Open Journal of Forestry
Vol.08 No.02(2018), Article ID:83205,15 pages
10.4236/ojf.2018.82012
Oil Mallee Plantings Increase Diversity of Beetles (Coleoptera) and Other Terrestrial Invertebrates within a Cropping Field
Mei Chen Leng1,2, Jonathan Majer1,2,3
1Department of Environment and Agriculture, Curtin University, Perth, Australia
2School of Biological Sciences, University of Western Australia, Crawley, Australia
3Western Australian Museum, Perth, Australia

Copyright © 2018 by authors and Scientific Research Publishing Inc.
This work is licensed under the Creative Commons Attribution International License (CC BY 4.0).
http://creativecommons.org/licenses/by/4.0/



Received: January 18, 2018; Accepted: March 19, 2018; Published: March 22, 2018
ABSTRACT
Planting oil mallee (Eucalyptus spp.) belts in cropping fields introduces a new native, perennial-based habitat type to the previously homogenous, exotic, annual-based cropping environment. The purpose of this investigation was to determine if oil mallee plantings enhance the diversity and abundance of ground-dwelling invertebrates, especially beetles, in an agro-forestry landscape by comparing four habitat types (wheat field, oil mallee belts, remnant woodland, and remnant roadside vegetation). Belts of oil mallees within a cropping paddock had substantially different assemblages of ground dwelling arthropod orders to the surrounding wheat crop. Of 93 coleopteran species trapped, 27 were found throughout all four habitats in the landscape, with the remaining species creating a strong distinction between the assemblages in the cropped areas and other habitat types in terms of species competition, diversity and abundance. Coleopteran assemblages in oil mallees were more similar to those in woodland and roadside remnants than to those in the adjacent wheat crop. Arthropod assemblages, in particular coleopterans, within this agricultural environment were thus influenced by habitat type. Oil mallee farming systems appear beneficial for arthropod conservation in the wheat belt of Western Australia.
Keywords:
Agriculture, Agroforestry, Hedgerows, Biodiversity, Conservation

1. Introduction
Agricultural impacts such as land clearing, tillage, livestock trampling and the use of pesticides and fertilizers, can make for a hostile environment for many of the invertebrates inhabiting the region (Pywell et al., 2005) . Due to crop rotation and use of land during different periods of the year, agricultural landscapes are seen as shifting mosaics of habitats for invertebrates (Holland et al., 2005) . With a better understanding of insect dynamics and distribution, their populations may be better managed and utilized to increase diversity, as well as to assist in pest control in an agro-forestry farming system (Thomas et al., 2001) .
Non-cropped areas, such as vegetation belts or blocks and field margins, are more stable and less prone to disturbance, having different vegetation composition structure than crop areas (Pywell et al., 2005) . These areas are important sites for K-strategists, which tend to be slower growing and better long-term competitors (Knox et al., 2001) . Amongst arthropods, K-strategists are rarely pests and are often beneficial (Price & Waldbauer, 1994; van Emden, 1976) . Sustaining populations of arthropods through non-cropped areas can also provide a food source for vertebrate fauna (Thomas & Marshall 1999) . A British silvopastoral study (Mcadam et al., 2007) demonstrated that the presence of trees on grasslands attracted terrestrial invertebrates and hence enhanced food supply, drawing avian fauna back into the area. By maintaining and establishing semi-natural habitats of appropriate spatial arrangement there is opportunity to create a heterogeneous mosaic of natural and semi-natural habitats and hopefully conserve biodiversity (Magagula, 2003) .
Non-cropped areas also have implications for farming practices. They can provide areas from which re-colonization can occur after adverse farming procedures, such as insecticide spraying (Mauremooto et al., 1995) . One way to maintain invertebrate biodiversity within an agro farming industry is to protect large existing vegetation remnants, whilst establishing a network of connecting corridors to promote gene flown and re-colonization (Major et al., 1999) . Joyce et al. (1999) determined that places where hedgerows and other field boundaries intersect, known as “nodes”, supported a higher number of forest carabids, probably due to nodes having higher plant species richness or providing a more stable environment. Migration of beetles within an agricultural environment between natural, semi-natural and managed habitats signifies the importance of topographically diverse habitats presented in an agricultural landscape. This is particularly significant for coleopteran recovery after disturbances (e.g., mowing, pruning, chemical treatment, etc.), coleopteran conservation and biotic diversity (Magagula, 2003) .
Western Australia (WA) currently has 4.3 million hectares affected by secondary salinity, mostly in the wheatbelt area, and this is foreseen to double within 50 years (ECITA, 2006) . Agro-forestry schemes involving native woody perennial crops are being assessed in low rainfall areas as a possible solution to salinity, and one such potential crop is oil mallee (Bell, 2005; Lefroy et al., 2005) . Oil mallees are Eucalyptus species of a multi-stemmed shrubby nature possessing lignotubers. Several species (Eucalyptus kochii ssp kochii, ssp borealis and ssp plenissima; E. horistes; E. angustissima; E. gratiae; E. loxophleba ssp lissophloia; and E. polybractea) have been selected for trials and commercial planting. Currently, around 10 million eucalypt oil mallee trees have been planted over thousands of hectares in the WA wheatbelt, arranged either in linear strips or belts (alley farming systems) or in blocks (Bell, 2005; Semple & Evans, 2004) . The original aim of this enterprise was to produce bioenergy and also eucalypt oil. Unfortunately, the bioenergy aspect has not been successfully implemented, although there is still great interest in distilling the plant material to produce valuable eucalyptus oil.
Our aim was to determine if oil mallee plantings increase the diversity and abundance of ground-dwelling arthropods, especially coleopterans, in a cropping field. We particularly focused on Coleoptera due to their high species richness and ecological (niche) diversity (Grove & Stork, 2000) . We sought to compare both ordinal arthropod and beetle species data from pitfall trapping within four habitat types (wheat field, oil mallee belts, remnant woodland, and remnant roadside vegetation) and to observe temporal variation within these habitats.
2. Materials and Methods
The study was conducted on a farm in the Cuballing region of the central wheatbelt of WA (32˚49'S, 117˚10'E). The site consisted of a 72 ha wheat field and 37 ha of adjacent remnant native woodland (Figure 1). The field is bounded by roads on two sides and strips of remnant vegetation approximately 5 m wide persist between the field edge and the road. The woodland was historically grazed by sheep but has been fenced off for the last 10 years to encourage natural regeneration. The field was developed and cleared around 1930 with a series of contour banks and dams installed in the late 1950’s to control erosion and catch water runoff. Linear strips of oil mallees (four tree rows wide) were planted along these contour lines in 1999. Two species of mallee trees were planted, Eucalyptus loxophleba ssp lissophloia (York Gum Mallee), native to the south-western Australia and a small proportion of E. polybractea (Blue Mallee), native to south-eastern Australia.
The field had been continuously cropped in recent years in a rotation of cereals, canola and lupins. When the study commenced there had been no spraying of pesticides since 2003, when Cypermethrinâ had been used as a blanket application for invertebrate pests. In the previous year (2002), Bifenthrinâ had been used for red-legged earth mite (Halotydeus destructor) control in the canola crop. At the beginning of 2006, there were sheep in the field to graze the crop stubbles but these were removed 2 weeks before sampling began. The surrounding agricultural lands were cultivated with cereal crops such as barley, oats (mixed with livestock grazing) and wheat.
Wet pitfall traps were installed throughout the field and adjacent landscape to sample ground invertebrates in the four habitat types (Figure 1). Pitfall traps consisted of plastic vials that were 45 mm internal diameter and 85 mm in depth. They were placed at even intervals within habitat types in groups of 10, flush to

Figure 1. Aerial photograph of study site where oil mallee trees had been planted along contours. White lines denote the boundary fence and dots indicate pitfall trap positions. Habitats are coded accordingly; rem = woodland; road = roadside vegetation; M1 - M4 = oil mallees; P1 - P4 = wheat field.
ground level and sealed. The following day, the lids were removed and the traps were filled one third full with a mixture of ethanol-glycerol and left open for a week. Samples were taken once in the dry season before cropping commenced (02-09 April 2006), and once in the wet season after the crop had established (19-26 August 2006). Locations of the pitfall traps were recorded using a GPS to ensure that the second sampling occurred in the same location as the first and to obtain location measurements of the traps.
Invertebrate samples were cleaned through a 0.5 mm sieve and stored in 70% ethanol prior to being sorted to ordinal level under a compound microscope using a dichotomous key (Harvey & Yen 1997) . The coleopterans were further sorted to morphospecies using Naumann et al. (1991) and then identified, where possible, to species level with the assistance of a local coleopteran taxonomist.
One-way analysis of variance (ANOVA) was conducted using the statistical package SPSS 14.0 (2005) on the ordinal data to compare the numbers within the four habitats. A Bonferroni post-hoc test was performed to determine where the significance, if any, lay. Orders that occurred in less than and equal to 10 traps were removed before the test was conducted and the data were log-transformed. The Kruskal-Wallis non-parametric test was conducted on orders that had non-normal distribution due to small sample size.
Ordinations were conducted using PRIMER v6 (2006) , on ordinal and beetle species data to determine if there were any characteristic differences in the assemblages found within the different habitats. A Bray-Curtis (1957) similarity matrix was constructed on square root-transformed data. To assist in interpretation, a non-metric multi-dimensional scaling (nMDS) test was performed (50 restarts) to determine community compositions (Clarke & Gorley, 2006) . Analysis of similarity (ANOSIM) was carried out to determine the degree of similarity (999 permutations). Further testing was conducted on the beetle data using PC-ORD 4.30 (McCune & Mefford, 1999) to determine if there were any indicator species, by running a two-way indicator species analysis (TWINSPAN) test (Hill, 1979) . To ensure that the data were not skewed, any beetle species occurring in two or fewer samples were removed.
Species richness was estimated using the software EstimateS v8.2.0 (Colwell, 2006) to calculate the Michaelis-Mention (Colwell & Coddington, 1994) curve-fit estimate of species richness. The estimation of total species was taken from equal numbers of samples (10) at 1000 randomizations.
3. Results
3.1. Ordinal Data
The orders Acarina, Araneae, Coleoptera, Diptera, Hymenoptera and Psocoptera were the most abundant taxa across the two seasons, showing high means across all four different habitats (Appendix 1). The ANOVA showed that certain taxa trapped in April (Araneae, Acarina, Thysanoptera, Psocoptera, Diptera, Coleoptera and Hymenoptera) exhibited statistically significant differences between habitat types. Wheat field habitat sites P1, P2 and to a lesser extent P3, were identified as having significantly higher values of Coleoptera than in the other habitats (P < 0.001). Four taxa in August (Chilopoda, Psocoptera, Diptera and Coleoptera) exhibited statistically significant differences between habitat types. Wheat field habitat sites were identified as having significantly greater numbers of Coleoptera than in other habitats (P < 0.001).
The nMDS ordination indicated that ordinal composition differed between the four habitat types (P < 0.001) with a clear separation between the wheat field and oil mallees assemblages and a partial separation of the woodland. The roadside habitat seemed to be intermediate between the oil mallees and woodland, although some oil mallee samples were closely aligned with the woodland (Figure 2).
3.2. Coleoptera Species Composition
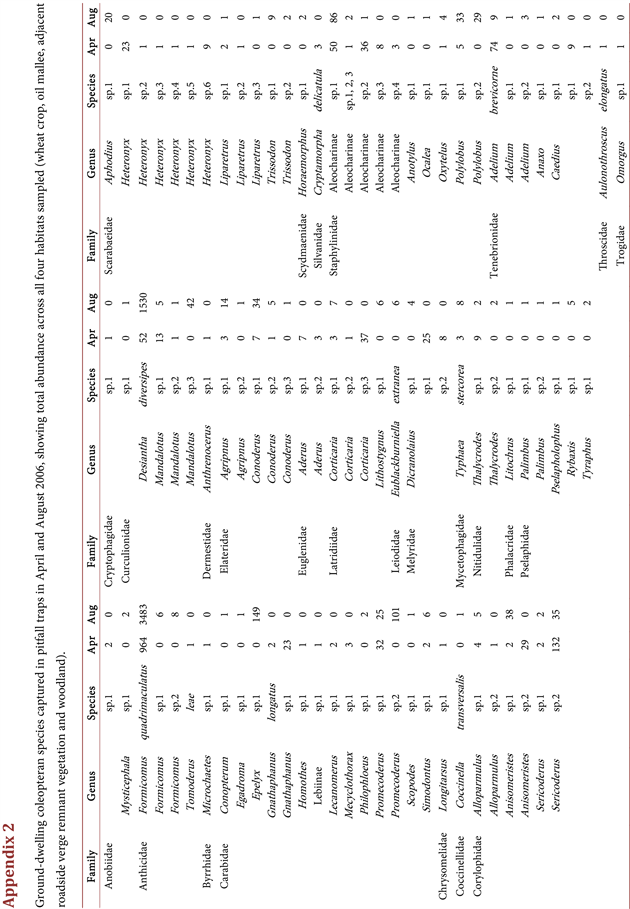
A total of 93 Coleopteran species was trapped over the two sampling periods and, of these, 23 species were present in both sampling periods, with a general trend of greater abundance during the August sampling period. April had a total of 57 coleopteran species collected, whilst in August 59 species were collected (Appendix 2). The ordination of coleopterans from April and August provided very similar results, so data have been combined (Figure 3). There was clear segregation between the coleopteran species assemblages in the different habitat types, particularly the wheat field from the oil mallees and woodland habitats. From Figure 3, three groups can be derived: wheat, mallee and roadside/woodland (P < 0.001).

Figure 2. Ordination of ground-dwelling invertebrate assemblages identified to ordinal level. Each data point represents data collected from a pitfall trap that was placed in one of the four habitat types and left open for one week during the dry season (April 2006) and one week during the wet season (August 2006).

Figure 3. Ordination of ground-dwelling coleopteran assemblages identified to species level. Each data point represents data collected from a pitfall trap that was placed in one of the four habitat types and left open for one week during the dry season (April 2006) and one week during the wet season (August 2006).
The TWINSPAN (Table 1) analysis indicates that the coleopteran species can be divided into three groups; oil mallees, woodland/roadside plus field (P4) and the wheat field. A total of 18 indicator species were derived, with 10 in the mallees (Promecoderus sp.2, (Carabidae) Sericoderus sp.2 (Corylophidae), Mandalotus sp.1 and sp.2 (Curculionidae), Aderus sp.1 (Aderidae), Lithostygnus sp.1 (Lathridiidae), Eublackburniella extranea (Leiodidae), Melyridae sp.1, Heteronyx sp.1 (Scarabaeidae) and Aleocharinae sp.2 (Staphylinidae)), five in the woodland/roadside and P4 (Formicomus quadrimaculatus, Formicomus sp.1, Formicomus sp.2 (Anthicidae), Epelyx sp.1 (Carabidae) and Aleocharinae sp.1 (Staphylinidae)) and three in the remaining wheat field habitat sites of P1, P2 and P3 (Ganthaphanus sp1 (Carabidae) and Conoderus sp.1 (Elateridae)).
Species richness was estimated using EstimateS from the total species captured from 10 samples in each habitat type. Michaelis-Menten estimate of asymptote showed that the roadside had slightly higher species richness followed by oil mallee, woodland and wheat field (Table 2).
4. Discussion
A total of 19 arthropod orders were trapped over the two seasons and all orders showed a consistent presence within the roadside and oil mallee habitat. Invertebrate abundance was slightly higher at the end of the dry season (April) than during the winter growing season (August). Three major orders, Araneae, Coleoptera and Hymenoptera showed variation in trends according to seasons.
Araneae were found to favour the woodland habitat during the dry season, whilst in the wet season they appeared to be distributed throughout the other habitats (wheat field, oil mallees and roadside) more evenly. These findings are similar to British studies (Maudsley et al., 2002; Pywell et al., 2005) , which showed that Araneae favoured shrubby hedgerow foliage and vegetation during the winter periods. In the Mediterranean-type environment of this study, woodland vegetation may be used as a summer refuge when there is very little vegetation in the wheat field and their activity is high. On the whole, results were similar to another British study (Mcadam et al., 2007) , suggesting that Araneae prefer woodland vegetation. This may be due to the greater vegetation complexity (species diversity and structure) found in woodland compared to the other three habitats.
It was interesting to note that coleopterans seemed to favour the wheat field area during both seasons. Duelli et al. (1990) found similar results when examining arthropod movements in cultivated and natural areas. Carabids and staphylinids were shown to be associated with crop fields and surrounding grass strips. They concluded that these grass strip habitats were the source of diversity, which influences arthropod distribution in the crop and semi-natural areas. There were 27 Coleoptera species trapped across all the sampled habitat types, indicating that a majority of the beetle species have a cosmopolitan distribution. There appears to be three main clusters of Coleoptera species found in this landscape (wheat field, oil mallee and woodland/roadside-P4 (Figure 3),

Table 1. TWINSPAN of coleopteran species (combined April and August samples).
Indicator species exhibiting significance are shown in bold **P < 0.05. Group clustering denoted by: Mallee; Remnant/Road plus P4; and P1-3.
Table 2. Coleopteran species richness estimates derived from Michaelis-Menten asymptote values extrapolated from 10 samples using EstimateS.
*M-M Michaelis-Menten.
indicating that the nature of the habitat is determining beetle distributions. A similar result was found by Vanbergen et al. (2005) working on carabid communities in a heterogeneous landscape in Britain. Dominant Coleoptera groups, such as those from the Carabidae, Scarabaeidae, Melyridae and Tenebrionidae families, have been associated with Eucalyptus and Acacia vegetation communities (Austin et al., 2004) . Species from these families captured in this study were found to have resided in the woodland, roadside and oil mallee habitats, which were both dominated by Eucalyptus species.
When comparing seasons, the majority of coleopteran species (29) were found in the oil mallee during the dry late autumn (April). However, in the wet season (August) there was a shift in composition to a more even spread of coleopteran species (23) in the oil mallees and wheat field. The large numbers of Coleoptera caught in the wheat field may be due to species movement into more productive areas. There is some evidence of this between the two seasons, with some species moving from the oil mallees during the dry season into the wheat field in the wet season, or occurring in all habitats in the dry season to only being concentrated in the wheat field during the wet season (e.g., Sericoderus sp.1 (Corylophidae), Agripnus sp.1 (Elateridae) and Formicomus quadrimaculatus (Anthicidae)). Other species were found to occur only in the oil mallee and woodland habitats in the dry season, but dispersed throughout all the habitats in the wet season (e.g., Simodontus sp.1 (Carabidae), Conoderus sp.2 (Elateridae), Aleocharinae sp.1 (Staphylinidae) and Polylobus sp.1 (Staphylinidae)). This type of movement could explain why field site P4 unexpectedly had similar coleopteran assemblages to woodland and roadside. This could also explain why there was higher estimated species richness for the roadside and oil mallee, suggesting that there might be woodland-type and wheat field type fauna existing in these two habitats.
Of the coleopteran species trapped, Coccinella transversalis (Transverse ladybird) was the only known ‘beneficial’ identified. It is a significant predator of aphids and to lesser extent scale insect crawlers. Pest species collected included Desiantha diversipes (Desiantha weevil), a sporadic pest of wheat/cereal seedlings and Adelium brevicorne (Bronzed field beetle), whose larvae can cause damaged to canola seedlings. Of lesser note are Typhaea stercorea (Hairy fungus beetle) and Corticaria species, which do not cause damage to stored grain, but have an impact on its market value. Further investigations are needed to determine whether the remaining coleopteran species are of benefit or detriment to agricultural production.
Over the two seasons, Hymenoptera were consistently present in higher abundance in the roadside and oil mallee habitats, suggesting that this is their preferred habitat. Their occurrence in the wheat field and woodland may be due to wide diversity within this order, giving them an ability to disperse into a wide range of habitats. Alternatively, a spill-over effect could be occurring where there is ‘leakage’ into the adjacent habitats. Dangerfield et al. (2003) found that invertebrates were either found across all habitats, or mostly concentrated in a particular habitat, but gradually leaked across to other habitats over distances up to 400 m. A spill-over effect occurs in areas where species come from a low productivity area into a high productivity area (Rand et al., 2006) . In other words, species will move from natural or semi-natural habitat into high productivity areas, such as a crop field, and vice versa once the crop has been harvested.
The planting of oil mallee belts in this landscape appears to be beneficial for the conservation of some arthropods. The transformation from a homogenous to a heterogeneous landscape has facilitated a more diverse arthropod distribution. This has allowed orders normally found in remnant habitat to return to the field area, as well as providing refuge sites during seasonal adversity. While some coleopteran species were found throughout all four habitats, there is a strong distinction between the assemblages in the oil mallees and wheat field. This signifies the importance of retaining and enhancing the topographically diverse habitats presented in agricultural landscapes. An understanding of this diversity is important in ecological conservation, in particular the preservation of biodiversity.
Acknowledgements
We thank Susan and Mike Hesford for allowing this study to be conducted on their property and for their generous hospitality and assistance. Patrick Smith, formerly of CSIRO Sustainable Ecosystems, provided considerable assistance in setting up this study. We are also grateful to Andras Szito, entomologist and curator of the insect collection at the WA Department of Agriculture and Food for his expertise and assisting with taxonomic identification of beetles.
Cite this paper
Leng, M. C., & Majer, J. (2018). Oil Mallee Plantings Increase Diversity of Beetles (Coleoptera) and Other Terrestrial Invertebrates within a Cropping Field. Open Journal of Forestry, 8, 167-181. https://doi.org/10.4236/ojf.2018.82012
References
- 1. Austin, A. D., Yeates, D. K., Cassis, G., Fletcher, M. J., La Salle, J., Lawrence, J. F., McQuillan, P. B., Mound, L. A., Bickel, D. J., Gullan, P. J., Hales, D. F., & Taylor, G. S. (2004). Insects “Down Under”—Diversity, Endemism and Evolution of the Australian Insect Fauna: Examples from Selected Orders. Australian Journal of Entomology, 43, 216-234. https://doi.org/10.1111/j.1326-6756.2004.00448.x [Paper reference 1]
- 2. Bell, S. J. (2005). Constructing Sustainable Rural Landscapes: Oil Mallees and the Western Australian Wheatbelt. Geographic Research, 43, 194-208.https://doi.org/10.1111/j.1745-5871.2005.00313.x [Paper reference 2]
- 3. Bray, J. R., & Curtis, J. T. (1957). An Ordination of the Upland Forest Communities of Southern Wisconsin. Ecological Monographs, 27, 325-349.https://doi.org/10.2307/1942268 [Paper reference 1]
- 4. Clarke, K. R., & Gorley, R. N. (2006). Primer v6: User Manual. Primer-E, Plymouth. [Paper reference 1]
- 5. Colwell, R. K. (2006). Estimate S, Version 8.0: Statistical Estimation of Species Richness and Shared Species from Samples (Software and User’s Guide). http://viceroy.eeb.uconn.edu/estimates [Paper reference 1]
- 6. Colwell, R. K., & Coddington, J. A. (1994). Estimating Terrestrial Biodiversity through Extrapolation. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 345, 101-118. https://doi.org/10.1098/rstb.1994.0091 [Paper reference 1]
- 7. Dangerfield, J. M., Pik, A. J., Britton, D., Holmes, A., Giligns, M., Oliver, I., Briscoe, D. & Beattie, A. J. (2003). Patterns of Invertebrate Biodiversity across a Natural Edge. Austral Ecology, 28, 227-236. https://doi.org/10.1046/j.1442-9993.2003.01240.x [Paper reference 1]
- 8. Duelli, P., Studer, M., Marchand, I., & Jakob, S. (1990). Population Movements of Arthropods between Natural and Cultivated Areas. Biological Conservation, 54, 193-207.https://doi.org/10.1016/0006-3207(90)90051-P [Paper reference 1]
- 9. ECITA (2006). Living with Salinity—A Report on Progress, the Extent and Economic Impact of Salinity in Australia. Senate Environment, Communications, Information Technology and the Arts References Committees (ECITA). [Paper reference 1]
- 10. Grove, S. J., & Stork, N. E. (2000). An Inordinate Fondness for Beetles. Invertebrate Taxonomy, 14, 733-739. https://doi.org/10.1071/IT00023 [Paper reference 1]
- 11. Harvey, M. S., & Yen, A. L. (1997). Worms to Wasps: An Illustrated Guide to Australia’s Terrestrial Invertebrates. Melbourne: Oxford University Press. [Paper reference 1]
- 12. Hill, M. O. (1979). TWINSPAN: A FORTRAN Program for Arranging Multivariate Data in an Ordered Two-Way Table by Classification of the Individuals and Attributes. Ithaca, NY: Cornell University. [Paper reference 1]
- 13. Holland, J. M., Thomas, C. F. G., Birkett, T., Southway, S., & Oaten, H. (2005). Farm-Scale Spatiotemporal Dynamics of Predatory Beetles in Arable Crops. Journal of Applied Ecology, 42, 1140-1152. https://doi.org/10.1111/j.1365-2664.2005.01083.x [Paper reference 1]
- 14. Joyce, K. A., Holland, J. M., & Doncaster, C. P. (1999). Influences of Hedgerow Intersections and Gaps on the Movement of Carabid Beetles. Bulletin of Entomological Research, 89, 523-531. https://doi.org/10.1017/S000748539900067X [Paper reference 1]
- 15. Knox, B. R., Ladiges, P., Evans, B., & Saint, R. (2001). Biology. Sydney: McGraw-Hill. [Paper reference 1]
- 16. Lefroy, E. C., Flugge, F., Avery, A., & Hume, I. (2005). Potential of Current Perennial Plant-Based Farming Systems to Deliver Salinity Management Outcomes and Improve Prospects for Native Biodiversity: A Review. Australian Journal of Experimental Agriculture, 45, 1357-1367. https://doi.org/10.1071/EA04160 [Paper reference 1]
- 17. Magagula, C. N. (2003). Changes in Carabid Beetle Diversity within a Fragmented Agricultural Landscape. African Journal of Ecology, 41, 23-30.https://doi.org/10.1046/j.1365-2028.2003.00403.x [Paper reference 2]
- 18. Major, R. E., Smith, D., Cassis, G., Gray, M., & Colgan, D. J. (1999). Are Roadside Strips Important Reservoirs of Invertebrate Diversity? A Comparison of the Ant and Beetle Faunas of Roadside Strips and Large Remnant Woodlands. Australian Journal of Zoology, 47, 611-624. https://doi.org/10.1071/ZO99048 [Paper reference 1]
- 19. Maudsley, M., Seeley, B., & Lewis, O. (2002). Spatial Distribution Patterns of Perdatory Arthropods within an English Hedgerow in Early Winter in Relation to Habitat Variables. Agriculture, Ecosystems and Environment, 89, 77-89.https://doi.org/10.1016/S0167-8809(01)00320-6 [Paper reference 1]
- 20. Mauremooto, J. R., Wratten, S. D., Worner, S. P., & Fry, G. L. A. (1995). Permeability of Hedgerows to Predatory Carabid Beetles. Agriculture, Ecosystems and Environment, 52, 141-148. https://doi.org/10.1016/0167-8809(94)00548-S [Paper reference 1]
- 21. Mcadam, J. Sibbald, A., Teklehaimanot, Z., & Eason, W. (2007). Developing Silvopastoral Systems and Their Effects on Diversity of Fauna. Agroforestry Systems, 70, 81-89.https://doi.org/10.1007/s10457-007-9047-8 [Paper reference 2]
- 22. McCune, B., & Mefford, M. J. (1999). PC-ORD: Multivariate Analysis of Ecological Data. Version 4.30. Gleneden Beach, OR: MjM Software. [Paper reference 1]
- 23. Naumann, I. D. et al. (1991). The Insects of Australia: A Textbook for Students and Research Workers. Volumes 1 & 2. Carlton, VIC: Melbourne University Press. [Paper reference 1]
- 24. Price, P. W., & Waldbauer, G. P. (1994). Ecological Aspects of Pest Management. In R. L. Metcalf, & W. H. Luckmann (Eds.), Introduction to Insect Pest Management, New York: John Wiley & Sons, Inc. [Paper reference 1]
- 25. PRIMER-E (2006). Primer 6 for Windows. Plymouth: PRIMER-E Ltd. [Paper reference 1]
- 26. Pywell, R. F., James, K. L., Herbert, I., Meek, W. R., Carvell, C.., Bell, D., & Sparks, T. H. (2005). Determinants of Overwintering Habitat Quality for Beetles and Spiders on Arable Farmland. Biological Conservation, 123, 79-90. https://doi.org/10.1016/j.biocon.2004.10.010 [Paper reference 3]
- 27. Rand, T. A., Tylianakis, J. M., & Tscharntke, T. (2006). Spillover Edge Effects: The Dispersal of Agriculturally Subsidized Insect Natural Enemies into Adjacent Natural Habitats. Ecology Letters, 9, 603-614. https://doi.org/10.1111/j.1461-0248.2006.00911.x [Paper reference 1]
- 28. Semple, K. E., & Evans, P. D. (2004). Wood-Cement Composites—Suitability of Western Australian Mallee Eucalypt, Blue Gum and Melaeucas. A Report for RIRDC/Land & Water Australia/FWPRDC/MDBC. [Paper reference 1]
- 29. SPSS (2005). SPSS for Windows, Rel. 14.0. Chicago: SPSS Inc. [Paper reference 1]
- 30. Thomas, C. F. G., & Marshall, E. J. P. (1999). Arthropod Abundance and Diversity in Differently Vegetated Margins of Arable Fields. Agriculture, Ecosystems and Environment, 72, 131-144. https://doi.org/10.1016/S0167-8809(98)00169-8 [Paper reference 1]
- 31. Thomas, C. F. G., Parkinson, L., Griffiths, G. J. K., Garcia, A. F, & Marshall, E. J. P. (2001). Aggregation and Temporal Stability of Carabid Beetle Distributions in Field and Hedgerow Habitats. Journal of Applied Ecology, 38, 100-116.https://doi.org/10.1046/j.1365-2664.2001.00574.x [Paper reference 1]
- 32. van Emden, H. F. (1976). Insect-Pest Management in Multiple Cropping Systems—A Strategy. International Rice Research Institute, Proceedings, Symposium on Cropping Systems Research and Development for the Asian Rice Farmer, Los Banos, 21-24 September 1976. [Paper reference 1]
- 33. Vanbergen, A. J., Woodcock, B A., Watt, A. D., & Niemela, J. (2005). Effects of Land-Use Heterogenity on Carabid Communities at the Landscape Scale. Ecograph, 28, 3-16.https://doi.org/10.1111/j.0906-7590.2005.03991.x [Paper reference 1]
Appendix 1
Mean individual count of orders collected by pitfall trapping during April and August 2006.
F value from one-way ANOVA; χ2 from Kruskal Wallis; *P < 0.1; **P < 0.05; ***P < 0.001; NS not significant; NT not tested; NP not present.