http://dx.doi.org/10.1126/science.332.6033.1031
- V. Kumar, C. Wijmenga and S. Withoff, “From GenomeWide Association Studies to Disease Mechanisms: Celiac Disease as a Model for Autoimmune Diseases,” Seminars in Immunopathology, Vol. 34, No. 4, 2012, pp. 567-580. http://dx.doi.org/10.1007/s00281-012-0312-1
- L. A. Hindorff, P. Sethupathy, H. A. Junkins, E. M. Ramos, J. P. Mehta, F. S. Collins and T. A. Manolio, “Potential Etiologic and Functional Implications of GenomeWide Association Loci for Human Diseases and Traits,” Proceedings of the National Academy Sciences of the United States of America, Vol. 106, No. 23, 2009, pp. 9362- 9367. http://dx.doi.org/10.1073/pnas.0903103106
- V. Kumar, H. J. Westra, J. Karjalainen, D. V. Zhernakova, T. Esko, B. Hrdlickova, R. Almeida, A. Zhernakova, E. Reinmaa, U. Vosa, M. H. Hofker, R. S. Fehrmann, J. Fu, S. Withoff, A. Metspalu, L. Franke and C. Wijmenga, “Human Disease-Associated Genetic Variation Impacts Large Intergenic Non-Coding RNA Expression,” PLoS Genet, Vol. 9, No. 1, 2013, Article ID: e1003201. http://dx.doi.org/10.1371/journal.pgen.1003201
- B. N. Chorley, X. Wang, M. R. Campbell, G. S. Pittman, M. A. Noureddine and D. A. Bell, “Discovery and Verification of Functional Single Nucleotide Polymorphisms in Regulatory Genomic Regions: Current and Developing Technologies,” Mutation Research, Vol. 659, No. 1-2, 2008, pp. 147-157. http://dx.doi.org/10.1016/j.mrrev.2008.05.001
- L. Prokunina and M. E. Alarcon-Riquelme, “Regulatory SNPs in Complex Diseases: Their Identification and Functional Validation,” Expert Reviews in Molecular Medicine, Vol. 6, No. 10, 2004, pp. 1-15. http://dx.doi.org/10.1017/S1462399404007690
- P. R. Buckland, “The Importance and Identification of Regulatory Polymorphisms and Their Mechanisms of Action,” Biochimica et Biophysica Acta, Vol. 1762, No. 1, 2006, pp. 17-28. http://dx.doi.org/10.1016/j.bbadis.2005.10.004
- W. Sadee, D. Wang, A. C. Papp, J. K. Pinsonneault, R. M. Smith, R. A. Moyer and A. D. Johnson, “Pharmacogenomics of the RNA World: Structural RNA Polymorphisms in Drug Therapy,” Clinical Pharmacology and Therapeutics, Vol. 89, No. 3, 2011, pp. 355-365. http://dx.doi.org/10.1038/clpt.2010.314
- R. Siegel, D. Naishadham and A. Jemal, “Cancer Statistics, 2012,” CA: A Cancer Journal for Clinicians, Vol. 62, No. 1, 2012, pp. 10-29. http://dx.doi.org/10.3322/caac.20138
- W. H. Chow, S. S. Devesa, J. L. Warren and J. F. Fraumeni Jr., “Rising Incidence of Renal Cell Cancer in the United States,” JAMA, Vol. 281, No. 17, 1999, pp. 1628- 1631. http://dx.doi.org/10.1001/jama.281.17.1628
- B. Schlehofer, W. Pommer, A. Mellemgaard, J. H. Stewart, M. McCredie, S. Niwa, P. Lindblad, J. S. Mandel, J. K. McLaughlin and J. Wahrendorf, “International RenalCell-Cancer Study. VI. The Role of Medical and Family History,” International Journal of Cancer, Vol. 66, No. 6, 1996, pp. 723-726. http://dx.doi.org/10.1002/(SICI)1097-0215(19960611)66:6<723::AID-IJC2>3.0.CO;2-1
- J. Clague, J. Lin, A. Cassidy, S. Matin, N. M. Tannir, P. Tamboli, C. G. Wood and X. Wu, “Family History and Risk of Renal Cell Carcinoma: Results from a Case-Control Study and Systematic Meta-Analysis,” Cancer Epidemiology Biomarkers & Prevention, Vol. 18, No. 3, 2009, pp. 801-807. http://dx.doi.org/10.1158/1055-9965.EPI-08-0601
- M. L. Nickerson, M. B. Warren, J. R. Toro, V. Matrosova, G. Glenn, M. L. Turner, P. Duray, M. Merino, P. Choyke, C. P. Pavlovich, N. Sharma, M. Walther, D. Munroe, R. Hill, E. Maher, C. Greenberg, M. I. Lerman, W. M. Linehan, B. Zbar and L. S. Schmidt, “Mutations in a Novel Gene Lead to Kidney Tumors, Lung Wall Defects, and Benign Tumors of the Hair Follicle in Patients with the Birt-Hogg-Dube Syndrome,” Cancer Cell, Vol. 2, No. 2, 2002, pp. 157-164. http://dx.doi.org/10.1016/S1535-6108(02)00104-6
- American Cancer Society, “Cancer Facts and Figures,” 2012.
- M. Yeager, N. Orr, R. B. Hayes, K. B. Jacobs, P. Kraft, S. Wacholder, M. J. Minichiello, P. Fearnhead, K. Yu, N. Chatterjee, Z. Wang, R. Welch, B. J. Staats, E. E. Calle, H. S. Feigelson, M. J. Thun, C. Rodriguez, D. Albanes, J. Virtamo, S. Weinstein, F. R. Schumacher, E. Giovannucci, W. C. Willett, G. Cancel-Tassin, O. Cussenot, A. Valeri, G. L. Andriole, E. P. Gelmann, M. Tucker, D. S. Gerhard, J. F. Fraumeni Jr., R. Hoover, D. J. Hunter, S. J. Chanock and G. Thomas, “Genome-Wide Association Study of Prostate Cancer Identifies a Second Risk Locus at 8q24,” Nature Genetics, Vol. 39, No. 5, 2007, pp. 645- 649.
- J. E. Phillips and V. G. Corces, “CTCF: Master Weaver of the Genome,” Cell, Vol. 137, No. 7, 2009, pp. 1194- 1211. http://dx.doi.org/10.1016/j.cell.2009.06.001
- M. Tang, B. Chen, T. Lin, Z. Li, C. Pardo, C. Pampo, J. Chen, C. L. Lien, L. Wu, L. Ai, H. Wang, K. Yao, S. P. Oh, E. Seto, L. E. Smith, D. W. Siemann, M. P. Kladde, C. L. Cepko and J. Lu, “Restraint of Angiogenesis by Zinc Finger Transcription Factor CTCF-Dependent Chromatin Insulation,” Proceedings of the National Academy Sciences of the United States of America, Vol. 108, No. 37, 2011, pp. 15231-15236. http://dx.doi.org/10.1073/pnas.1104662108
Appendices
Supplemental material is available for this article.
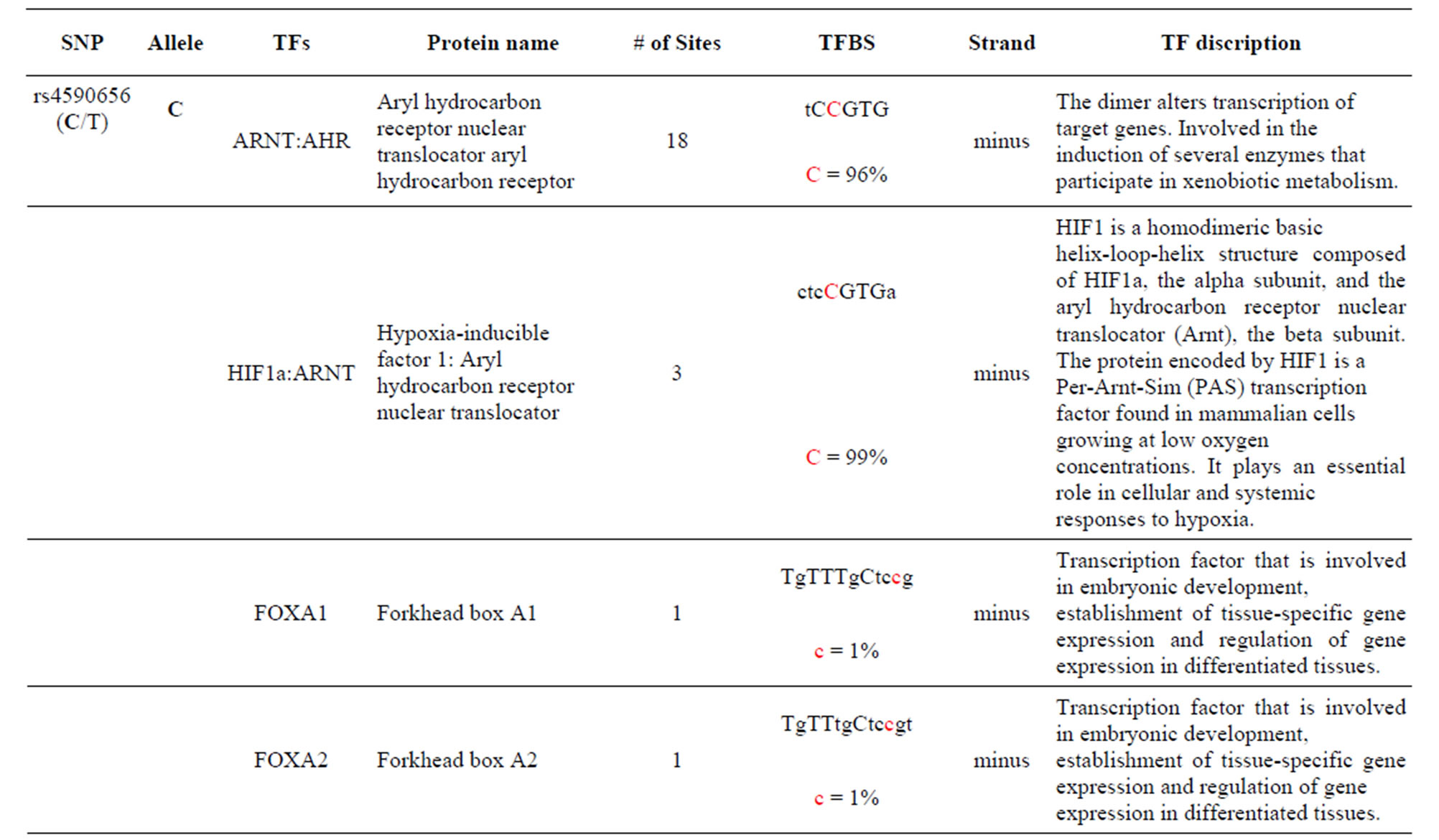
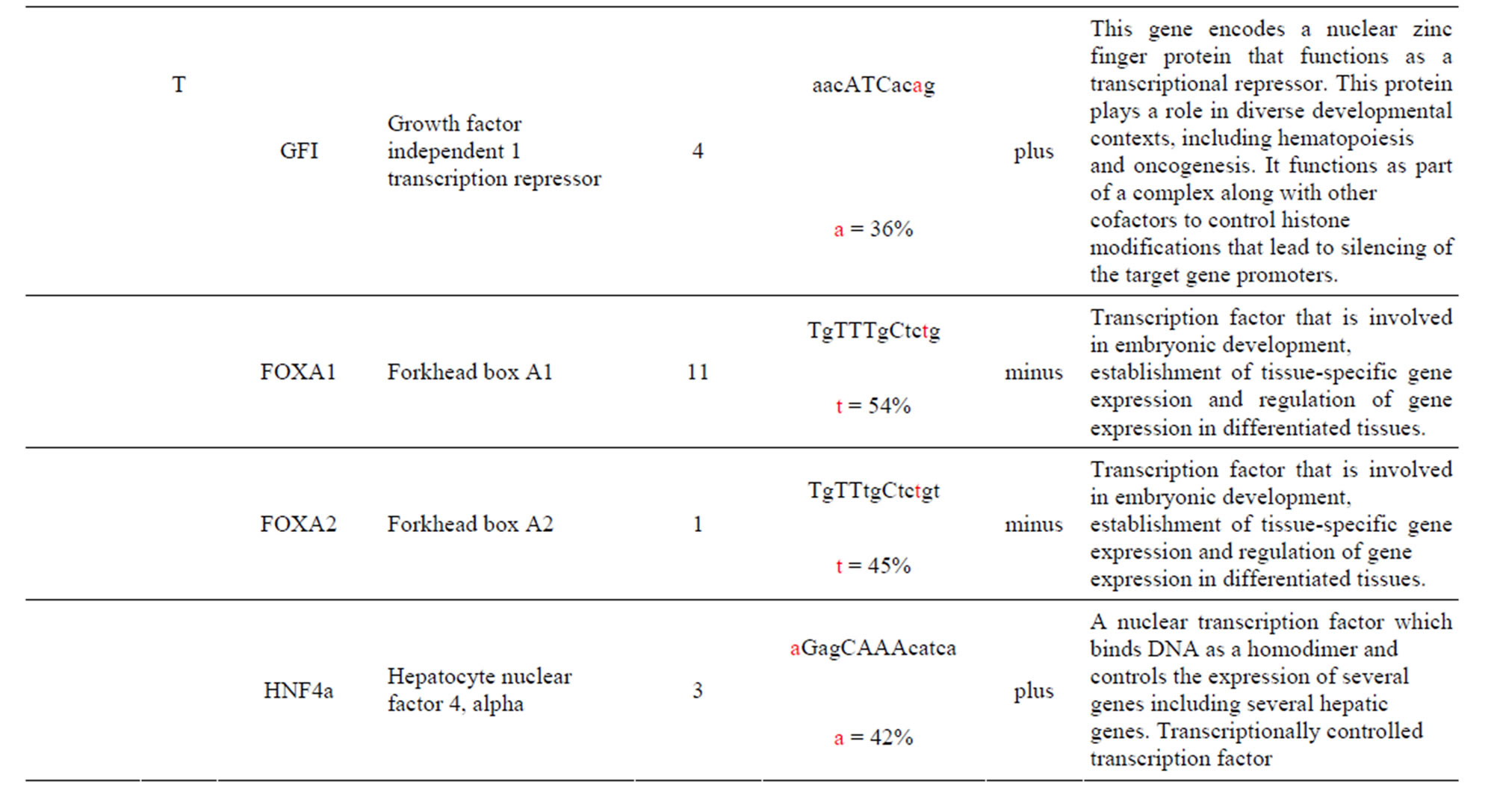

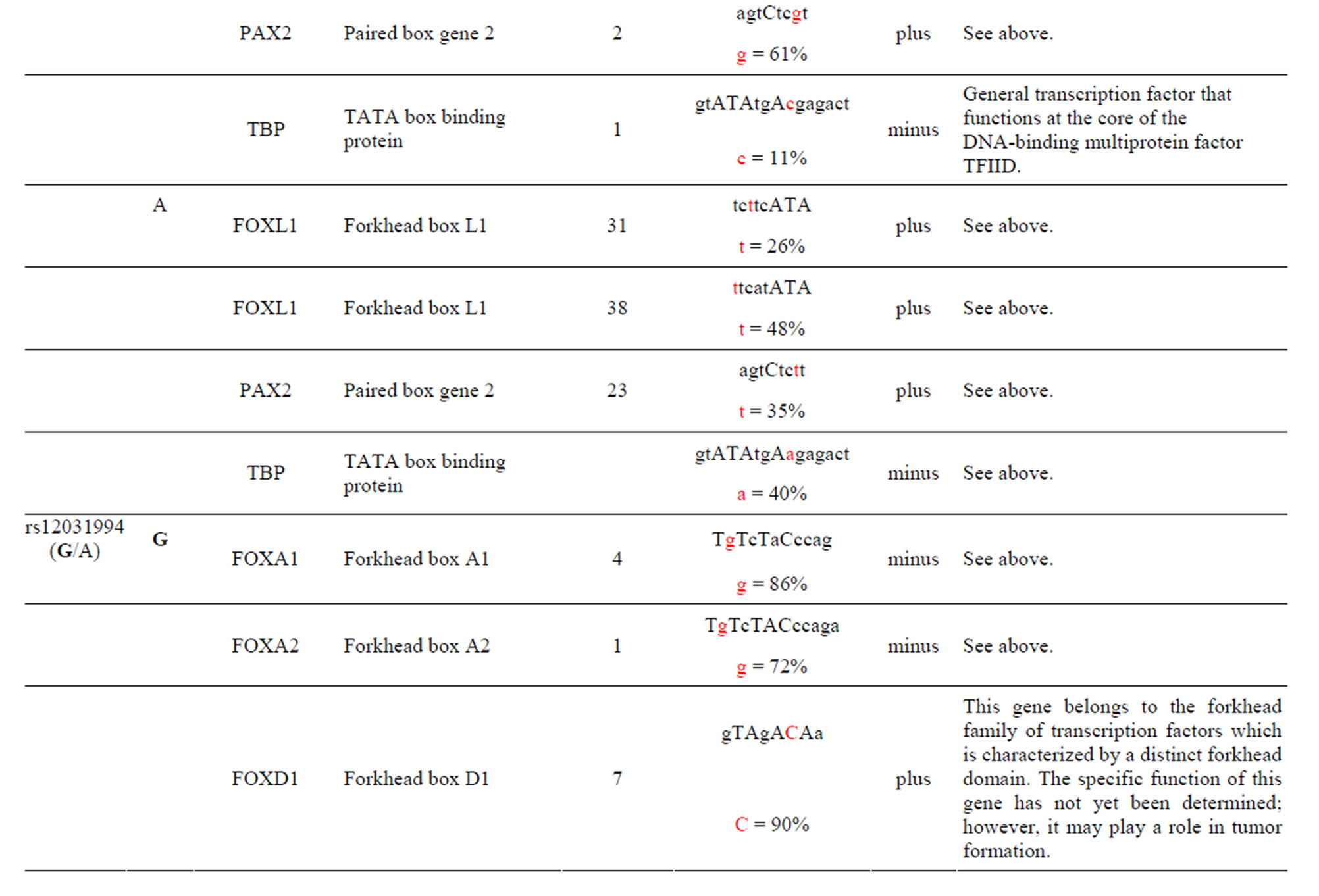

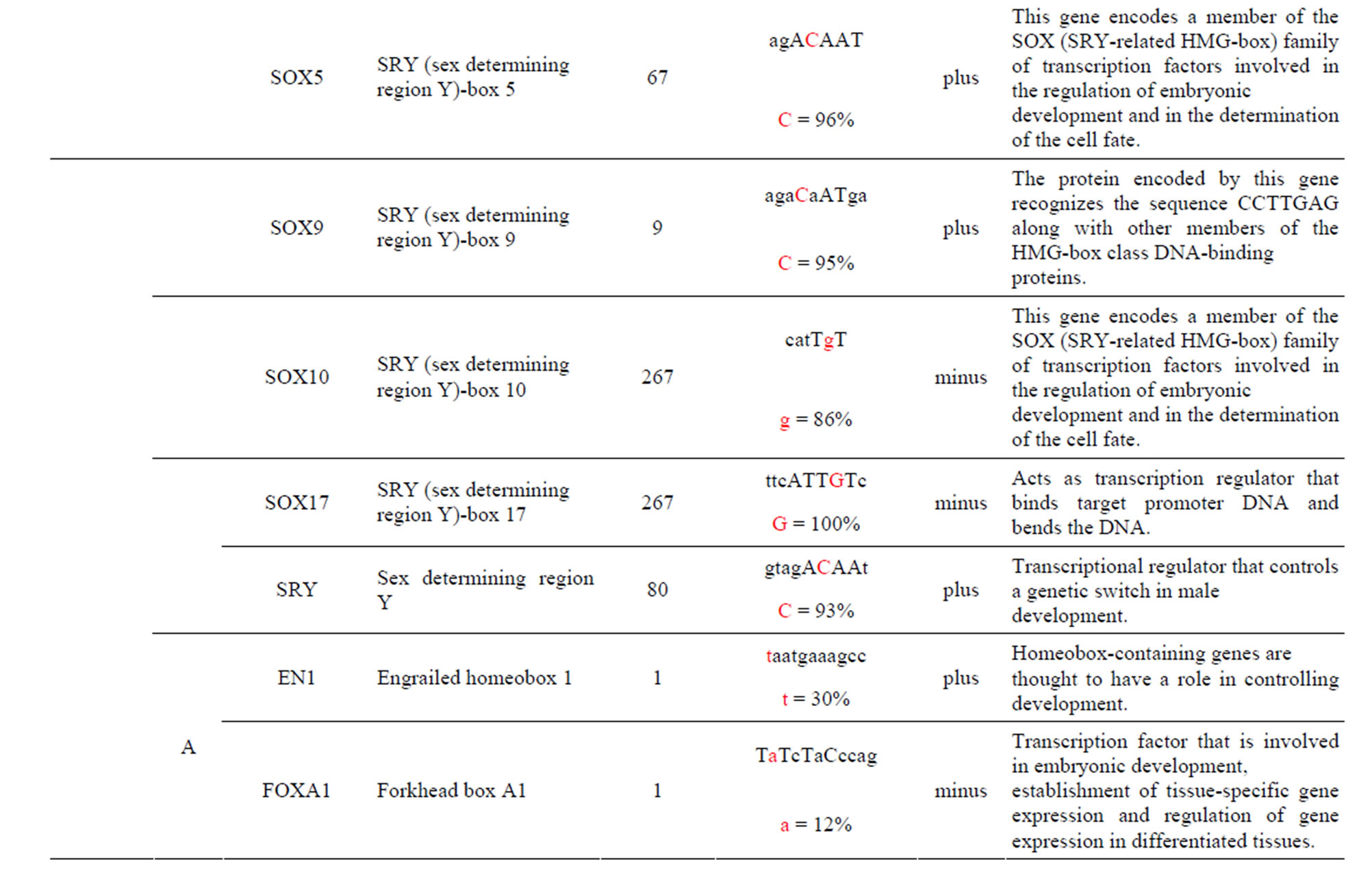
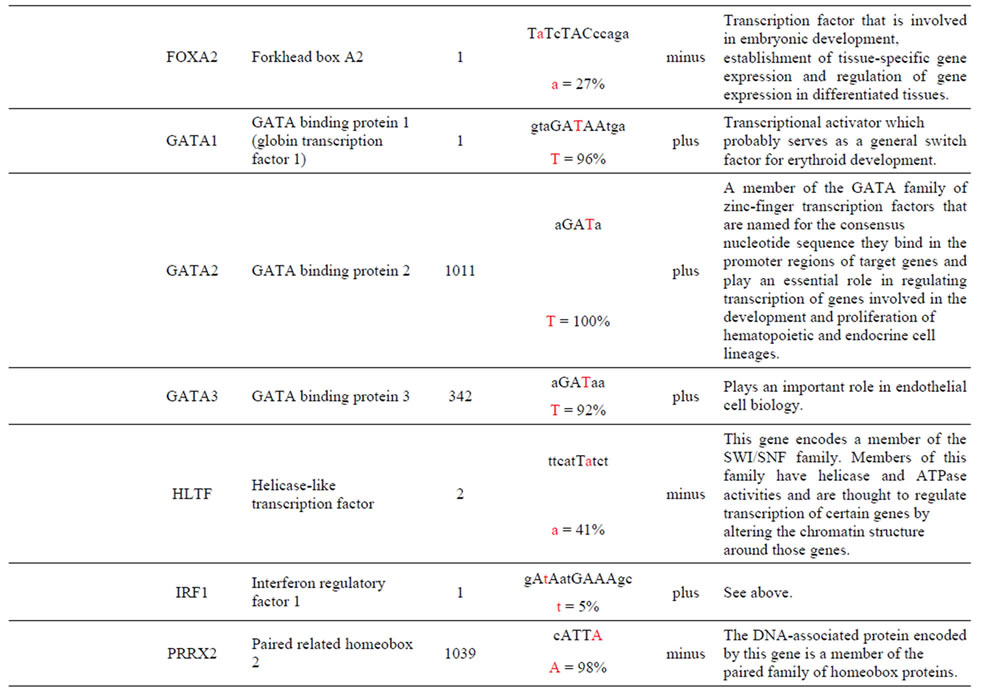

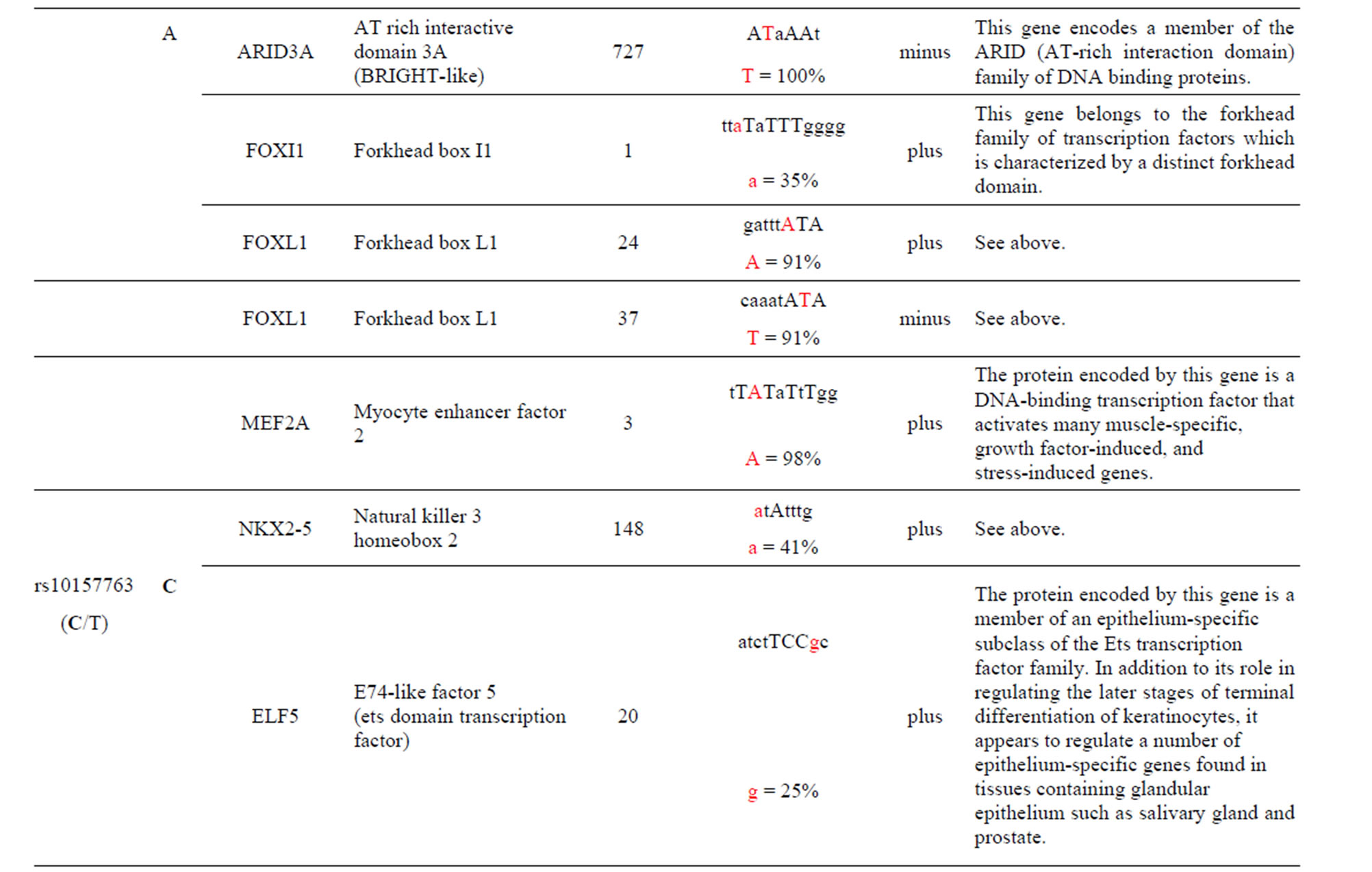
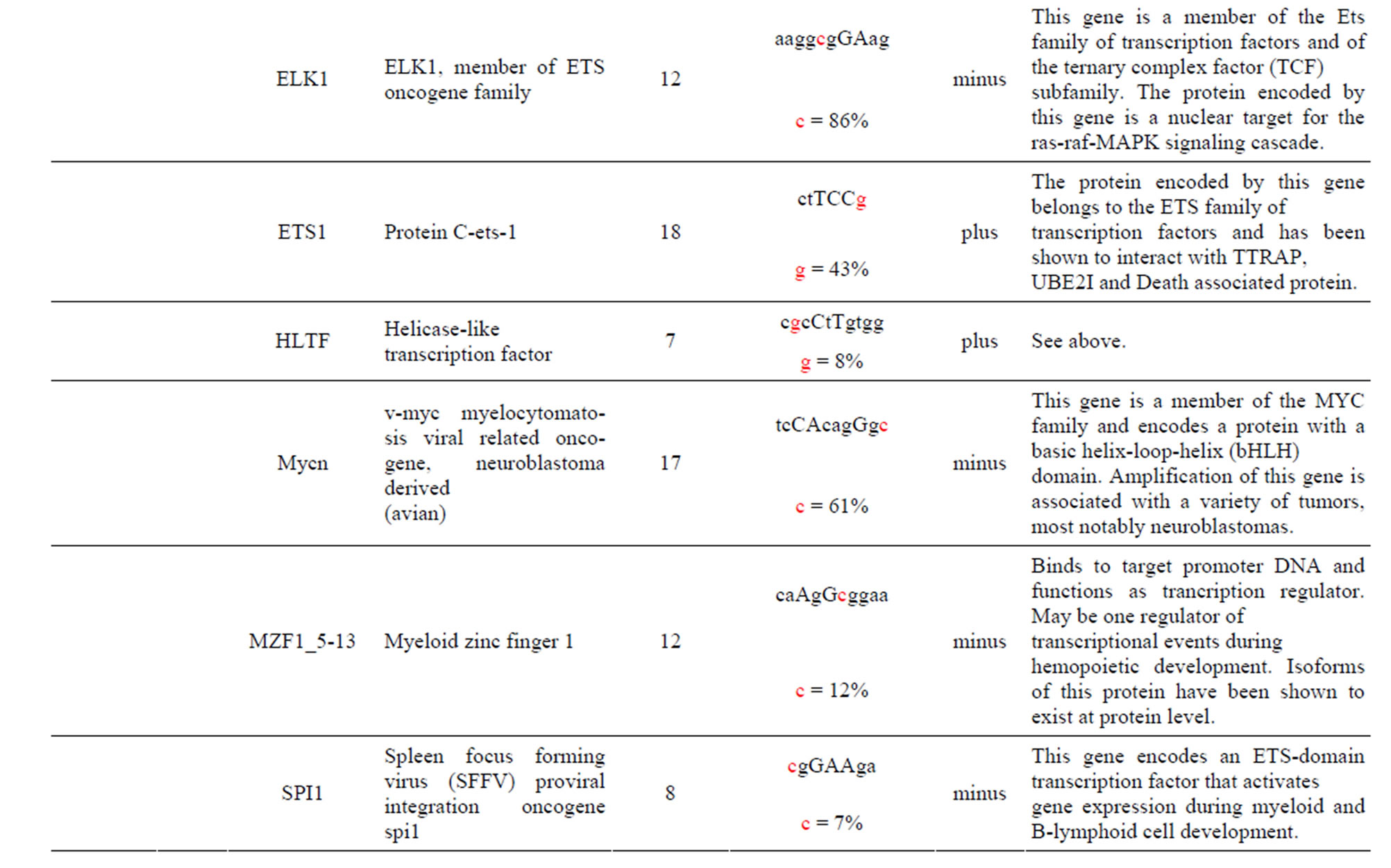

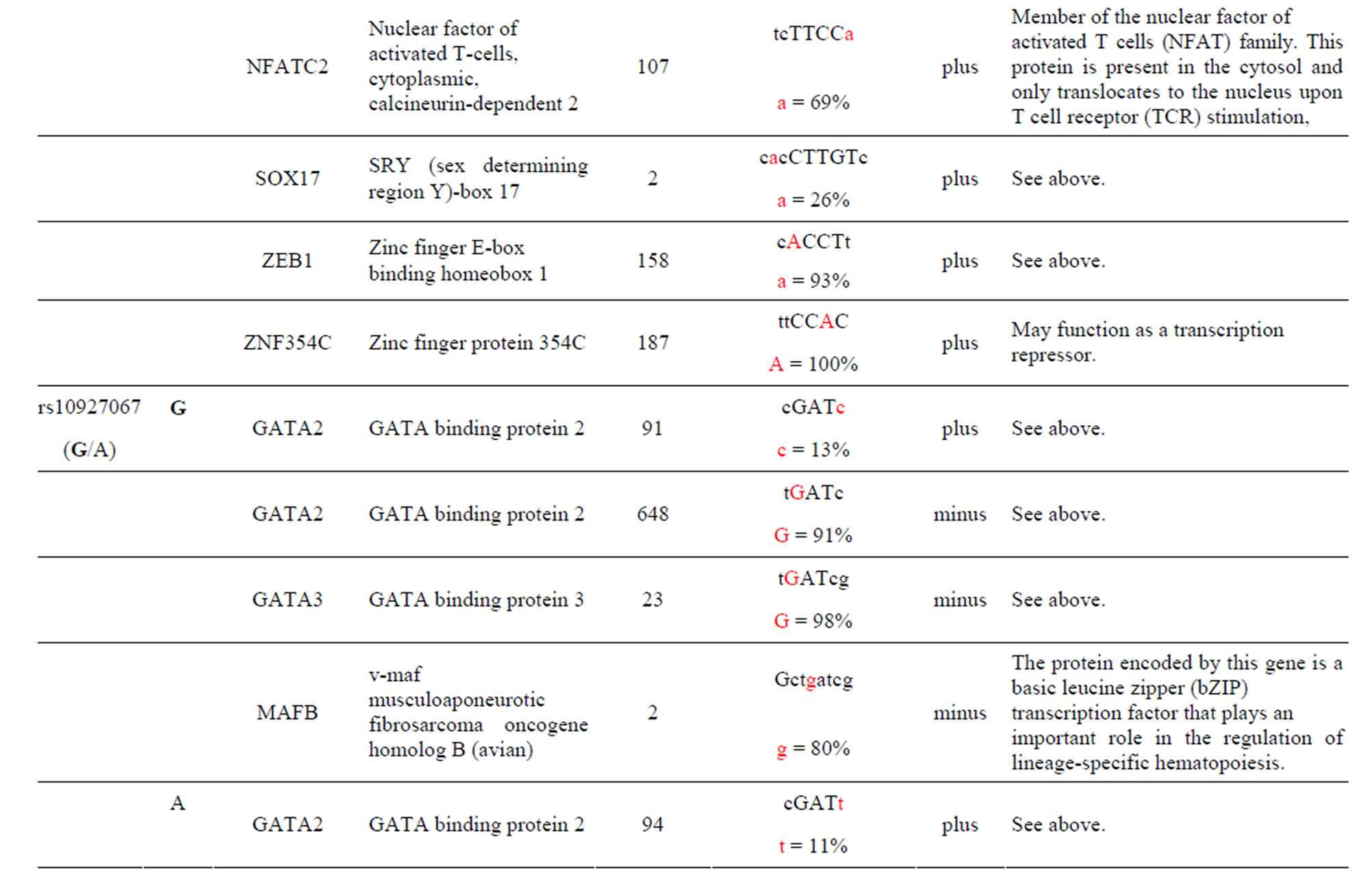
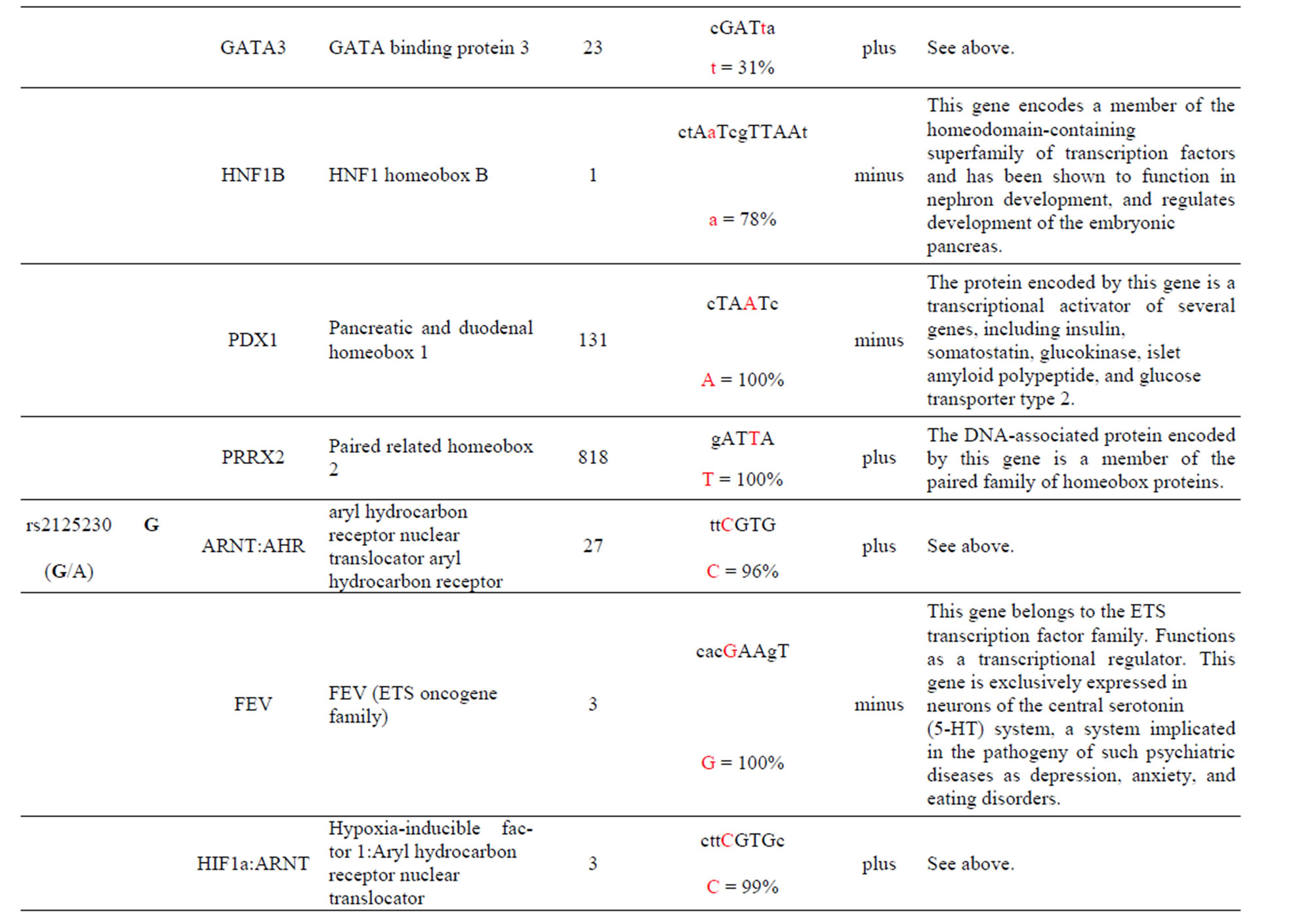
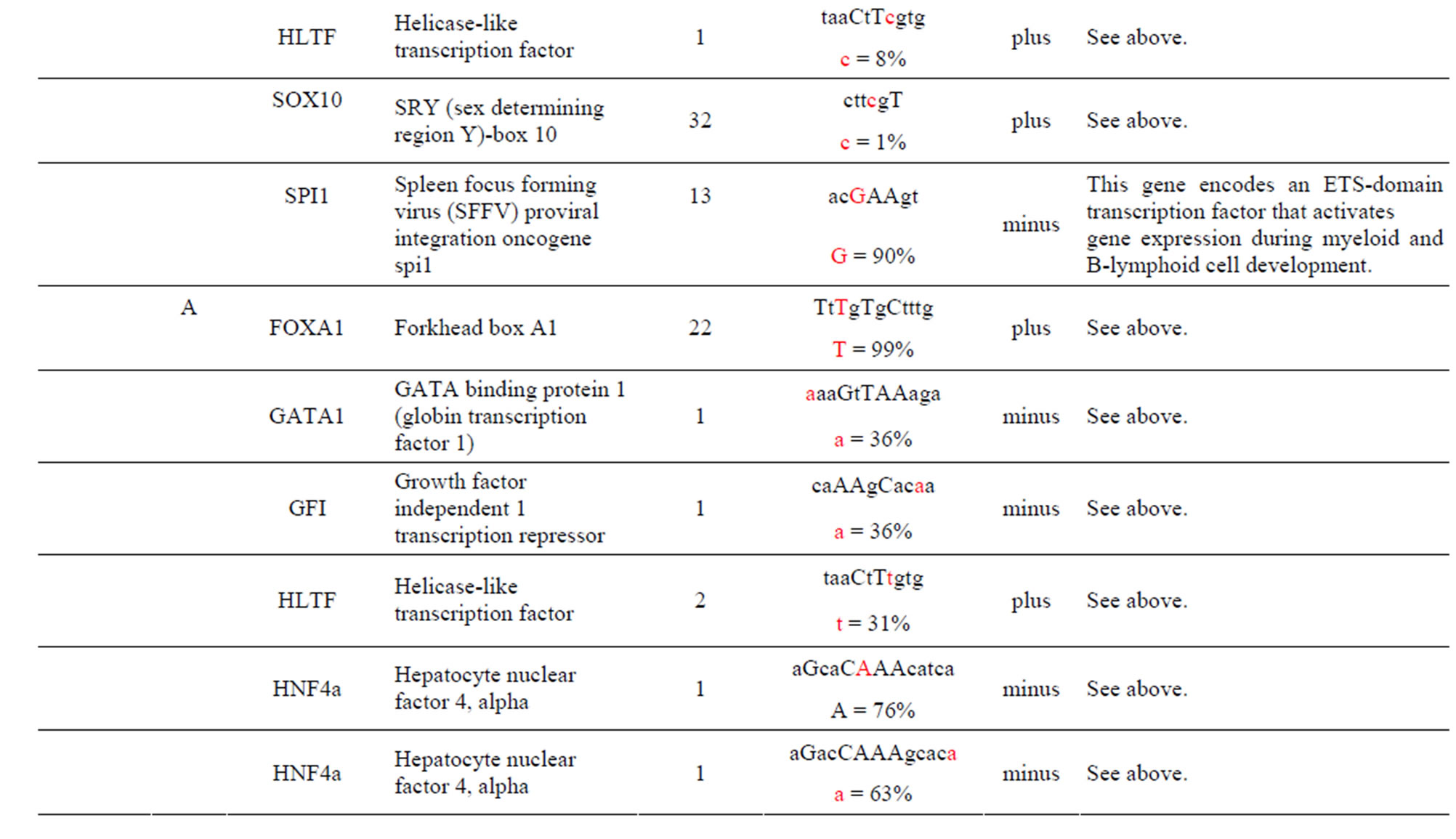
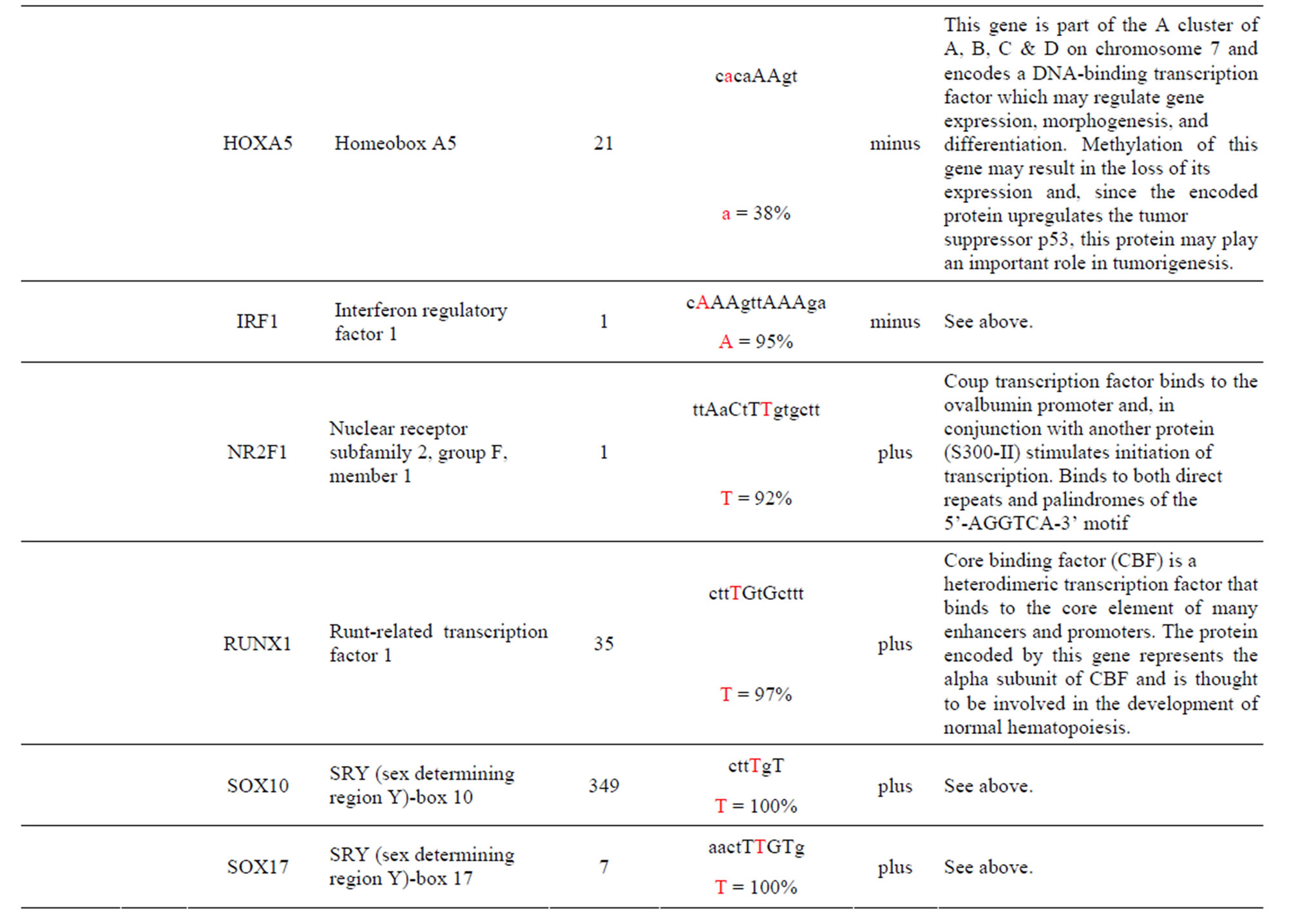
Supplement . The AKT3 SNPs from intron 1 (147,408 bp) that were examined in this study. Also listed are the transcriptional factors (TF), their potential binding sites (TFBS) containing these SNPs and DNA strand orientation. Where upper case nucleotide designates the 90% conserved BS region and red is the SNP location of the alleles in the TFBS. Below the TFBS is the nucleotide occurrence (%) obtained from the Jaspar Core database. Also listed are the number (#) of binding sites in the gene for the given TF. Note: TFs can bind to more than one nucleotide sequence.